Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
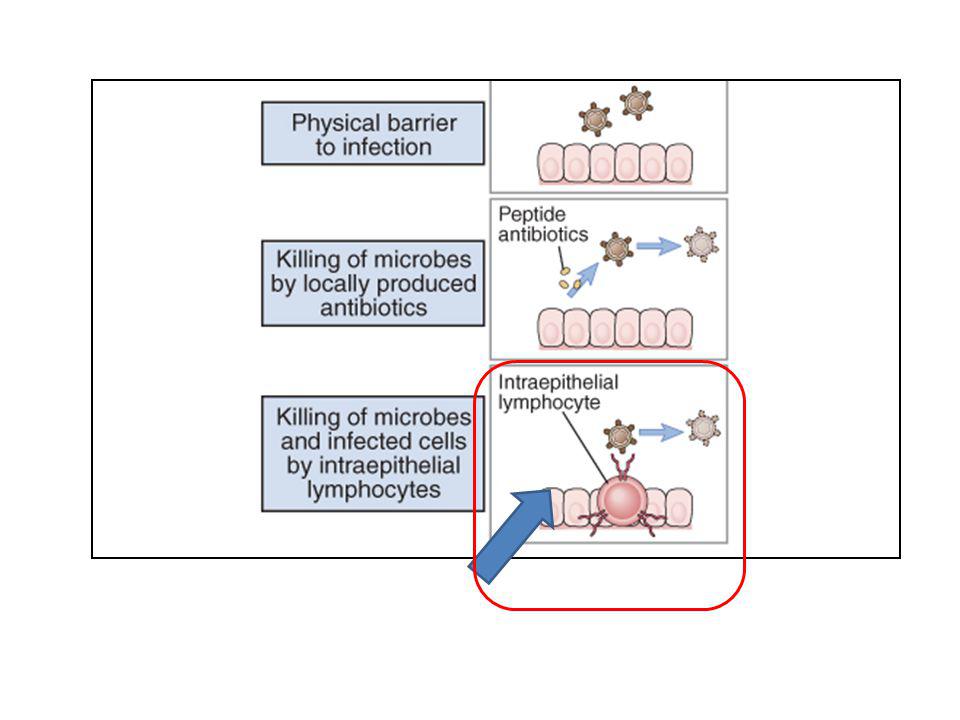
Barriere anatomiche: Fattori meccanici:
epidermide della pelle, mucose del tratto respiratorio e gastrointestinale Le superfici epiteliali costituiscono una vera barriera fisica all’invasione dei microorganismi. Le cellule epiteliali infatti sono tenute insieme da giunzioni strette che formano un vero muro contro l’ambiente esterno. Fattori meccanici: -La desquamazione dell’epidermide consente la rimozione dei microrganismi che hanno aderito alla superficie epiteliale. - Il movimento delle ciglia dell’epitelio respiratorio e la peristalsi intestinale facilitano l’eliminazione degli agenti patogeni entrati con l’aria inspirata e con l’alimentazione. -Il muco, prodotto nel tratto respiratorio ed intestinale, intrappola i microorganismi consentendone l’eliminazione. -Le lacrime e la saliva generano un movimento fluido che previene l’instaurarsi di infezioni a livello della bocca e degli occhi. Nabissi 14

2
DIFESE PRIMARIE: BARRIERE CHIMICO-FISICHE
Nabissi 14

3
The different steps of the response against pathogens.
Nabissi 14

5
Fattori chimici Peptidi antibiotici detti defensine.
La sintesi delle defensine avviene nel midollo osseo, nei precursori dei neutrofili (pro-mielociti). Le defensine sono inpacchettate in granuli primari (azzurrofili) che si fondono poi con i vacuoli dei fagociti. I vacuoli vengono poi rilasciati nel sangue e nei tessuti. Nabissi 14 Nabissi 2012
. Le defensine sono inpacchettate in granuli primari (azzurrofili) che si fondono poi con i vacuoli dei fagociti. I vacuoli vengono poi rilasciati nel sangue e nei tessuti. Nabissi 14. Nabissi")
6
-il lisozima e la fosfolipasi A contenuti nelle lacrime e nella saliva degradano
la parete dei batteri. Il pH acido dello stomaco e i succhi digestivi, ricchi di enzimi come la pepsina e di acidi grassi, uccidono i microrganismi che invadono il tratto gastro-intestinale. Il fluido presente sugli epiteli di rivestimento dei polmoni è ricco di due proteine surfactante A e B che rivestono i microrganismi facilitandone l’eliminazione. le cellule del Paneth dell’intestino producono peptidi antibatterici dette criptocidine. Nabissi 14

7
RECETTORI SOLUBILI DELL’IMMUNITA’ INNATA I recettori solubili comprendono i fattori della cascata del complemento C3, la lectina legante il mannosio (MBL), le proteine A e D del surfactante (SP-A, SP-D) e le pentrassine. MBL, SP-A e SP-D appartengono alla famiglia delle collettine, data la presenza di un dominio di tipo collagenico e sono presenti normalmente nel sangue, aumentando di concentrazione durante l’infiammazione (es. la proteina C reattiva prodotta nel fegato). Questi recettori solubili sono in grado di riconoscere strutture della membrana batterica e funginea, in particolare i domini zuccherinici e lipidici, legandosi con scarsa specificità ma con alta avidita’ in quanto piu’ MBL possono legarsi allo stesso batterio. Le pentrassine riconoscono cellule apoptotiche e materiale nucleare di cellule morte, quindi agendo principalmente su cellule del proprio organismo (self). Nabissi 14

8
Fattori biologici La maggior parte delle superfici epiteliali è associata ad una normale flora di batteri non patogeni che competono con i microorganismi patogeni per il nutrimento ed i siti di attacco alle cellule epiteliali. La normale flora può anche produrre sostanze antimicrobiche come l’acido lattico o batteriotossine. Nabissi 14

9
Quando un microrganismo attraversa le barriere anatomiche e comincia a replicare nei tessuti sottostanti, viene inizialmente fagocitato da macrofagi residenti nel tessuto connettivo. Grazie alle sostanze chemiotattiche liberate nel sito di infezione, viene reclutato un gran numero di neutrofili. Nelle prime fasi dell’infiammazione acuta la componente cellulare principale è rappresentata dai neutrofili. I neutrofili hanno vita breve e muoiono dopo aver compiuto un ciclo di fagocitosi. Il pus è costituito da neutrofili morti o che stanno morendo. Nabissi 14

10
Globuli Bianchi o Leucociti:
-granulociti polimorfonucleati (neutrofili, eosinofili e basofili) -linfociti -monociti (si differenziano in macrofagi nei tessuti) Globuli rossi: -eritrociti Trombociti: -piastrine Nabissi 14
 -linfociti. -monociti (si differenziano in macrofagi nei tessuti) Globuli rossi: -eritrociti. Trombociti: -piastrine. Nabissi 14.")
11
Nabissi 14

12
INFIAMMAZIONE Nabissi 14

13
CARATTERISTICHE GENERALI DELL’INFIAMMAZIONE
L’infiammazione è una reazione complessa ad agenti lesivi, come microbi e cellule danneggiate, in genere necrotiche, e consiste nella risposta vascolare, nella migrazione ed attivazione dei leucociti ed in una reazione sistemica. Caratteristica peculiare della reazione infiammatoria è la reazione dei vasi sanguigni, che porta all’accumulo di liquidi e leucociti nei tessuti extravascolari. Nabissi 14

14
L’infiammazione si differenzia in:
Acuta : esordio rapido e durata relativamente breve, con formazione di essudato (edema) e migrazione di leucociti. Cronica: ha una durata maggiore ed è associata istologicamente alla presenza di linfociti e macrofagi, proliferazione di vasi sanguigni, fibrosi e necrosi tissutale. LE RISPOSTE VASCOLARI E CELLULARI DELL’INFIAMMAZIONE, SIA ACUTA CHE CRONICA, SONO MEDIATE DA FATTORI CHIMICI CHE SONO PRODOTTI IN RISPOSTA ALLO STIMOLO INFIAMMATORIO. I mediatori chimici amplificano la risposta infiammatoria, agendo a tempi diversi. Al termine del processo infiammatorio i mediatori vengono degradati o dispersi. Nabissi 14
 e migrazione di leucociti. Cronica: ha una durata maggiore ed è associata istologicamente alla presenza di linfociti e macrofagi, proliferazione di vasi sanguigni, fibrosi e necrosi tissutale. LE RISPOSTE VASCOLARI E CELLULARI DELL’INFIAMMAZIONE, SIA ACUTA CHE CRONICA, SONO MEDIATE DA FATTORI CHIMICI CHE SONO PRODOTTI IN RISPOSTA ALLO STIMOLO INFIAMMATORIO. I mediatori chimici amplificano la risposta infiammatoria, agendo a tempi diversi. Al termine del processo infiammatorio i mediatori vengono degradati o dispersi. Nabissi 14.")
15
- La risposta infiammatoria è strettamente legata al processo di riparazione, in quanto l’infiammazione distrugge, diluisce o isola l’agente infettivo e mette in moto dei meccanismi che mirano alla guarigione ed alla ricostruzione del tessuto danneggiato (rigenerazione (cell.parench.) o cicatrizzazione (tess. fibroso) ) - L’infiammazione è una risposta protettiva che consiste di due componenti principali: reazione vascolare e reazione cellulare. Nabissi 14

16
-La reazione cellulare comprende cellule circolanti (neutrofili, monociti, eosinofili, linfociti, basofili e piastrine) e cellule del tessuto connettivo (mastociti [circondano i vasi sanguigni], fibroblasti [tessuto connettivo], macrofagi e linfociti. -La matrice extracellulare e la membrana basale partecipano alla risposta infiammatoria attraverso proteine fibrose strutturali (collagene, elastina), glicoproteine adesive (fibronectina, laminina, collagene) e proteoglicani. Nabissi 14
![-La reazione cellulare comprende cellule circolanti (neutrofili, monociti, eosinofili, linfociti, basofili e piastrine) e cellule del tessuto connettivo (mastociti [circondano i vasi sanguigni], fibroblasti [tessuto connettivo], macrofagi e linfociti.](http://slideplayer.it/slide/2787254/10/images/16/-La+reazione+cellulare+comprende+cellule+circolanti+%28neutrofili%2C+monociti%2C+eosinofili%2C+linfociti%2C+basofili+e+piastrine%29+e+cellule+del+tessuto+connettivo+%28mastociti+%5Bcircondano+i+vasi+sanguigni%5D%2C+fibroblasti+%5Btessuto+connettivo%5D%2C+macrofagi+e+linfociti..jpg "-La matrice extracellulare e la membrana basale partecipano alla risposta infiammatoria attraverso proteine fibrose strutturali (collagene, elastina), glicoproteine adesive (fibronectina, laminina, collagene) e proteoglicani. Nabissi 14.")
17
CARATTERISTICHE GENERALI DELL’INFIAMMAZIONE
COMPONENTI DELLA RISPOSTA INFIAMMATORIA ACUTA E CRONICA Nabissi 14

18
Le reazioni infiammatorie ACUTA sono scatenate da diversi stimoli.
Infezioni (batteri, virus, parassiti) Traumi (superficiali, profondi) Agenti fisici e chimici (ustioni, congelamento), U.V., sostanze chimiche ambientali) Necrosi tissutale (varie cause) Reazioni immunitarie (ipersensibilità) Per quanto gli stimoli inducono reazioni in parte distinte, alcune caratteristiche base rimangono comuni. Nabissi 14
 Traumi (superficiali, profondi) Agenti fisici e chimici (ustioni, congelamento), U.V., sostanze chimiche ambientali) Necrosi tissutale (varie cause) Reazioni immunitarie (ipersensibilità) Per quanto gli stimoli inducono reazioni in parte distinte, alcune. caratteristiche base rimangono comuni. Nabissi 14.")
19
CARATTERISTICHE GENERALI DELL’INFIAMMAZIONE
EFFETTI LOCALI EFFETTI SISTEMICI Rubor (rossore) Sintomi specifici della patologia Calor (calore) - Febbre Tumor (tumefazione) - Anoressia Dolor (dolore) - Debolezza Funcio lesa (lesione) Alterazione eq.idrico - Perdita minerali Leucocitosi Modificazioni biochimico-cliniche di fase acuta. Nabissi 14
 Sintomi specifici della patologia. Calor (calore) - Febbre. Tumor (tumefazione) - Anoressia. Dolor (dolore) - Debolezza. Funcio lesa (lesione) - Alterazione eq.idrico. - Perdita minerali. Leucocitosi. Modificazioni biochimico-cliniche. di fase acuta. Nabissi 14.")
20
INFIAMMAZIONE ACUTA L’infiammazione acuta è la risposta rapida che serve a portare nella sede della lesione mediatori, leucociti e proteine plasmatiche. Consiste in: - Alterazione del calibro vascolare che determinano un aumento del flusso ematico. - Modificazioni strutturali nella microvascolarizzazione: aumento della permeabilità che comporta la fuoriuscita di liquidi e proteine plasmatiche dal circolo sanguigno (formazione di edema) - Fuoriuscita dei leucociti dal microcircolo, accumolo nella sede di lesione e attivazione per l’eliminazione della causa infettiva video Nabissi 14
 - Fuoriuscita dei leucociti dal microcircolo, accumolo nella sede di lesione e attivazione per l’eliminazione della causa infettiva. video. Nabissi 14.")
21
MODIFICAZIONI VASCOLARI
VASODILATAZIONE: porta ad un aumento del flusso ematico, causando il rossore ed il calore della regione colpita dal processo infiammatorio. Indotta da istamina e ossido nitrico che agiscono sulla muscolatura liscia dei vasi. AUMENTO DELLA PERMEABILITA’: la fuoriuscita di liquido porta ad un aumento della viscosità del sangue (STASI) con conseguente concentrazione dei globuli rossi al centro dei piccoli vasi ed accumulo dei leucociti in prossimità dell’endotelio. Nabissi 14
 con conseguente concentrazione dei globuli rossi al centro dei piccoli vasi ed accumulo dei leucociti in prossimità dell’endotelio. Nabissi 14.")
22
UNO DEI FENOMENI PRECOCI CHE SI MANIFESTANO NEL SITO DELL’INFIAMMAZIONE E’ L’ALTERAZIONE DEL FLUSSO EMATICO E DEL CALIBRO VASCOLARE. La vasodilatazione permette un rallentamento del flusso ematico e l’avvicinamento dei leucociti alla parete dei vasi (2). L’adesione dei leucociti all’endotelio è mediata dal riconoscimento fra molecole adesive leucocitarie ed endoteliali (3). I fattori chemio-tattici regolano la motilità e la migrazione direzionale dei leucociti nel sito infiammatorio tissutale (4). Nabissi 14
. L’adesione dei leucociti all’endotelio è mediata dal riconoscimento fra molecole adesive leucocitarie ed endoteliali (3). I fattori chemio-tattici regolano la motilità e la migrazione direzionale dei leucociti nel sito infiammatorio tissutale (4). Nabissi 14.")
23
FORMAZIONE DELL’EDEMA
Con la perdita delle proteine plasmatiche si riduce la pressione osmotica all’interno dei vasi ed aumenta quella dei liquidi interstiziali. Questo fenomeno in associazione all’aumento della pressione idrostatica (data dalla vasodilatazione), causa una notevole fuoriuscita di liquidi nei tessuti interstiziali ed una riduzione del rientro di liquidi a livello delle venule post-capillari. Nabissi 14
, causa una notevole fuoriuscita di liquidi nei tessuti interstiziali ed una riduzione del rientro di liquidi a livello delle venule post-capillari. Nabissi 14.")
24
EDEMA Edema: puo’ essere essudato o trasudato ed indica un eccesso di liquido nelle cavità sierose o interstiziali. Essudazione: fuoriuscita di liquido, proteine e cellule ematiche dal sistema vascolare verso il tessuto interstiziale o nelle cavità corporee, causato da un’alterazione della permeabilità dei vasi Trasudazione: fuoriuscita di liquido dal sistema vascolare verso il tessuto interstiziale o nelle cavità corporee, derivato da uno squilibrio osmotico o idrostatico. Non si ha aumento della permeabilità vascolare. Pus: essudato purolento ricco di leucociti, cellule morte e spesso microbi. Nabissi 14

25
In che modo l’endotelio diventa permeabile?
Nabissi 14

26
Formazione di aperture nell’endotelio delle venule
Indotto da istamina, bradichinine, leucotrieni, Sostanza P ed altri mediatori chimici, con risposta rapida, reversibile e di breve durata: RISPOSTA IMMEDIATA TRANSITORIA (15-30 min). Interessa le venule di piccolo diametro e non capillari ed arteriole (probabilimente per la presenza di una minore concentrazione di recettori). Il legame dei mediatori chimici ai loro recettori presenti a livello delle cellule endoteliali attiva dei segnali intracellulari (fosforilazione di proteine del citoscheletro) che porta alla contrazione delle cellule endoteliali e alla separazione delle giunzioni intercellulari. Altri fattori coinvolti sono IL-1, TNF ed IFN-g. L’azione di queste citochine è ritardata e di lunga durata. Nabissi 14
. Interessa le venule di piccolo diametro e non capillari ed arteriole (probabilimente per la presenza di una minore concentrazione di recettori). Il legame dei mediatori chimici ai loro recettori presenti a livello delle cellule endoteliali attiva dei segnali intracellulari (fosforilazione di proteine del citoscheletro) che porta alla contrazione delle cellule endoteliali e alla separazione delle giunzioni intercellulari. Altri fattori coinvolti sono IL-1, TNF ed IFN-g. L’azione di queste citochine è ritardata e di lunga durata. Nabissi 14.")
27
Lesioni endoteliale diretta con necrosi e distacco delle cellule endoteliali
Danno diretto all’endotelio dello stimolo lesivo (ustioni, infezioni batteriche litiche), è una risposta immediata sostenuta. La fuoriuscita di liquidi e proteine inizia immediatamente e perdura a lungo fino alla riparazione o trombizzazione dei vasi. In questa risposta sono coinvolti tutti i tipi di vasi. Nabissi 14
, è una risposta immediata sostenuta. La fuoriuscita di liquidi e proteine inizia immediatamente e perdura a lungo fino alla riparazione o trombizzazione dei vasi. In questa risposta sono coinvolti tutti i tipi di vasi. Nabissi 14.")
28
Lesioni endoteliali mediate da leucociti
I leucociti aderiscono alle pareti dell’endotelio dopo un breve periodo dall’inizio del processo infiammatorio e possono liberare metaboliti tossici (radicali liberi dell’O2 od enzimi proteolitici) che causano danno alle pareti dei vasi, dando luogo a maggiore permeabilità vascolare. Soprattutto si verifica a livello dei capillari polmonari e glomerulari. Nabissi 14
 che causano danno alle pareti dei vasi, dando luogo a maggiore permeabilità vascolare. Soprattutto si verifica a livello dei capillari polmonari e glomerulari. Nabissi 14.")
29
Aumento della transcitosi
Formazione di canali costituiti da agglomerati di vescicole e vacuoli interconnessi, detti organuli vescicolo-vacuolari, localizzati principalmente a livello delle giunzioni cellulari. Il VEGF e l’istamina aumentano il numero e le dimensioni di questi organuli portando ad un aumento della permeabilità. Nabissi 14

30
Fuoriuscita di liquidi e proteine da vasi neoformati
Durante la riparazione le cellule endoteliali proliferano e formano nuovi vasi (angiogenesi). La fuoriuscita, da vasi neoformati con il processo dell’angiogenesi, perdura fino a quando le cellule endoteliali dei vasi non si differenziano completamente (diventando poco permeabili). Inoltre alcuni fattori come il VEGF che stimolano il processo angiogenetico rendono i vasi pre-esistenti piu’ permeabili. Nabissi 14
. La fuoriuscita, da vasi neoformati con il processo dell’angiogenesi, perdura fino a quando le cellule endoteliali dei vasi non si differenziano completamente (diventando poco permeabili). Inoltre alcuni fattori come il VEGF che stimolano il processo angiogenetico rendono i vasi pre-esistenti piu’ permeabili. Nabissi 14.")
31
Nabissi 14

32
EVENTI CELLULARI Una funzione primaria dell’infiammazione è quella di portare i leucociti nella sede della lesione e di attivarli. Le sequenze di eventi che portano al travaso sono: Attivazione dell’endotelio per indurre il reclutamento leucociti (marginazione, rotolamento, adesione) Diapedesi Migrazione Nabissi 14
 Diapedesi. Migrazione. v=WEGGMaRX8f0. Nabissi 14.")
33
Il reclutamento dei leucociti dal circolo sanguigno consiste nel rotolamento ed adesione all’endotelio vascolare a cui segue il passaggio attraverso l’endotelio e la membrana basale. Il reclutamento leucocitario è selettivo, permettendo l’accumulo di diverse popolazioni leucocitarie, reclutate con diverse cinetiche. La selettività è determinata da molecole d’adesione leucocitarie, endoteliali e da fattori chemotattici. Nabissi 14

34
transitoria (rolling).
Marginazione Con la stasi le condizioni emodinamiche cambiano ed un maggior numero di leucociti assume una posizione periferica lungo le pareti del vaso. Mediante questo processo di marginazione i leucociti rotolano lentamente lungo le pareti dei vasi aderendovi in maniera transitoria (rolling). Le pareti dei vasi vengono rivestite dai leucociti (pavimentazione). Dopo una solida adesione i leucociti inseriscono degli pseudopodi nelle giunzioni delle pareti del vaso per poi migrare al di fuori dei vasi. Nabissi 14
. Le pareti dei vasi vengono rivestite dai leucociti (pavimentazione). Dopo una solida. adesione i leucociti inseriscono degli pseudopodi nelle giunzioni delle pareti del vaso per. poi migrare al di fuori dei vasi. Nabissi 14.")
35
I recettori di adesione appartengono a quattro famiglie: Selectine
Adesione e Diapedesi L’adesione e la migrazione dei leucociti sono regolate dal legame di molecole d’adesione complementari presenti sui leucociti e sull’endotelio. I mediatori chimici influiscono su questo processo modulando l’espressione di superficie e l’avidità di queste molecole. I recettori di adesione appartengono a quattro famiglie: Selectine Integrine Glicoproteine mucina-simili Immunoglobuline Nabissi 14

36
Nabissi 14

37
LE SELECTINE: I principali iniziatori dell’adesione sono le tre selectine espresse sui leucociti (L-Selectina), sulle cellule endoteliali (P e E-Selectina) e piastrine attivate (P-selectine). Le selectine (E, P, L) si legano attraverso il loro dominio di lectina alle forme sialilate degli oligosaccaridi, presenti nelle glicoproteine mucina-simile. Nabissi 14

38
Le selectine appartengono alla famiglia di proteine chiamata LECAM (Leucocyte Endothelium Cell Adhesion Molecules) sono glicoproteine transmembrana a catena singola capaci di legare i ligandi con alta velocità di attacco ma con bassa affinità. L’espressione della selectina E viene indotta sulle cellule endoteliali da citochine infiammatorie dopo poche ore (3-4) dalla stimolo infiammatorio e diminuisce dopo circa 8-10 ore. La selectina E è riconosciuta da monociti, linfociti T di memoria e polimorfonucleati. La selectina P è sempre presente nell’endotelio ma inattiva, localizzata nei corpuscoli di Weibel e Palade; solo dopo attivazione da mediatori (es. istamina) viene esposta in membrana. Il processo di attivazione ed inattivazione è molto rapido. Nabissi 14

39
Regolazione e funzione
Molecole d’adesione Distribuzione Ligando Regolazione e funzione L-selectine Leucociti Gly CAM-1 Glicani sialilati tipo Lewis X Homing dei linfociti memoria T vergini e dei leucociti infiammatori E-selectine Endoteliali attivati da citochine Legame dei leucociti nei siti d’infiammazione P-selectine Endoteliali e piastrine Immagazzinamento intracellulare, rapida traslocazione e attivazione da istamina, trombossani e superossidi Nabissi 14

40
a1b2 integrine, fibrinogeno
Immunoglobuline Le immunoglobuline sono le molecole d’adesione presenti a livello delle cellule endoteliali che fungono da ligandi per le integrine presenti sui leucociti. Le due molecole d’adesione endoteliale sono: ICAM-1/2 (molecola d’adesione intercellulare-1) VCAM-1 (Molecole d’adesione cellulare vascolare-1) ICAM e VCAM riconosciute da linfociti e monociti, mentre i polimorfonucleati riconoscono solo ICAM-1 Molecole d’adesione Ligando ICAM-1 a1b2 integrine, fibrinogeno ICAM-2 a1b2 integrine VCAM-1 a4b1 integrine Nabissi 14
 VCAM-1 (Molecole d’adesione cellulare vascolare-1) ICAM e VCAM riconosciute da linfociti e monociti, mentre i polimorfonucleati riconoscono solo ICAM-1. Molecole d’adesione. Ligando. ICAM-1. a1b2 integrine, fibrinogeno. ICAM-2. a1b2 integrine. VCAM-1. a4b1 integrine. Nabissi 14.")
41
Integrine Per terminare il rotolamento le cellule devono formare nuovi legami addizionali più saldi. Le molecole d’adesione coinvolte in questa fase sono le integrine. Ci sono differenti sottofamiglie di integrine ed i componenti di ogni sottofamiglia esprimono una catena conservata β associata a differenti catene a. INTEGRINE b1 INTEGRINE b2 Anche conosciute come VLA (very late activation) Sono responsabili dell’adesione alle proteine della matrice extracellulare come collagene, laminina e fibronectina. Comunemente chiamate LFA-1 (leukocyte function-associated antigen-1) responsabili dell’adesione intercellulare e alla matrice. Riconoscono i ligandi ICAM-1, ICAM-2 and ICAM-3 sulle cellule endoteliali. Nabissi 14
 Sono responsabili dell’adesione alle proteine della matrice extracellulare. come collagene, laminina e fibronectina. Comunemente chiamate LFA-1 (leukocyte function-associated antigen-1) responsabili dell’adesione intercellulare e alla matrice. Riconoscono i ligandi ICAM-1, ICAM-2 and ICAM-3 sulle cellule endoteliali. Nabissi 14.")
42
'inside–out' signalling, Un attivatore intracellulare, si lega alla β-integrin tail, portando un cambio conformazionale aumentando l’affinità per componenti extracellulari (attivazione delle integrine) favorendo la migrazione . 'outside–in' signalling. Legame delle integrine al ligando extracellulare cambia la loro conformazione . L’insieme dei due eventi contribuisce al controllo della polarità cellulare, ri-arrangiameno del citoscheletro, espressione genica, proliferazione e sopravvivenza cellulare Nabissi 14

43
Nabissi 14

44
Nabissi 14

45
Nabissi 14

46
INDUZIONE DELLE MOLECOLE D’ADESIONE
Istamina, trombina e PAF (fattore attivante le piastrine) stimolano la distribuzione della P-selectina dai corpi di Weibel-Palade alla superficie cellulare. Macrofagi, mastociti, cellule endoteliali rispondono agli stimoli infettivi rilasciando citochine (TNF), IL-1 e chemochine. TNF, IL-1 stimolano le cellule endoteliali a produrre molecole di adesione (ex. E- selectine) e i leucociti le molecole complementari alla E-selectina. TNF, IL-1 stimolano anche la produzione di ICAM-1 e VCAM-1. Nabissi 14
 stimolano la distribuzione della. P-selectina dai corpi di Weibel-Palade alla superficie cellulare. Macrofagi, mastociti, cellule endoteliali rispondono agli stimoli infettivi rilasciando. citochine (TNF), IL-1 e chemochine. TNF, IL-1 stimolano le cellule endoteliali a produrre molecole di adesione (ex. E- selectine) e i leucociti le molecole complementari alla E-selectina. TNF, IL-1 stimolano anche la produzione di ICAM-1 e VCAM-1. Nabissi 14.")
47
DIAPEDESI Il passaggio dei leucociti attraverso l’endotelio è chiamato diapedesi, le cellule attivate migrano in direzione del gradiente di concentrazione chimica. Le molecole di adesione presenti nelle giunzioni intercellulari dell’endotelio coinvolte nel passaggio dei leucociti appartengono alla superfamiglia delle immunoglobuline e sono chiamate PECAM (Platelet endothelial Cell Adhesion Molecule) o CD31. Nabissi 14
 o CD31. Nabissi 14.")
48
Uno stimolo infiammatorio induce l’espressione di ICAM-1.
Dopo adesione e rolling I leucociti migrano grazie alle PECAM-1 e CD99. Le chemochine ed integrine stimolani l’espressione e le interazioni fra LFA-1 e ICAM-1. Le interazione fra PECAM-1 e CD99 contribuiscono alla trasmigrazione dei leucociti Nabissi 14

49
Il tipo di leucociti migranti varia in relazione alla progressione temporale della risposta infiammatoria ed al tipo di stimolo. Nella maggior parte delle infiammazioni acute nell’infiltrato infiammatorio predominano i neutrofili per le prime 6-24 ore, sostituiti quindi dai monociti dopo ore. I neutrofili sono i primi perché sono piu’ numerosi e sensibili alle chemochine, quindi tendono a formare legami piu’ saldi. Comunque i neutrofili hanno vita breve andando incontro ad apoptosi (formando l’essudato), mentre i monociti sopravvivono piu’ a lungo. Nabissi 14
, mentre i monociti sopravvivono piu’ a lungo. Nabissi 14.")
50
I movimenti cellulari sono orientati da un gradiente chimico.
CHEMIOTASSI I movimenti cellulari sono orientati da un gradiente chimico. Le sostanze CHEMIOATTRATTRICI possono essere i prodotti di origine batterica o componenti del sistema del complemento (C5a), prodotti della via lipossigenasica (leucotrieni) e chemochine. Come avviene l’induzione del movimento cellulare orientato? Gli agenti chemiotattici si legano a specifici recettori transmembrana accoppiati alle proteine-G (GPCR) che generano segnali intracellulari che portano alla polimerizzazione dell’actina. Questo porta a modificazioni della membrana cellulare come la formazione di filopodi e contrazione cellulare. Nabissi 14
, prodotti della via lipossigenasica (leucotrieni) e chemochine. Come avviene l’induzione del movimento cellulare orientato Gli agenti chemiotattici si legano a specifici recettori transmembrana accoppiati alle proteine-G (GPCR) che generano segnali intracellulari che portano alla polimerizzazione dell’actina. Questo porta a modificazioni della membrana cellulare come la formazione di filopodi e contrazione cellulare. v=ZUUfdP87Ssg. Nabissi 14.")
51
Nabissi 14 Le molecole chemioattrattrici formano un gradiente anche all’interno del lume vascolare. L’eparan-solfato sulla superficie dell’endotelio previene la dissoluzione del gradiente di chemochine nel plasma e fornisce una via differenziale per la migrazione leucocitaria

52
Come i neutrofili possono muoversi attraverso i numerosi compartimenti (midollo, vasi, matrice extracellulare, sito d’infiammazione) disposti in siti lontani usando un singolo gradiente? Se l’attrazione verso le pareti dell’endotelio è guidata da IL-8, come fa il leucocita ad allontanarsi dal gradiente di IL-8 e migrare verso il sito d’infiammazione? Le molecole chemiotrattrici di origine batterica (fMLP, LPS) inibiscono il segnale di IL-8, agendo sugli stessi pathways molecolari (PI3K, PTEN, ecc) Nabissi 14
 inibiscono il segnale di IL-8, agendo sugli stessi pathways molecolari (PI3K, PTEN, ecc) Nabissi 14.")
53
DIFETTI DELLA FUNZIONE LEUCOCITARIA
LAD: Leukocyte Adhesion Deficiency La caratteristica di questa patologia è la mancanza di un corretto processo di adesione dei leucociti all’endotelio con conseguente marcata leucocitosi e severe infezioni ricorrenti. Senza la fase di adesione infatti, i leucociti non sono in grado di lasciare il torrente circolatorio per migrare nel sito di infezione. Negli ultimi 20 anni sono stati identificati tre momenti nello corso dell’adesione in cui si possono avere problemi: nel rolling, nell’attivazione delle integrine, nella adesione forte, Nabissi 14

54
L’incapacità da parte dei leucociti di raggiungere il sito infiammatorio determina infezioni batteriche ricorrenti, localizzate principalmente a livello della cute e delle mucose. Queste patologie sono evidenti fin dalla nascita, in cui i pazienti possono mostrare già una infezione particolare all’ombelico con ritardo nel distacco del cordone. Nei casi più severi della malattia l’individuo muore durante l’infanzia, mentre nei casi moderati sopravvive fino all’età adulta presentando gravi gengiviti, periodontiti, ulcere cutanee, otiti, enterocoliti, perforazioni dell’ileo e dell’appendice, laringotracheiti e l’incapacità di rimarginare ferite chirurgiche. Nabissi 14

55
Nabissi 14

56
LAD I: è una malattia autosomica recessiva dovuta a mutazioni del gene ITGB2 localizzato nel cromosoma 21 che codifica per la subunità b2 (CD18) delle integrine. Segno caratteristico ritardo nel distacco del cordone ombelicale. La severità e le complicazioni dei pazienti affetti da LAD I sono direttamente correlate con il grado di deficienza della catena CD18: individui con meno dell’1% di espressione hanno una forma molto grave con morte durante l’infanzia; mentre pazienti con un’espressione che va dal 2.5 al 10% riescono a sopravvivere fino all’età adulta. Nabissi 14

57
LAD II: il difetto genetico riguarda un trasportatore del fucosio localizzato nell’apparato del Golgi. La conseguenza è una scorretta glicosilazione e quindi la mancata espressione del ligando (sLeX antigen) per le selectine, espresse dalle cellule endoteliali e quindi un rolling insufficiente. I pazienti presentano il raro gruppo sanguigno Bombay e alla nascita sono di peso e statura normali con un corretto distacco del cordone ombelicale. Più tardi con la crescita mostrano ritardo mentale, caratteristici lineamenti e statura piccola. LAD III: è molto simile alla LAD I ed il difetto genetico non è ancora esattamente conosciuto. Probabilmente si tratta del risultato di mutazioni che coinvolgono più geni responsabili dell’attivazione delle integrine b1, b2 e b3. Gli individui affetti hanno sintomi molto simili alla LAD I ma in più mostrano difetti nell’attivazione piastrinica e quindi una facilità nel sanguinamento. Nabissi 14

58
DIAGNOSI -conta delle cellule del sangue: rileva la profonda neutrofilia -FACS analisi con l’uso di anticorpi specifici per identificare la presenza o meno di CD18 e di glicoproteine fucosilate -Test di adesione -Sequenziamento genico -Diagnosi prenatale attraverso villocentesi dopo circa settimane di gestazione Nabissi 14

59
TERAPIA -Profilassi antibiotica
-per la LAD I ottimi risultati il trapianto di midollo. Risultati promettenti con terapia genica in cui cellule ematopoietiche vengono trasfettate con il gene normale della CD18. -per la LAD II ci sono risultati incoraggaianti con terapie che aumentano la formazione di glicoproteine fucosilate. -per la LAD III trasfusioni di sangue. Nabissi 14

60
ATTIVAZIONE LEUCOCITARIA
L’attivazione dei leucociti, ad agenti infettivi, comprende alcune risposte: produzione di metaboliti dell’acido arachidonico, a partire dai fosfolipidi, risultanti dall’attivazione della fosfolipasi A2 dovuto ad un aumento del calcio intracellulare ed altri segnali. - secrezione di citochine che regolano ed amplificano la reazione infiammatoria - modulazione delle molecole d’adesione leucocitarie mediante il rilascio di citochine che aumentano il legame alle molecole endoteliali degranulazione e secrezione di enzimi lisosomiali ad attivazione della cascata ossidativa (fagocitosi) Nabissi 14
 Nabissi 14.")
61
Fattori chemotattici I fattori chemotattici svolgono una duplice azione, in quanto stimolano la migrazione dei leucociti nel sito d’infiammazione ed aumentano l’adesione leucocitaria sulla membrana endoteliale. Gli stimoli chemotattici possono essere di natura batterica, lipidica o proteica. Origine batterica: fMLP (tripeptide formilato Met-Leu-Phe) Origine lipidica: (PAF, LTB4) Origine proteica: C5a, C3a, IL-5, M-CSF (Macrophage colony-stimulating factor ), Chemochine) Nabissi 14
 Origine lipidica: (PAF, LTB4) Origine proteica: C5a, C3a, IL-5, M-CSF (Macrophage colony-stimulating factor ), Chemochine) Nabissi 14.")
62
Chemochine Le chemochine, chemotattic cytochine sono una famiglia di piccole proteine che agiscono principalmente come fattori chemiotattici di attrazione specifica per i leucociti. Attualmente sono state identificate circa 50 chemochine e circa 20 recettori per le chemochine. Le chemochine sono classificate in quattro gruppi in relazione alla disposizione dei residui di cisteina nelle proteine mature: Le chemochine C-C: presentano i primi due residui di cisteina adiacenti. Includono la proteina chemiotattica per i monociti, la proteina infiammatoria dei macrofagi; generalmente attraggono monociti, eosinofili, basofili e linfociti, nei siti d’infiammazione cronica. La forma attiva delle CC spesso comprende dimeri o tetrameri Nabissi 14

63
Nabissi 14

64
Vengono indotte da prodotti microbici e da IL-1 e TNF.
Le chemochine C-X-C: hanno un residuo aminoacidico che separa le due cisteine, come IL-8 (CXCL8) che attrae i leucociti polimorfonucleati nel sito dell’infiammazione acuta. Vengono indotte da prodotti microbici e da IL-1 e TNF. Nabissi 14
 che attrae i leucociti polimorfonucleati nel sito dell’infiammazione acuta. Vengono indotte da prodotti microbici e da IL-1 e TNF. Nabissi 14.")
65
Le chemochine CX3C: contengono tre aminoacidi fra i due residui di cisteina, Il solo menbro di questa classe è chiamato FRACTALCHINA. Questa chemochina esiste in due forme, una legata alla superficie cellulare ed indotta da citochine infiammatorie (TNFa) promuove l’adesione di monociti e linfociti T e una solubile che ha un potente effetto chemiotattico d’attrazione per le stesse cellule. La linfotactina (XCL1) è il solo membro della quarta famiglia e contiene un singolo residuo di cisteina Nabissi 14
 è il solo membro della quarta famiglia e contiene un singolo residuo di cisteina. Nabissi 14.")
66
extracellulare (tumori/metastasi).
Comunque in generale le chemochine agiscono stimolando il reclutamento dei leucociti nell’infiammazione e controllano la normale migrazione cellulare attraverso vari tessuti. Inoltre possono anche indurre degranulazione, produzione di ROS e attivare la trascrizione dei geni per i recettori delle chemochine e di geni per enzimi coinvolti nella degradazione della matrice extracellulare (tumori/metastasi). Nabissi 14
. Nabissi 14.")
67
Patologie infiammatorie e chemochine
Le chemochine sono state correlate a diversi tipi di malattie con una prominente componente infiammatoria. Alti livelli di chemochine CC (CCL2, CCL3, CCL5) sono riscontrate in pazienti con artrite reumatoide in coincidenza con il reclutamento di monociti e linfociti T nei tessuti sinoviali. La chemochina CCL11 ed il suo recettore CCR3 nell’asma, contribuiscono al reclutamento di eosinofili nel polmone. Nabissi 14
 sono riscontrate in pazienti con artrite reumatoide in coincidenza con il reclutamento di monociti e linfociti T nei tessuti sinoviali. La chemochina CCL11 ed il suo recettore CCR3 nell’asma, contribuiscono al reclutamento di eosinofili nel polmone. Nabissi 14.")
68
Recettori di membrana dell’immunità innata
Recettori per i peptidi formilati, formati da 7 domini transmembrana accoppiati a proteine G. Riconoscono peptidi che iniziano con formilmetionina, caratteristica delle proteine batteriche e proteine self come la b-amiloide o la lipotossina (presenti nel plasma in presenza d’infiammazione). Recettori per il mannosio o lectinici riconoscono strutture ricche di mannosio, come le membrane batteriche. Recettori SCAVENGER che riconoscono strutture batteriche e lipoproteine alterate self, come nel processo dell’arteriosclerosi in cui l’infiammazione ha un ruolo importante. TOLL receptor che hanno un dominio costituito da leucine ripetute Nabissi 14
. Recettori per il mannosio o lectinici riconoscono strutture ricche di mannosio, come le membrane batteriche. Recettori SCAVENGER che riconoscono strutture batteriche e lipoproteine alterate self, come nel processo dell’arteriosclerosi in cui l’infiammazione ha un ruolo importante. TOLL receptor che hanno un dominio costituito da leucine ripetute. Nabissi 14.")
69
I leucociti esprimono diversi tipi di recettori di membrana che sono coinvolti nella loro attivazione Nabissi 14

70
I recettori scavenger nei macrofagi sono responsabili delll’uptake di cellule apoptotiche, lipoproteine modificate ed altri ligandi di origine batterica. Nabissi 14

71
Nabissi 14

72
I Toll sono parte di complessi recettoriali che riconoscono diverse componenti batteriche o molecole dell’organismo danneggiato. Inoltre possono cooperare fra loro formando eterodimeri e aumentando le capacità di riconoscimento. TLR 7 e 8 di cui non si conosce il ligando, si attivano riconoscendo farmaci antivirali. Nell’insieme i TOLL costituiscono un gruppo di recettori in grado di riconoscere componenti virali, batterici, parassiti e molecole alterate dell’organismo ( LPS, Heat Shock Protein) CD14 è un componente recettoriale dell’immunità innata che si trova in forma libera o legato alla membrana dei fagociti. Il suo ruolo è quello di presentare endotossine (come LPS) a TRL4, rendendo responsive cellule mancanti di CD14 ma esprimenti TRL4 a prodotti microbici ed attivando cosi’ il processo infiammatorio, come nelle cellule endoteliali che esprimono TRL4 Nabissi 14
 CD14 è un componente recettoriale dell’immunità innata che si trova in forma libera o legato alla membrana dei fagociti. Il suo ruolo è quello di presentare endotossine (come LPS) a TRL4, rendendo responsive cellule mancanti di CD14 ma esprimenti TRL4 a prodotti microbici ed attivando cosi’ il processo infiammatorio, come nelle cellule endoteliali che esprimono TRL4. Nabissi 14.")
73
I Toll sono costituiti, nella parte citoplasmatica da un dominio TIR (Toll/IL-1 receptor domain), che agisce da sensore citoplasmatico che attiva una complessa cascata d’attivazione del segnale. L’attivazione di TIR, che è un dominio d’interazione proteina-proteina recluta proteine adattatrici, come MyD88 che è costituita da un dominio TIR e uno DD (Death Domain). Grazie a DD vengono reclutate chinasi, denominate IRAK che fosforilano un’adattatore (TRAF6) che recluta diverse chinasi. Questa cascata porta all’attivazione di NfkB o della via della chinasi da stress JNK e p38. Nabissi 14

74
(enzima che produce NO), COX-2 (via delle lipossigenasi).
Il fattore trascrizionale NFkB attiva un programma pro-infiammatorio. I membri principali della famiglia NFkB sono p50 e p65 che sono complessati a livello citoplasmatico con il loro inibitore IkB. L’attivazione dei recettori che hanno come via quella di NFkB, porta all’attivazione di chinasi (IKK1 e IKK2) che fosforilano IkB, dopo essere state attivate da serin treonin chinasi TAK che fosforilano e distaccano l’ inibitore delle IKK (NEMO). p50 e p65 attivate migrano nel nucleo ed attivano la trascrizione di geni pro-infiammatori: (IL-1, IL-6), molecole d’adesione endoteliale, NO sintetasi (enzima che produce NO), COX-2 (via delle lipossigenasi). Nabissi 14
 che fosforilano IkB, dopo essere state attivate da serin treonin chinasi TAK che fosforilano e distaccano l’ inibitore delle IKK (NEMO). p50 e p65 attivate migrano nel nucleo ed attivano la trascrizione di geni pro-infiammatori: (IL-1, IL-6), molecole d’adesione endoteliale, NO sintetasi. (enzima che produce NO), COX-2 (via delle lipossigenasi). Nabissi 14.")
75
Nabissi 14

76
IMMUNITA’ INNATA Recettori per le citochine . Ad esempio INF-g, la principale citochina che attiva i macrofagi, prodotta dai linfociti attiva i macrofagi e viene secreta dalle cellule natural killer (NK) nella risposta immunitaria innata e dai linfociti T attivati dall’antigene nella risposta immunitaria acquisita. I recettori per le opsonine , promuovono la fagocitosi dei microbi rivestiti da varie proteine trasmettendo i segnali che attivano i fagociti. La OPSONIZZAZIONE è il processo che riveste le cellule microbiche di opsonine (anticorpi, proteine del complemento, lectine). Uno dei processi principali di opsonizzazione permette di rivestire le particelle batteriche con anticorpi IgG, i quali sono poi riconosciuti dal recettore ad alta affinità Fcg (FcgRI) presenti nel lato esterno della membrana dei fagociti. Un altro processo di oposonizzazione comprende i frammenti della proteina del complemento C3, che si legano ai microbi e li presentano ai fagociti che esprimono il recettore CR1. Nabissi 14
 nella risposta immunitaria innata e dai linfociti T attivati dall’antigene nella risposta immunitaria acquisita. I recettori per le opsonine , promuovono la fagocitosi dei microbi rivestiti da varie proteine trasmettendo i segnali che attivano i fagociti. La OPSONIZZAZIONE è il processo che riveste le cellule microbiche di opsonine (anticorpi, proteine del complemento, lectine). Uno dei processi principali di opsonizzazione permette di rivestire le particelle batteriche con anticorpi IgG, i quali sono poi riconosciuti dal recettore ad alta affinità Fcg (FcgRI) presenti nel lato esterno della membrana dei fagociti. Un altro processo di oposonizzazione comprende i frammenti della proteina del complemento C3, che si legano ai microbi e li presentano ai fagociti che esprimono il recettore CR1. Nabissi 14.")
77
OPSONINE Le opsonine principali sono le immunoglobuline G (IgG) che vengono riconosciute dai recettori per la porzione cristallizzabile (FcgR) presente sui fagociti e le proteine derivanti dall’attivazione della cascata del complemento (C3b, C3bi, C1q) che vengono riconosciute da recettori specifici (CR1, 2, 3). Nabissi 14
 che vengono riconosciute dai recettori per la porzione cristallizzabile (FcgR) presente sui fagociti e le proteine derivanti dall’attivazione della cascata del complemento (C3b, C3bi, C1q) che vengono riconosciute da recettori specifici (CR1, 2, 3). Nabissi 14.")
78
FAGOCITOSI Nabissi 14

79
I LISOSOMI Il lisosoma è un organulo cellulare contenente enzimi digestivi capaci di degradare molte macromolecole biologiche permettono di digerire sia macromolecole introdotte dall’esterno che quelle danneggiate interne. Gli enzimi lisosomiali sono racchiusi in una vescicola membranosa rivestita di clatrina e rilasciati dalla porzione trans del Golgi Nabissi 14

80
LESIONI LEUCOCITARIE I leucociti liberano i prodotti antimicrobici anche nello spazio extracellulare (lisozima, ROI, metaboliti dell’AA (prostaglandine, leucotrieni)). Siccome queste sostanze possono causare danno endoteliale, se l’attivazione leucocitaria persiste, lo stesso infiltrato puo’ risultare dannoso. Rigurgito durante l’ingestione: quando il vacuolo d’ingestione rimane momentaneamente aperto prima della fusione fago-lisosoma Fagocitosi ostacolata: adesione dei leucociti a immunocomplessi depositati su superfici piatte attiva i leucociti ma non ne permette la fagocitosi. Quindi gli enzimi lisosomiali sono rilasciati nell’ambiente extracellulare. Nabissi 14
). Siccome queste sostanze possono causare danno endoteliale, se l’attivazione leucocitaria persiste, lo stesso infiltrato puo’ risultare dannoso. Rigurgito durante l’ingestione: quando il vacuolo d’ingestione rimane momentaneamente aperto prima della fusione fago-lisosoma. Fagocitosi ostacolata: adesione dei leucociti a immunocomplessi depositati su superfici piatte attiva i leucociti ma non ne permette la fagocitosi. Quindi gli enzimi lisosomiali sono rilasciati nell’ambiente extracellulare. Nabissi 14.")
81
UCCISIONE E DEGRADAZIONE
L’uccisione avviene principalmente mediante meccanismi ossigeno-dipendenti, con un improvviso aumento del consumo di ossigeno, della glicogenolisi, ossidazione del glucosio e produzione di intermedi liberi dell’ossigeno (ROI). La formazione dei ROI è dovuta all’attivazione della NADPH ossidasi che ossida il NADPH (nicotinamide-adenin-dinucleotide fosfato ridotto) riducendo l’ossigeno a ANIONE SUPEROSSIDO (O2-). Il O2- viene poi convertito in perossido d’idrogeno (H2O2) che puo’ essere ulteriormente ridotto a radicale idrossile altamente reattivo (OH-). Altri sistemi di degradazione coinvolgono la MIELOPEROSSIDASI (nei neutrofili) che in presenza di Cl- converte H2O2 in ipoclorito (HOCl) che distrugge in batteri mediante ALOGENAZIONE. Nabissi 14
. La formazione dei ROI è dovuta all’attivazione della NADPH ossidasi che ossida il NADPH (nicotinamide-adenin-dinucleotide fosfato ridotto) riducendo l’ossigeno a ANIONE SUPEROSSIDO (O2-). Il O2- viene poi convertito in perossido d’idrogeno (H2O2) che puo’ essere ulteriormente ridotto a radicale idrossile altamente reattivo (OH-). Altri sistemi di degradazione coinvolgono la MIELOPEROSSIDASI (nei neutrofili) che in presenza di Cl- converte H2O2 in ipoclorito (HOCl) che distrugge in batteri mediante ALOGENAZIONE. Nabissi 14.")
82
Nabissi 14

83
Nabissi 14

84
Azione dei ROS Gli effetti delle specie reattive sono molti, ma i danno cellulari principali sono: la perossidazione lipidica della membrana, le alterazioni ossidative delle proteine ed il danno al DNA. Il danno ossidativo ai lipidi di membrana avviene quando i doppi legami presenti negli acidi grassi insaturi vengono attaccati dai radicali liberi dell’ossigeno, generando perossidi che attraverso la reazione a catena producono danni alla membrana cellulare e da quelle degli organelli citoplasmatici. Le alterazioni ossidative delle proteine indotte da radicali liberi comportano la formazione di legami crociati proteina-proteina, causandone la frammentazione, mentre il danno al DNA comporta la dimerizzazione delle timine adiacenti sullo stesso filamento con rottura del filamento di DNA. Nabissi 14

85
Nabissi 14

86
RADICALI LIBERI DELL’OSSIGENO
Le specie reattive sono l’anione superossido (O2-), il perossido d’idrogeno (H2O2) ed il radicale idrossilico (OH-) e tali specie possono combinarsi con NO per formare altri intermedi reattivi dell’azoto (RNS). Il rilascio extracellulare di queste specie reattive puo’ aumentare l’espressione di chemochine, citochine e molecole d’adesione endoteliali nei leucociti, amplificando la cascata di eventi dell’infiammazione. Nabissi 14
, il perossido d’idrogeno (H2O2) ed il radicale idrossilico (OH-) e tali specie possono combinarsi con NO per formare altri intermedi reattivi dell’azoto (RNS). Il rilascio extracellulare di queste specie reattive puo’ aumentare l’espressione di chemochine, citochine e molecole d’adesione endoteliali nei leucociti, amplificando la cascata di eventi dell’infiammazione. Nabissi 14.")
87
Nabissi 14

88
RADICALI LIBERI DELL’OSSIGENO meccanismi antiossidanti
Un eccesso di produzione di questi mediatori puo’ chiaramente risultare dannosa per la cellula ospite, anche se sono comunque presenti dei meccanismi enzimatici antiossidanti, che sono: Ceruloplasmina: proteina sierica contenente rame Transferina: frazione sierica priva di ferro Superossido Dismutasi Catalasi Glutadione perossidasi: altro potente detossificatore del H2O2 Quindi il ruolo dei radicali liberi dell’ossigeno dipende dall’equilibrio tra la produzione e l’inattivazione di tali metaboliti. Nabissi 14

89
INIBITORI DEI ROS Gli inibitori primari comprendono sostanze chelanti (es. fenoli, vitamina E, A) l’attività di alcuni enzimi presenti all’interno della cellula (catalasi, glutadione-perossidasi, superossido dismutasi), i secondari comprendono sostanze che reagiscono con radicali propagatori della catena attraverso reazioni di trasferimento d’ idrogeno, elettroni o addizione con il risultato di terminare la catena di produzione di radicali liberi o con la formazione di radicali meno reattivi. La catalasi converte H2O2 in O2 + H2O ed è presente all’interno dei perossisomi, la superossido dismutasi è presente in molti tipi di cellule e converte l’anione superossido in H2O2. La glutadione perossidasi catalizza la degradazione dei radicali liberi utilizzando glutadione ridotto (GSH), convertendo cosi’ H2O2 in acqua e GSH in glutadione ossidato (GSSG). Importante è il rapporto intracellulare di GSSG e GSH in quando indica lo stato ossidativo della cellula e la capacità cellulare di detossificare Nabissi 14
 l’attività di alcuni enzimi presenti all’interno della cellula (catalasi, glutadione-perossidasi, superossido dismutasi), i secondari comprendono sostanze che reagiscono con radicali propagatori della catena attraverso reazioni di trasferimento d’ idrogeno, elettroni o addizione con il risultato di terminare la catena di produzione di radicali liberi o con la formazione di radicali meno reattivi. La catalasi converte H2O2 in O2 + H2O ed è presente all’interno dei perossisomi, la superossido dismutasi è presente in molti tipi di cellule e converte l’anione superossido in H2O2. La glutadione perossidasi catalizza la degradazione dei radicali liberi utilizzando glutadione ridotto (GSH), convertendo cosi’ H2O2 in acqua e GSH in glutadione ossidato (GSSG). Importante è il rapporto intracellulare di GSSG e GSH in quando indica lo stato ossidativo della cellula e la capacità cellulare di detossificare Nabissi 14.")
90
Nabissi 14

Presentazioni simili


 Costituito da una parte liquida: plasma e.>")

 SPECIFICA/SELETTIVA:>")





>")

 Pontieri - Elementi di Patologia Generale (ed. Piccin) Majno-Joris - Cellule,>")






