Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
I programmi di ricerca in banche dati possono essere
residenti in un calcolatore di cui siamo proprietari o utenti oppure essere utilizzabili via web

2
Il concetto di directory
bio1 directory principale acidi nucleici primo livello di sub-directories proteine secondo livello di sub-directories human coli mouse

3
Come si effettua una ricerca in una banca dati
si possono effettuare ricerche utilizzando parole-chiave (es.: emoglobina) e ricavando i nomi dei files che le contengono oppure si possono utilizzare sequenze in input per ricavare liste di sequenze simili ad esse
 e ricavando i nomi dei files che le contengono. oppure si possono utilizzare sequenze in input per ricavare liste di sequenze simili ad esse.")
4
confronto tra sequenze
a cosa serve effettuare ricerche per similarità di sequenza in banche dati confronto tra sequenze costruzione di alberi filogenetici identificazione di domini funzionali costruzione di modelli per omologia in 3D

5
similarità omologia ≠ ATTENZIONE alla proprietà di linguaggio!
é un dato che prescinde da eventuali ipotesi sulla causa della similarità stessa due sequenze si dicono omologhe se condividono una stessa origine filogenetica

6
La similarità biologica è spesso dovuta ad omologia, ma può anche presentarsi per caso oppure per fenomeni di convergenza adattativa Ad esempio: l’ala di un uccello e l’ala di un pipistrello si sono evolute indipendentemente e di conseguenza non sono omologhe Nel trattare le sequenze è sempre più corretto utilizzare il termine similarità, in quanto è sempre possibile stabilire quanto due sequenze siano simili, mentre non sempre si può decidere se la similarità sia dovuta ad omologia, a convergenza adattativa, oppure al caso

7
un po’ di terminologia…
strutture o sequenze ortologhe in due organismi sono sequenze omologhe che sono evolute dalla stessa caratteristica nel loro ultimo antenato comune ma che non necessariamente mantengono la loro funzione ancestrale. sequenze omologhe la cui evoluzione riflette invece eventi di duplicazione genica si definiscono paraloghe. per esempio, la catena dell’ emoglobina e’ un paralogo della catena dell’ emoglobina e della mioglobina, dal momento che ambedue si sono evolute dallo stesso gene ancestrale attraverso ripetuti eventi di duplicazione genica.

8
COPPIE in questa prima parte della lezione ci occuperemo solo di
di sequenze

9
allineamento similarità calcolo della similarità tra due sequenze date
non si possono allineare due sequenze senza definire criteri si similarità per valutare la similarità tra due sequenze, dobbiamo prima allinearle

10
INPUT acidi nucleici proteine
calcolo della similarità tra due sequenze date acidi nucleici (4 possibili residui nucleotidici) INPUT proteine (20 possibili residui aminoacidici) la bioinformatica in generale studia le sequenze come stringhe di caratteri
 INPUT. proteine. (20 possibili residui aminoacidici) la bioinformatica in generale studia le sequenze come stringhe di caratteri.")
11
calcolo della similarità tra due sequenze date
cominciamo quindi col definire una prima semplice misura di similarità, data dalla somma dei caratteri delle due sequenze che si appaiano esattamente facciamo scorrere una delle due sequenze sull’altra in tutte le posizioni possibili (generiamo tutti i possibili allineamenti) e valutiamo la similarità di sequenza di ognuno degli allineamenti generati
 e valutiamo la similarità di sequenza di ognuno degli allineamenti generati.")
12
AAKKQW AAKQW calcolo della similarità tra due sequenze date
definiamo similarità di sequenza tra le due sequenze come il più alto dei punteggi ottenuti l’allineamento associato alla più alta valutazione della similarità di sequenza verrà scelto come il migliore degli allineamenti possibili esempio: AAKKQW AAKQW

13
3 1 4 calcolo della similarità tra due sequenze date (senza gaps)
AAKKQW AAKKQW 4 3 1 AAKQW AAKQW AAKKQW AAKKQW AAKQW AAKQW AAKKQW AAKKQW AAKQW AAKQW AAKKQW AAKKQW AAKQW AAKQW AAKKQW AAKKQW AAKQW AAKQW

14
AAKKQW AAKQW 6 caratteri 5 caratteri
calcolo della similarità tra due sequenze date (senza gaps) AAKQW AAKKQW 6 caratteri 5 caratteri abbiamo valutato 10 (5+5) allineamenti abbiamo confrontato 30 (6x5) caratteri
 AAKQW. AAKKQW. 6 caratteri. 5 caratteri. abbiamo valutato 10 (5+5) allineamenti. abbiamo confrontato 30 (6x5) caratteri.")
15
1 2 3 4 5 6 7 8 9 10 Numero degli allineamenti generati 1 3 6 10 15 20 24 27 29 30 Numero dei confronti tra residui effettuati AAKKQW AAKQW

16
O(nm) ~ O(n2) calcolo della similarità tra due sequenze date
un programma per calcolatore ha un tempo di esecuzione che in generale dipende dal numero di operazioni necessarie per eseguirlo un programma che calcoli la similarità di sequenza tra due sequenze date deve effettuare un numero di confronti che dipende dal prodotto della lunghezza delle due sequenze O(nm) ~ O(n2)
 ~ O(n2)")
17
1) sequenza in input (query sequence)
ricerca per similarità in una banca dati Quando si deve effettuare una ricerca per similarità di sequenza in una banca dati, l’operazione di confronto tra due sequenze deve inoltre essere ripetuta per ogni coppia di sequenze: 1) sequenza in input (query sequence) 2) ognuna delle sequenze della banca dati
 sequenza in input (query sequence) 2) ognuna delle sequenze della banca dati.")
18
esiste quindi una forte necessità di utilizzare algoritmi RAPIDI!
ricerca per similarità in una banca dati esiste quindi una forte necessità di utilizzare algoritmi RAPIDI!

19
ricerca per similarità in una banca dati
… e non abbiamo ancora considerato tutte le ulteriori complicazioni legate alla possibile presenza di gaps nell’allineamento tra due sequenze!

20
10 25 calcolo della similarità tra due sequenze date
necessità di trattare l’inserimento e/o la delezione di caratteri (gaps) IPLMTRWDQEQESDFGHKLPIYTREWCTRG |||||||||| CHKIPLMTRWDQQESDFGHKLPVIYTREW 10 IPLMTRWDQEQESDFGHKLP-IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVIYTREW 25
 IPLMTRWDQEQESDFGHKLPIYTREWCTRG |||||||||| CHKIPLMTRWDQQESDFGHKLPVIYTREW. 10. IPLMTRWDQEQESDFGHKLP-IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVIYTREW. 25.")
21
calcolo della similarità tra due sequenze date
l’inserimento di gaps comporta una modifica del nostro iniziale SEMPLICE metodo di misura della similarità possiamo associare un punteggio di penalizzazione (gap penalty) per ogni gap aggiunto all’allineamento o attribuire un punteggio di penalizzazione diverso per l’apertura di un gap nell’allineamento o per il suo allungamento (gap extension penalty)
 per ogni gap aggiunto all’allineamento. o attribuire un punteggio di penalizzazione diverso per l’apertura di un gap nell’allineamento o per il suo allungamento (gap extension penalty)")
22
calcolo della similarità tra due sequenze date
gap creation penalty (es.: -1 per ogni gap) IPLMTRWDQEQESDFGHKLP-IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVIYTREW gap extension penalty (es.: -0.1 per ogni ins/del successiva alla prima) IPLMTRWDQEQESDFGHKLP----IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVGSSIYTREW
 IPLMTRWDQEQESDFGHKLP-IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVIYTREW. gap extension penalty (es.: -0.1 per ogni ins/del successiva alla prima) IPLMTRWDQEQESDFGHKLP----IYTREWCTRG ||||||||| |||||||||| |||||| CHKIPLMTRWDQ-QESDFGHKLPVGSSIYTREW.")
23
LENTO calcolo della similarità tra due sequenze date
un algoritmo di allineamento che tenesse conto del possibile inserimento di un gap in ogni possibile posizione delle due sequenze e di ogni possibile lunghezza di un gap in ogni possibile posizione sarebbe estremamente LENTO

24
nuovi algoritmi calcolo della similarità tra due sequenze date
da ciò discende la necessità di trovare soluzioni alternative, ovvero nuovi algoritmi per l’allineamento delle sequenze

25
Matrici di punti - dot matrices
consideriamo una matrice in cui la prima riga in alto coincide con la sequenza1 scritta da sinistra a destra e in cui la prima colonna a sinistra coincide con la sequenza2 scritta dall’alto in basso in ogni punto in cui la sequenza nella riga e nella colonna coincidono, introduciamo un asterisco nel grafico. Le regioni delle sequenze che possono essere allineate senza introdurre gap emergono come una serie contigua di punti sulla diagonale

26
calcolo della similarità tra due sequenze date - Uso di matrici di punti (DOT MATRICES)
g e t d y h o f l k m a r g e t d y h o f *

27
calcolo della similarità tra due sequenze date - Uso di matrici di punti (DOT MATRICES)
le dot matrices esprimono una buona rappresentazione grafica di un allineamento consentono di visualizzare similarità di sequenza anche in presenza di gaps, che appaiono come ‘salti di diagonale’ esistono programmi in grado di sfruttare gli schemi tipo ‘dot matrix’ per valutare la similarità tra sequenze e identificare il miglior allineamento

28
Matrici di punti - dot matrices
il rumore di fondo é molto alto perché molti dei match tra sequenze costruiti in questo modo sono casuali e dipendono da singole occorrenze dello stesso residuo in posizioni diverse delle due sequenze. possiamo calcolare il numero di match in una finestra, per esempio di 5 o 15 residui / basi, e decidere di introdurre un punto nel grafico solo se una certa percentuale minima di questi (es. 50%) sono identici. il punto viene posizionato al centro della finestra.
 sono identici. il punto viene posizionato al centro della finestra.")
29
calcolo della similarità tra due sequenze date - Uso di matrici di punti (DOT MATRICES)
")
30
Le ricerche di similarità si basano sul confronto sistematico di una sequenza di partenza (generalmente chiamata query) con ognuna delle sequenze del database. Considerando che molti database di sequenze, come ad esempio GenBank contengono oltre un milione di sequenze diverse è necessario disporre di programmi in grado di portare avanti queste ricerche in modo automatico. Alcuni programmi, come BLAST, sono estremamente efficienti e sono in grado di portare a termine una ricerca di similarità in pochi secondi. Questi programmi si basano essenzialmente su procedure che: * per ogni sequenza del database * effettua l'allineamento; * attribuisce un punteggio; * se il punteggio è alto lo memorizza; * ripete fino alla fine delle sequenze; * mostra la lista dei migliori allineamenti trovati

31
E' importante notare che è essenziale attribuire un punteggio agli allineamenti, altrimenti non si avrebbe modo di stabilire se un allineamento è migliore di un altro. Deve essere notato che con l'approccio della dot matrix non viene trovato alcun allineamento, ma viene semplicemente mostrata una matrice che deve essere interpretata visivamente. Quindi per effettuare ricerche di similarità questo semplice approccio non e’ sufficiente: abbiamo bisogno di un sistema che sia anche in grado di trovare automaticamente gli allineamenti migliori. Molto schematicamente il problema degli allineamenti viene risolto da programmi che sono in grado di identificare il "percorso" migliore all'interno di una dot matrix. Cioè il percorso che totalizza il massimo punteggio. Per percorso si intende l'insieme di caselle che corrispondono agli aminoacidi appaiati.

32
MATRICI DI SOSTITUZIONE
calcolo della similarità tra due sequenze date - le matrici di sostituzione nel caso delle sequenze proteiche, gli allineamenti e il calcolo della similarità possono essere notevolmente migliorati dall’introduzione di diversi schemi di punteggio, noti come MATRICI DI SOSTITUZIONE che comprendono punteggi diversi da 0 e da 1 per l’appaiamento di residui amminoacidici

33
Molto semplicemente, il punteggio può essere calcolato attribuendo un valore positivo ad ogni lettera appaiata correttamente e una penalità ad ogni gap. La somma di questi valori rappresenta quindi il punteggio dell'intero allineamento. Nel caso più semplice possiamo assegnare il valore di '1' ad ogni identità e '0' ad ogni "mismatch" cioè ai caratteri non appaiati correttamente. Un tale criterio non è però molto valido perchè non ha senso penalizzare tutti i mismatch allo stesso modo. Ad esempio la leucina e l'isoleucina sono molto simili; quindi se questi due aminoacidi vengono appaiati possiamo pensare che viene rispettato un certo criterio di similarità, anche se non c'è identità assoluta. Estendendo questo ragionamento possiamo attribuire ad ogni possibile coppia di aminoacidi un punteggo di appaiamento. In questo modo otteniamo una matrice di sostituzione.

34
le matrici di sostituzione
potremmo raggruppare residui in classi a seconda della similarità delle loro caratteristiche chimico-fisiche, e sommare 1 al punteggio per appaiamenti di residui della stessa classe e sottrarre 1 per residui di classi diverse E D R K H C I L V G A F Y W P

35
le matrici di sostituzione
Margaret Dayhoff raccolse statistiche sulle frequenze di sostituzioni amminoacidiche nelle sequenze proteiche allora note via via che le sequenze divergono, le mutazioni si accumulano per misurare la probabilità relativa di una particolare sostituzione (per esempio Asp--> Glu) possiamo contare quanti Asp sono diventati Glu in allineamenti di sequenze omologhe
 possiamo contare quanti Asp sono diventati Glu in allineamenti di sequenze omologhe.")
36
1 PAM = 1 Percent Accepted Mutation
le matrici di sostituzione è comunque necessario evitare di considerare allineamenti in cui possano essere avvenuti sostituzioni multiple in determinate posizioni per cui questi calcoli devono venire effettuati su coppie di sequenze MOLTO SIMILI tra loro, in modo da poter assumere che nessuna posizione è mutata più di una volta la divergenza di due sequenze si può misurare in PAM: 1 PAM = 1 Percent Accepted Mutation

37
le matrici di sostituzione
due sequenze sono separate da 1 PAM se hanno il 99% di identità la frequenza delle sostituzioni amminoacidiche può essere calcolata in coppie di sequenze poco divergenti (1 PAM) frequenze di sostituzioni amminoacidiche per sequenze più divergenti possono essere calcolate a partire da queste, fino ad ottenere matrici PAM 250, corrispondenti a similarità di sequenza del 20%
 frequenze di sostituzioni amminoacidiche per sequenze più divergenti possono essere calcolate a partire da queste, fino ad ottenere matrici PAM 250, corrispondenti a similarità di sequenza del 20%")
38
le matrici di sostituzione
PAM % identità se due sequenze sono filogeneticamente distanti è opportuno usare matrici PAM con indici più alti, e viceversa

39
Il tutto si può facilmente rappresentare con una matrice di 20x20, quattrocento valori. La seguente tabella corrisponde alla matrice PAM250

40
Allineamento globale o locale?
LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||. | | | .| .| || || | || TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHKAG Allineamento locale: TGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||||||||.||||| TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHK

41
Ma qual è l’allineamento MIGLIORE?
Allineamento globale o locale? sost. cons. 3 1 id. 14 13 Allineamento globale: LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||. | | | .| .| || || | || TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHKAG Allineamento locale: TGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||||||||.||||| TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHK Ma qual è l’allineamento MIGLIORE?

42
Allineamento globale o locale?
1) scegliamo il miglior allineamento dal punto di vista biologico, e poi… 2) cerchiamo il modo di privilegiarlo dal punto di vista computazionale spesso gli allineamenti locali hanno una migliore rispondenza con la realtà funzionale
 scegliamo il miglior allineamento dal punto di vista biologico, e poi… 2) cerchiamo il modo di privilegiarlo dal punto di vista computazionale. spesso gli allineamenti locali hanno una migliore rispondenza con la realtà funzionale.")
43
Allineamento globale o locale?
similarità locali servono a identificare proteine anche diverse, ma che contengono lo stesso dominio

44
Allineamento globale o locale?
a livello di DNA, troviamo regioni con similarità locali che riflettono situazioni interessanti: ad esempio introni/esoni, inserzioni/delezioni, transposoni, regioni promotore… esoni introni

45
Allineamento globale o locale?
gli allineamenti globali possono comunque essere utilizzati per confrontare accuratamente due sequenze la cui similarità sia estesa per tutta la lunghezza.

46
allineamenti tra due sequenze
abbiamo definito un nuovo schema di punteggi per la valutazione della similarità tra due sequenze una matrice di sostituzione per valutare l’appaiamento tra qualsiasi coppia di residui penalizzazioni appropriate per l’apertura o l’estensione di un gap

47
programmazione dinamica
allineamenti tra due sequenze ci serve ora un metodo (un algoritmo) per generare il miglior allineamento possibile tra due sequenze, tenendo anche conto delle possibili inserzioni e delezioni un algoritmo ‘famoso’ per l’allineamento tra due sequenze si basa su una tecnica matematica nota come programmazione dinamica
 per generare il miglior allineamento possibile tra due sequenze, tenendo anche conto delle possibili inserzioni e delezioni. un algoritmo ‘famoso’ per l’allineamento tra due sequenze si basa su una tecnica matematica nota come. programmazione dinamica.")
48
allineamenti tra due sequenze - programmazione dinamica
buone notizie: il metodo trova il miglior allineamento globale tra due sequenze cattive notizie: 1) spesso l’output mostra più allineamenti DIVERSI col massimo del punteggio 2) ci mette TROPPO TEMPO per effettuare una ricerca esaustiva
 spesso l’output mostra più allineamenti DIVERSI col massimo del punteggio. 2) ci mette TROPPO TEMPO per effettuare una ricerca esaustiva.")
49
Algoritmi dinamici di allineamento
per effettuare un allineamento è prima di tutto necessario scegliere una matrice di sostituzione per valutare gli appaiamenti tra residui e definire dei punteggi di penalizzazione per i gaps algoritmi di allineamento che utilizzano una tecnica di programmazione dinamica: Needleman e Wunsch (1970) Smith e Waterman (1981)
 Smith e Waterman (1981)")
50
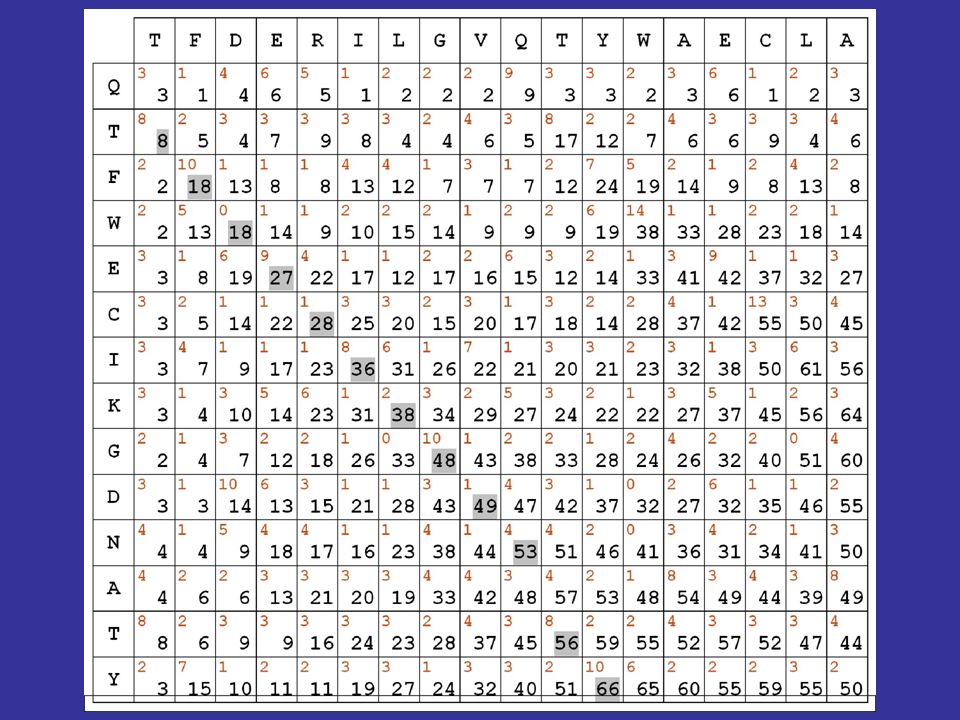
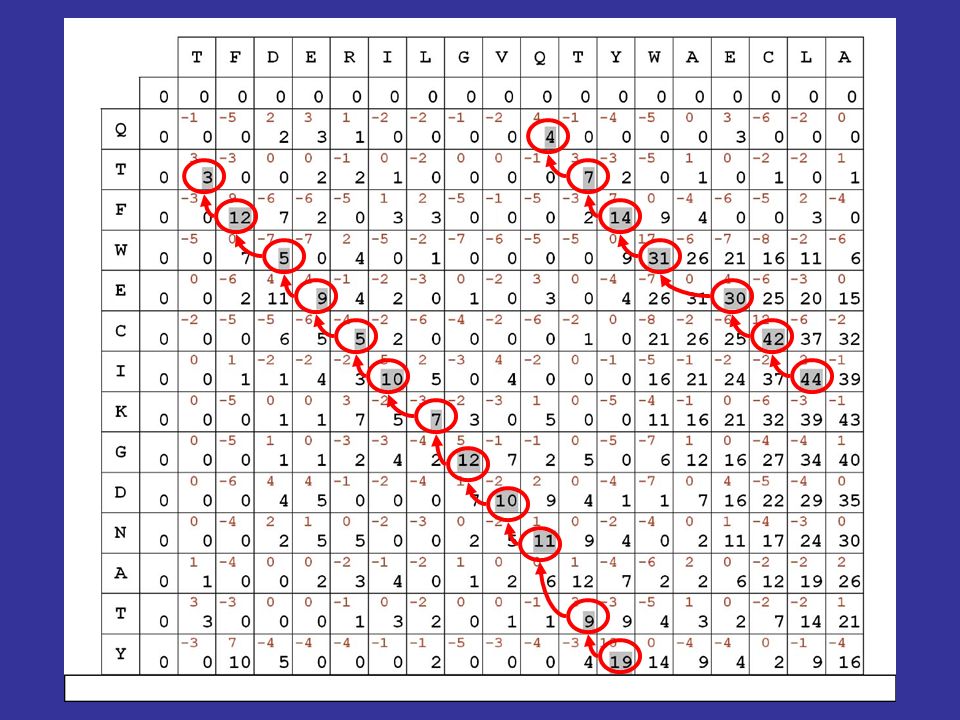
Algoritmi dinamici di allineamento
dividiamo la procedura in 3 passi successivi 1) consideriamo le due sequenze da allineare in una specie di dot matrix : nelle caselle scriviamo i punteggi in rosso derivati dalla matrice di sostituzione scelta
 consideriamo le due sequenze da allineare in una specie di dot matrix : nelle caselle scriviamo i punteggi in rosso derivati dalla matrice di sostituzione scelta.")
52
Algoritmi dinamici di allineamento
calcolando le somme lungo le diagonali, effettueremmo un’operazione equivalente al calcolo dei punteggi ottenuto facendo scorrere le due sequenze l’una sull’altra resterebbe aperto il problema della valutazione dei gaps

53
Algoritmi dinamici di allineamento
passo 2 della procedura: ricerca del percorso che consente di ottenere il massimo punteggio in base a delle regole stabilite, tenendo anche conto dei gaps qualche semplice osservazione ci consente di risparmiare tempo di calcolo:

54
Algoritmi dinamici di allineamento
se una sequenza è scritta da sinistra a destra e l’altra dall'alto in basso, allora qualsiasi percorso valido deve mantenere sempre una direzione tendenziale che va dall'angolo in alto a sinistra a quello in basso a destra

55
Algoritmi dinamici di allineamento
Misura della similarità: i punteggi in diagonale si SOMMANO fuori dalla diagonale, si PENALIZZA di 5 punti

56
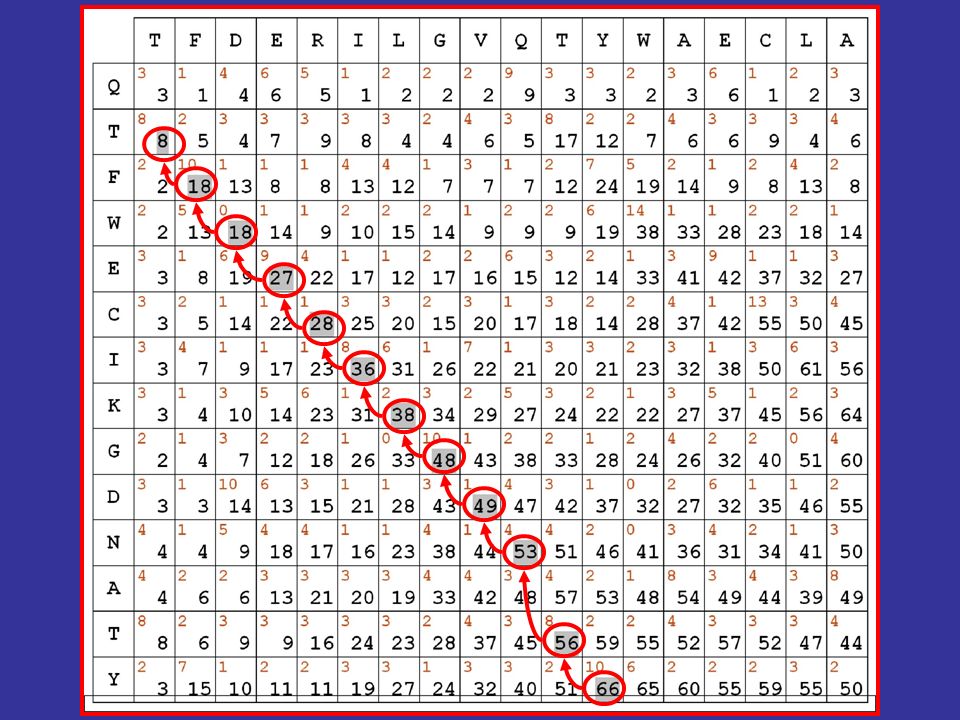
Algoritmi dinamici di allineamento
vediamo quindi come si calcolano le cifre riportate in nero nella matrice evidenzieremo poi in grigio il migliore percorso all’interno della matrice, secondo le regole e i punteggi stabiliti

58
Algoritmi dinamici di allineamento
il migliore allineamento globale per le sequenze in matrice risulta quindi il seguente: TFDERILGVQ-TYWAECLA || | | | . || QTFWECIKGDNATY

59
Algoritmi dinamici di allineamento
il fatto di aver usato matrici di sostituzione contenenti esclusivamente valori positivi fa sì che il valore massimo della matrice si trovi sempre nell’ultima riga o nell’ultima colonna ne consegue che l’allineamento ottenuto è un allineamento globale

60
Algoritmi dinamici di allineamento
la procedura descritta corrisponde quasi esattamente all’algoritmo per l’allineamento globale pubblicato da Needleman e Wunsch nel 1970

61
Algoritmi dinamici di allineamento
se le matrici contenessero invece sia valori positivi che negativi (come le pam viste nel corso della scorsa lezione), i valori più alti potrebbero trovarsi anche in porzioni INTERNE alla matrice e descrivere di conseguenza allineamenti locali
, i valori più alti potrebbero trovarsi anche in porzioni INTERNE alla matrice e descrivere di conseguenza allineamenti locali.")
63
Algoritmi dinamici di allineamento
TFDERILGVQ-TYWAECLA || | | | . || QTFWECIKGDNATY Allineamento globale TFDERILGVQTYWAECLA ||.| ||. QTFW-ECIKGDNATY Allineamento locale

64
é importante notare che il punteggio massimo lo è in senso relativo
Algoritmi dinamici di allineamento il valore assoluto dei punteggi che associamo agli allineamenti dipende dai valori contenuti nella matrice di sostituzione che utilizziamo ci sono matrici i cui valori variano tra 0 e 100, altre che variano da -1 a +1, adottando punteggi decimali é importante notare che il punteggio massimo lo è in senso relativo

65
Algoritmi dinamici di allineamento
L’ algoritmo di Needleman e Wunsch è stato sviluppato per l’allineamento globale L’ algoritmo di Smith e Waterman è stato sviluppato per l’allineamento locale Ma ciò che realmente fa diventare un algoritmo di questo tipo locale o globale è il tipo di matrice di sostituzione che si usa: se contiene solo valori allineamenti globali se contiene valori +/ allineamenti locali

66
Metodi euristici per l’allineamento
gli algoritmi descritti effettuano delle ricerche esaustive ed esplorano tutto lo spazio degli allineamenti possibili si tratta comunque di algoritmi di ordine n2, ovvero per allineare due sequenze lunghe ognuna 1000 residui, effettuano 1000x1000 = un milione di confronti

67
similarità di sequenza
Metodi euristici per l’allineamento PGM allineamento similarità di sequenza seq1 seq2 per effettuare ricerche di similarità in banche dati, c’è comunque necessità di algoritmi più veloci PGM lista di proteine simili alla query seq DB

68
Metodi euristici per l’allineamento
la crescita esponenziale delle dimensioni delle banche dati di sequenze biologiche ha portato allo sviluppo di programmi (come FASTA e BLAST) in grado di effettuare velocemente ricerche di similarità, grazie a soluzioni euristiche che, come vedremo, sono basate su assunzioni non certe, ma estremamente probabili. in pratica la ricerca è resa più veloce a scapito della certezza di avere veramente trovato la soluzione migliore.
 in grado di effettuare velocemente ricerche di similarità, grazie a soluzioni euristiche che, come vedremo, sono basate su assunzioni non certe, ma estremamente probabili. in pratica la ricerca è resa più veloce a scapito della certezza di avere veramente trovato la soluzione migliore.")
69
Metodi euristici per l’allineamento (FASTA)
ricerca di parole contigue per sola identità sulla stessa diagonale calcolo di punteggi con matrice di sostituzione, selezione dei migliori punteggi

70
Metodi euristici per l’allineamento (FASTA)
unione di frammenti che possono essere congiunti entro una soglia di accettabilità applicazione di S&W su una stretta banda per ottimizzare l’allineamento

71
Lipman e Pearson Metodi euristici per l’allineamento (FASTA)
FASTA è stato pubblicato nel 1985 da Lipman e Pearson

72
similarità di sequenza
Allineamento multiplo di sequenze PGM allineamento similarità di sequenza seq1 seq2 PGM lista di proteine simili alla query seq DB é possibile utilizzare una lista di sequenze simili tra loro per generare un ALLINEAMENTO MULTIPLO DI SEQUENZE

Presentazioni simili












>")





 Algoritmi di allineamento 2) Algoritmi di ricerca in database>")
