Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Matrice ExtraCellulare
Introduzione

2
Take-home message Il comportamento cellulare (metabolismo, proliferazione, differenziamento o staminalità, migrazione, apoptosi) non dipende solo da caratteristiche intrinseche delle cellule, ma è strettamente condizionato dall’interazione tra cellule, matrice e microambiente cellulare.
 non dipende solo da caratteristiche intrinseche delle cellule, ma è strettamente condizionato dall’interazione tra cellule, matrice e microambiente cellulare.")
3
Extracellular Matrix (ECM)
A collection of proteins and carbohydrates produced by cells that forms a matrix outside the cell a.k.a. connective tissue, interstitium The extracellular matrix is interesting to biotech because it: Is involved in disease processes, including atherosclerosis, cancer, arthritis, lupus… Is a major impediment to gene therapy Interactions of the matrix with cell surface receptors influence cell behavior and gene expression. Creating artificial matrices is a big part of tissue engineering, and one of the biggest challenges.

4
Lukashev ME, Werb Z. ECM signalling: orchestrating cell behaviour and misbehaviour. Trends Cell Biol. 8: , 1998.

5
“Matrice extracellulare e matrichine, segnali criptici fondamentali per il dialogo locale cellula-matrice” (1) La matrice extracellulare è una struttura dinamica ricca di proteine multimodulari altamente glicosilate e di polisaccaridi che sorreggono meccanicamente e orientano le cellule che vi poggiano ma soprattutto modulano e pilotano ad ogni momento il loro comportamento (metabolismo, proliferazione, differenziamento o staminalità, migrazione, apoptosi) A loro volta le cellule condizionano la composizione e dinamica della matrice.
 A loro volta le cellule condizionano la composizione e dinamica della matrice.")
6
Matrisoma Elenco delle proteine che costituiscono la matrice extracellulare e di quelle che contribuiscono alle matrici in diverse situazioni. Le proteine della matrice tipicamente contengono ripetizioni di un insieme caratteristico di domini: LamG (Laminin G), TSPN (thrombospondin), FN3 (fibronectin 3), VWA (von Willebrandt A), Ig (immunoglobulin), EGF (epidermal growth factor), prodomini del collagene, ecc. Molti di questi domini non sono specifici delle proteine della MEC, ma la loro disposizione è unica e caratteristica. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
, TSPN (thrombospondin), FN3 (fibronectin 3), VWA (von Willebrandt A), Ig (immunoglobulin), EGF (epidermal growth factor), prodomini del collagene, ecc. Molti di questi domini non sono specifici delle proteine della MEC, ma la loro disposizione è unica e caratteristica. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
7
“Matrice extracellulare e matrichine, segnali criptici fondamentali per il dialogo locale cellula-matrice” (2) La proteolisi della matrice in situazioni normali e patologiche induce varchi che facilitano la migrazione cellulare e provoca il rilascio in soluzione nel liquido interstiziale di frammenti peptidici o la comparsa sulla superficie proteica di moduli precedentemente situati in posizione criptica, che sono riconosciuti da integrine sulla superficie cellulare comportandosi da fattori di crescita o di differenziamento. I proteoglicani e l’acido ialuronico permettono una concentrazione localizzata di tali segnali contribuendo alla diversificazione del microambiente cellulare.

8
Matrice extracellulare, introduzione (1)
Nonostante la MEC sia composta soltanto da cinque classi di macromolecole -collageni, elastina, acido ialuronico, proteoglicani, glicoproteine - essa può assumere una ricca varietà di forme con proprietà meccaniche altamente differenziate.

10
Matrice extracellulare

12
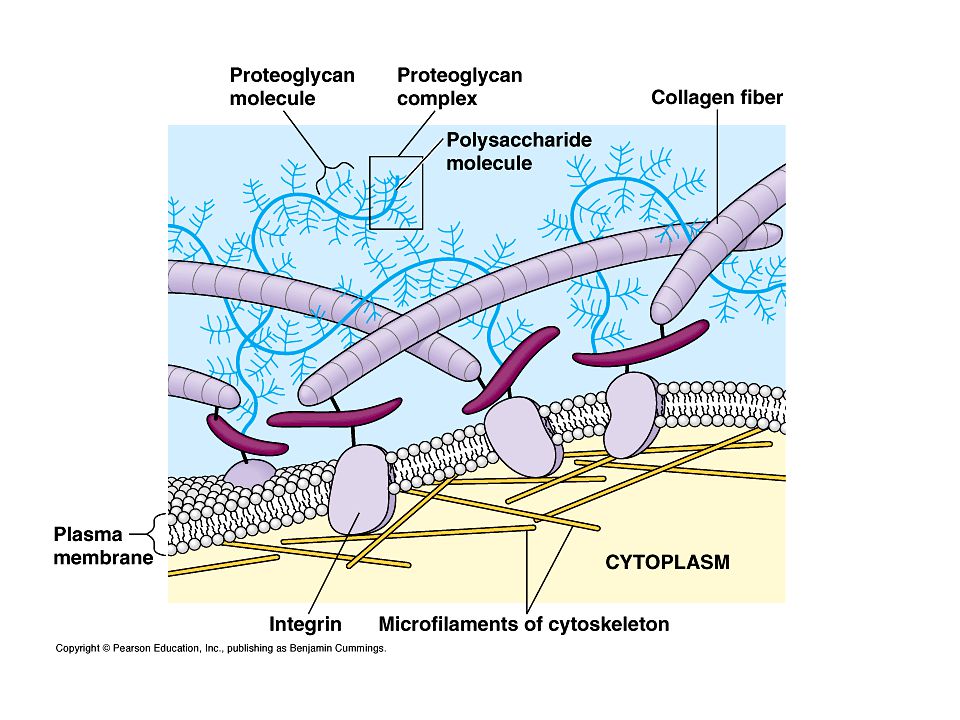
Matrice Extracellulare Materiale acellulare che circonda le cellule
Struttura della MEC: Fibre: collageno ed elastina, che forniscono resistenza e flessibilità Proteoglicani: complessi proteine- polisaccaridi che forniscono una matrice voluminosa Glicoproteine adesive: “incollano le cellule” e la MEC, ad es. fibronectina e laminina. Funzione Sostegno meccanico per le cellule e per il tessuto Integrazione delle cellule in tessuti Influenzamento della forma e il movimento della cellule Influenzamento dello sviluppo e il differenziamento delle cellule Coordinamento delle funzioni cellulari mediante segnalamento attraverso recettori di adesione (e recettori associati) Riserva di molecole di segnalamento extracellulare. Legame con proteasi della matrice e loro inibitori
 Riserva di molecole di segnalamento extracellulare. Legame con proteasi della matrice e loro inibitori.")
13
MEC: Macromolecole secrete localmente da cellule specializzate
Fibroblasti nei tessuti connettivi Condroblasti, osteoblasti: tessuti connettivi specializzati Cellule epiteliali e fibroblasti: lamina basale

14
Matrice Extracellulare, MEC
E’ costituita da proteine e polisaccaridi secreti da quasi tutti i tipi cellulari (eccezione: eritrociti) Riempie gli spazi tra le cellule Collega tra di loro cellule e tessuti. Una sottile lamina basale sostiene meccanicamente e metabolicamente cellule epitaliali, cellule muscolari, cellule adipose e i nervi periferici: LAMINA BASALE E’ particolarmente abbondante nei tessuti connettivi.
 Riempie gli spazi tra le cellule. Collega tra di loro cellule e tessuti. Una sottile lamina basale sostiene meccanicamente e metabolicamente cellule epitaliali, cellule muscolari, cellule adipose e i nervi periferici: LAMINA BASALE. E’ particolarmente abbondante nei tessuti connettivi.")
15
Matrice Extracellulare
Gran varietà di polisaccaridi e (glico)proteine) altamente versatili che vengono secreti localmente e si assemblano in una rete organizzata Variazioni in: quantità relative macromolecole della matrice modo di organizzarsi Stupefacente diversità di forme, adattate alle richieste funzionali dei vari tessuti: Calcificazione: strutture dure e compatte delle ossa o dei denti Matrice trasparente: cornea Strutture cordoniformi: tendini Lamina di controllo della permeabilità: “lamina” basale
proteine) altamente versatili che vengono secreti localmente e si assemblano in una rete organizzata. Variazioni in: quantità relative macromolecole della matrice. modo di organizzarsi. Stupefacente diversità di forme, adattate alle richieste funzionali dei vari tessuti: Calcificazione: strutture dure e compatte delle ossa o dei denti. Matrice trasparente: cornea. Strutture cordoniformi: tendini. Lamina di controllo della permeabilità: lamina basale.")
16
MEC: RUOLO Impalcatura relativamente inerte per stabilizzare la struttura fisica tessuti??? NO!! Ruolo attivo e complesso di regolazione del comportamento delle cellule Influenza su: Sviluppo Migrazione cellulare Proliferazione Apoptosi Forma Funzioni metaboliche

17
MACROMOLECOLE EXTRACELLULARI (1)
Polisaccaridi di tipo glicosaminoglicani (GAGs) di solito legati covalentemente a proteine formando proteoglicani. Proteine fibrose: Predominantemente strutturali: collagene, elastina Predominantemente adesive: es: Fibronectina Laminina
 di solito legati covalentemente a proteine formando proteoglicani. Proteine fibrose: Predominantemente strutturali: collagene, elastina. Predominantemente adesive: es: Fibronectina. Laminina.")
18
MACROMOLECOLE EXTRACELLULARI (2)
Fibre collagene: rafforzano e aiutano ad organizzare la matrice. Fibre di elastina: conferiscono resilienza (capacità di riprendere la forma primitiva dopo deformazione) Proteine multiadesive: aiutano le cellule ad aderire alla matrice
 Proteine multiadesive: aiutano le cellule ad aderire alla matrice.")
19
GAGs & Proteoglicani “Sostanza fondamentale” altamente idratata tipo gel in cui sono incluse le proteine fibrose: Fase acquosa del gel di polisaccaridi permette la diffusione di nutrienti, metaboliti, cataboliti e ormoni tra sangue e cellule.

20
University of Virginia
La matrice idratata La MEC è essenzialmente un gel (solido fibroso permeato da un liquido viscoso) creato dalle cellule che vi aderiscono. Fibre strutturali e Elastina Matrice idratata Fluido extracellulare Proteoglicani Molecole adesive Fibronectina Laminina University of Virginia
 creato dalle cellule che vi aderiscono. Fibre strutturali e Elastina. Matrice idratata. Fluido extracellulare. Proteoglicani. Molecole adesive. Fibronectina. Laminina. University of Virginia.")
21
Liquido interstiziale
Il liquido interstiziale è fondamentale per mantenere l’omeostasi fra le aree intracellulari e extracellulari. E’ composto da un vettore acquoso che contiene soprattutto acidi grassi, aminoacidi, zuccheri, coenzimi, messaggeri chimici quali citochine, ormoni, neurotrasmettitori, sali minerali, gas in soluzione (ad es. O2, CO2) e prodotti di scarto.
 e prodotti di scarto. LP=102.")
22
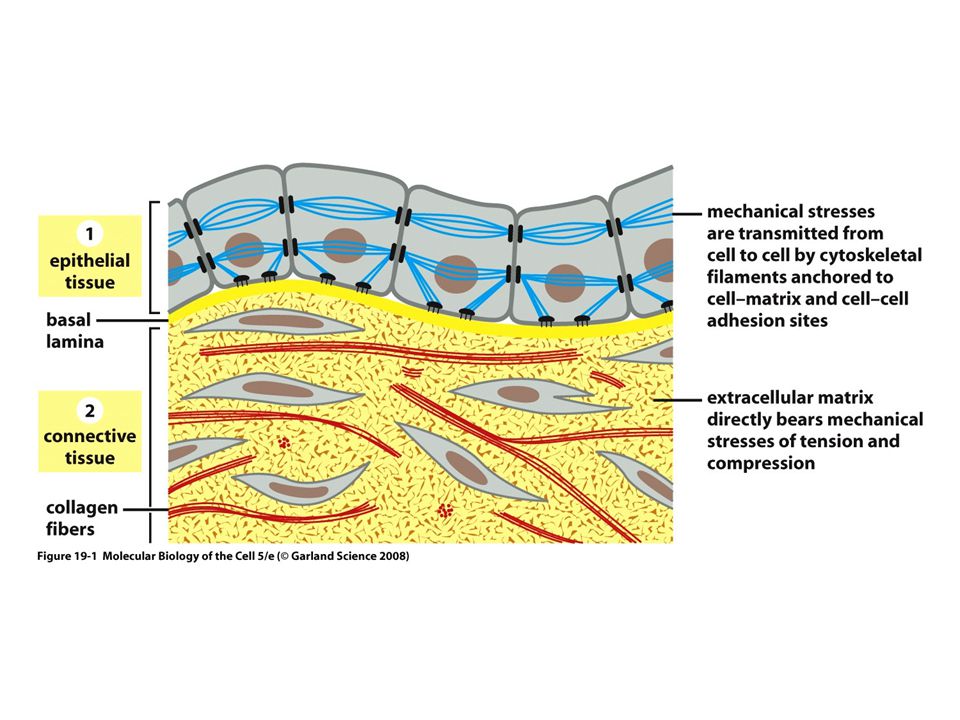
Meccanotrasduzione e biotensegrità
Mediante il processo di meccanotrasduzione, le cellule sono in grado di convertire alterazioni di tipo meccanico in alterazioni chimiche o genetiche. Le cellule mantengono la loro morfologia e funzione mediante un sistema di tensione integrato denominato tensegrità.

24
All’interno delle cellule I microtubuli costituiscono le strutture di compressione, e l’actina, strettamente associate ai filament di miosina, le strutture di tensione. La tensione meccanica, generata dai movimenti ,è trasmessa mediante pressione alla MEC che a sua volta trasferisce i movimenti alle cellule mediante le integrine. Il citoscheletro trasduce queste forze in segnali chimici e stimoli meccanici che sono trasferiti al nucleo tramite i filamenti intermedi che sono collegati a proteine dette nesprine e Sun. Queste a loro volta communicano con la lamina nucleare e la lamina con il DNA. Il nucleo stesso ha un sistema di tensegrità intriseca e attiva la proliferazione, il metabolismo, il differenziamento o l’apoptosi in risposta a tali stimoli meccanici

25
Biotensegrità La morfologia appiattita o arrotondata degli elementi cellulari e la struttura tridimensionale dei patterns tissutali formando ghiandole, alveoli, dotti e papillae dipendono dalla rigidità o flessibilità della membrana basale, nonchè del suo movimento coordinato. Noguera R, Nieto OA, Tadeo I, Fariñas F, Alvaro T. Extracellular matrix, biotensegrity and tumor microenvironment. An update and overview. Histol Histopathol Jun;27(6):
:")
28
Lamina basale

29
ECM

31
Collageni (Introd. 1) Si trovano in tutti i Metazoi e forniscono forza strutturale a tutte le matrice extracellulari, incluso le forti fibre dei tendini, le matrici organiche delle ossa e cartilagine, gli strati laminari delle membrane basali, la matrice viscosa dell’umor vitreo e la matrice interstiziale del derma e delle capsule intorno agli organi. Tipica dei collageni sono le ripetizioni delle triplette Gly-X-Y in cui X è frequentemente la prolina e Y frequentemente la 4- idrossiprolina. Questa struttura ripetitiva forma «coiled coils» trimerici stabili, a forma di bastoncello, che possono avere lunghezza variabile. Le subunità dei collageni si assemblano come omotrimeri o come un insieme ristretto di eterotrimeri. Prolina Idrossiprolina Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
:a")
32
Collageni (Introd. 2) Il tipo I dei collageni delle ossa e tendini contiene quasi interamente una lunga (~1000 aa) e rigida tripla elica di collagene (più i prodomini terminali non collagenosi che vengono rimossi durante il processo di biosintesi della proteina). I trimeri a bastoncello si associano in oligomeri di ordine superiore e fibrille che possono creare ulteriormente legami incrociati («cross-links») mediante diverse reazioni enzimatiche e non che conferiscono una considerevole resistenza strutturale. Molti altri tipi di collagene hanno interruzioni nella struttura ripetuta Gly-X-Y, che introducono flessibilità alle molecole. Tutti i geni dei collageni codificano per domini non-collagenosi addizionali, alcuni dei quali sono i caratteristici pro-domini B- e C-terminali mentre gli altri sono domini condivisi con altre proteine della MEC e trattenuti nella proteina matura. Questi domini aggiuntivi conferiscono affinità di legame specifiche, permettendo alle molecole dei collageni di interagire le une con le altre e di assemblare le varie strutture. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 e rigida tripla elica di collagene (più i prodomini terminali non collagenosi che vengono rimossi durante il processo di biosintesi della proteina). I trimeri a bastoncello si associano in oligomeri di ordine superiore e fibrille che possono creare ulteriormente legami incrociati («cross-links») mediante diverse reazioni enzimatiche e non che conferiscono una considerevole resistenza strutturale. Molti altri tipi di collagene hanno interruzioni nella struttura ripetuta Gly-X-Y, che introducono flessibilità alle molecole. Tutti i geni dei collageni codificano per domini non-collagenosi addizionali, alcuni dei quali sono i caratteristici pro-domini B- e C-terminali mentre gli altri sono domini condivisi con altre proteine della MEC e trattenuti nella proteina matura. Questi domini aggiuntivi conferiscono affinità di legame specifiche, permettendo alle molecole dei collageni di interagire le une con le altre e di assemblare le varie strutture. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
33
Collageni (Introd. 3) Fra le proteine tipo collagene o affini ai collageni, alcune sono proteine di membrana (es. coll XVII degli emidesmosomi); Altre (ad es. la componente c1q del complemento e proteine simili) vengono secrete, ma la loro funzione principale non coinvolge la MEC e non sono considerate componenti del «matrisoma». Tuttavia, altre ancora (ad es. il dominio tipo collagene della acetilcolinesterasi) servono per ancorare altre proteine della MEC). Ancora altre (ad es. le proteine contenenti domini detti EMI) sono vere proteine della MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 vengono secrete, ma la loro funzione principale non coinvolge la MEC e non sono considerate componenti del «matrisoma». Tuttavia, altre ancora (ad es. il dominio tipo collagene della acetilcolinesterasi) servono per ancorare altre proteine della MEC). Ancora altre (ad es. le proteine contenenti domini detti EMI) sono vere proteine della MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
34
Esempi di strutture multidominio dei collageni
Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
:a")
35
Didascalia figura precedente (Esempi di strutture di collagene)
Il collagene di tipo I è un collagene fibrillare con un dominio collagenoso continuo di circa 100 AA (fucsia) comprendente ripetizioni Gy-X-Y che formano una triplice elica. E’ codificato da esoni multipli (notare linee verticali) che sono varianti di un esone primordiale che codifica sei di tali ripetizioni. Il dominio collagenoso è fiancheggiato da domini N- e C-terminali non-colagenosi, rimossi per proteolisi per permettere la fibrillogenesi del collagene maturo. I domini VWC in questo e negli altri collageni possono subire splicing alternativo e legare “bone morphogenetic proteins” (BMPs). Il collagene di tipo IX è un collagene “Fibril-associated” (FACIT); le interruzioni nel cominio collagene permettono il piegamento. Questo e altri collageni FACIT si associano con i collageni fibrillari e i loro N-terminali protrudono dalle fibrille e presumibilmente fungono da domini leganti proteine. Il collagene di tipo VI è un eterodimero di tre subunità correlate, una delle quali è molto più lunga e forma una testa globulare ad ogni estremità. Domini vWA sono di solito domini leganti proteine che probabilmente permettono interazioni con altre proteine durante la formazione di corte fibrille del coll VI. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 comprendente ripetizioni Gy-X-Y che formano una triplice elica. E’ codificato da esoni multipli (notare linee verticali) che sono varianti di un esone primordiale che codifica sei di tali ripetizioni. Il dominio collagenoso è fiancheggiato da domini N- e C-terminali non-colagenosi, rimossi per proteolisi per permettere la fibrillogenesi del collagene maturo. I domini VWC in questo e negli altri collageni possono subire splicing alternativo e legare bone morphogenetic proteins (BMPs). Il collagene di tipo IX è un collagene Fibril-associated (FACIT); le interruzioni nel cominio collagene permettono il piegamento. Questo e altri collageni FACIT si associano con i collageni fibrillari e i loro N-terminali protrudono dalle fibrille e presumibilmente fungono da domini leganti proteine. Il collagene di tipo VI è un eterodimero di tre subunità correlate, una delle quali è molto più lunga e forma una testa globulare ad ogni estremità. Domini vWA sono di solito domini leganti proteine che probabilmente permettono interazioni con altre proteine durante la formazione di corte fibrille del coll VI. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
36
ECM - GLICOSAMINOGLICANI

37
PROTEOGLICANI

38
ECM - PROTEOGLICANI

39
Proteoglicani (Introd. 1)
Possono essere considerati come glicoproteine con GAGs legati (polimeri ripetuti di disaccaridi con gruppi acidi e solfati legati) Sono frammisti alle fibrille collagene. Invece di fornire forza strutturale, conferiscono proprietà addizionali. L’aggiunta dei GAGs conferisce ai proteoglicani una elevata carica negativa che li porta ad assumere una conformazione distesa in grado di sequestrare sia acqua che cationi divalenti come il Calcio. Queste proprietà conferiscono funzioni di riempitori degli spazi vuoti e di lubrificazione. I GAGs e soprattutto l’eparan solfato, legano inoltre molti fattori secreti e di crescita nella MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 Sono frammisti alle fibrille collagene. Invece di fornire forza strutturale, conferiscono proprietà addizionali. L’aggiunta dei GAGs conferisce ai proteoglicani una elevata carica negativa che li porta ad assumere una conformazione distesa in grado di sequestrare sia acqua che cationi divalenti come il Calcio. Queste proprietà conferiscono funzioni di riempitori degli spazi vuoti e di lubrificazione. I GAGs e soprattutto l’eparan solfato, legano inoltre molti fattori secreti e di crescita nella MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
40
Proteoglicani (Introd. 2)
Nel genoma dei mammiferi ci sono circa 3 dozzine di proteoglicani della MEC che subentrano in diverse famiglie. Le due famiglie più ampie sono quelle basate su ripetizioni LRR («Leucine-Rich Repeat») e quelle contenenti domini LINK (regioni di legame all’acido ialuronico) e domini «C-type lectin» (moduli leganti carboidrati Ca-dipendenti) (ialectani). Molti dei proteoglicani LRR si legano a diversi collageni e a fattori di crescita. I membri della famiglia degli ialectani si legano a varie glicoproteine della MEC (ad es. tenascine) e, mediante il dominio LINK, all’acido ialuronico. Queste funzioni di legame contribuiscono alla regolazione dei complessi proteici nella MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a Leucine-rich repeat
 e quelle contenenti domini LINK (regioni di legame all’acido ialuronico) e domini «C-type lectin» (moduli leganti carboidrati Ca-dipendenti) (ialectani). Molti dei proteoglicani LRR si legano a diversi collageni e a fattori di crescita. I membri della famiglia degli ialectani si legano a varie glicoproteine della MEC (ad es. tenascine) e, mediante il dominio LINK, all’acido ialuronico. Queste funzioni di legame contribuiscono alla regolazione dei complessi proteici nella MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a Leucine-rich repeat.")
41
Proteoglicani (Introd. 3)
Inoltre, ci sono circa una dozzina di Pgs che non cadono in queste famiglie (es. lubricin/PRG4, endocan/ESM1, serglicina, ecc). Forse il più significativo di questi sia il Perlecano (HSPG2), una proteina multidominio che è il «core proteoglycan) di tutte le membrana basali. Ci sono inoltre molti altri esempi di Pgs che cadono in altre categorie (es. alcuni collageni, agrina, betaglicano, CD44 e altre glicoproteine) che sono talvolta o sempre modificate dal legame con GAGs. Ci sono infine due piccole famiglie di PGs integrali di membrana con catene laterali di eparan solfato (HS) Glipicani Sindecani …e alcuni PGs transmembrana a Condroitin Solfato (CS) Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
. Forse il più significativo di questi sia il Perlecano (HSPG2), una proteina multidominio che è il «core proteoglycan) di tutte le membrana basali. Ci sono inoltre molti altri esempi di Pgs che cadono in altre categorie (es. alcuni collageni, agrina, betaglicano, CD44 e altre glicoproteine) che sono talvolta o sempre modificate dal legame con GAGs. Ci sono infine due piccole famiglie di PGs integrali di membrana con catene laterali di eparan solfato (HS) Glipicani. Sindecani. …e alcuni PGs transmembrana a Condroitin Solfato (CS) Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
42
Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
:a")
43
ECM - Glicoproteine Laminina Fibronectina

44
FIBRONECTINA

46
Glicoproteine (Introd. 1)
Circa 200 tipi di glicoproteine complesse che svolgono miriadi di funzioni sono state identificate nel matrisoma dei mammiferi : Interazioni che permettono l’assemblaggio della MEC; Presenza di domini e motivi che promuovono l’adesione cellulare e segnalamento verso le cellule Domini che legano fattori di crescita. I fattori di crescita legati possono servire da serbatoi che possono venire rilasciati (ad es. mediante proteolisi) oppure presentati come ligandi in fase solida dalle proteine della MEC. Le più note sono le laminine (11 geni; 5 tipi di catene α, 3 tipi di catene β, 3 tipi di catena γ) e le fibronectine (1 gene che codifica diverse varianti di splicing). Sono molto studiate anche le trombospondine e le tenascine. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 oppure presentati come ligandi in fase solida dalle proteine della MEC. Le più note sono le laminine (11 geni; 5 tipi di catene α, 3 tipi di catene β, 3 tipi di catena γ) e le fibronectine (1 gene che codifica diverse varianti di splicing). Sono molto studiate anche le trombospondine e le tenascine. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
47
Glicoproteine (Introd. 2)
Due sottogruppi sono studiati particolarmente nell’ambito del sistema nervoso (es. netrins, slits, reelin, agrin, SCO-spondin) e del sistema emostatico (fattore di von Willebrand, vitronectina, fibrinogeno – una proteina facoltativa della MEC). Un altro gruppo molto studiato in diversi contesti normali e patologici che riguardano la regolazione del Transforming Growth Factor beta (TGF- β) comprende le fibrilline e le «Latent transforming growth-factor β-binding proteins”, LTBPs. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 e del sistema emostatico (fattore di von Willebrand, vitronectina, fibrinogeno – una proteina facoltativa della MEC). Un altro gruppo molto studiato in diversi contesti normali e patologici che riguardano la regolazione del Transforming Growth Factor beta (TGF- β) comprende le fibrilline e le «Latent transforming growth-factor β-binding proteins , LTBPs. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
48
Esempi di strutture caratteristiche delle glicoproteine della MEC
Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
:a")
49
Fattori di crescita e secreti legati alla MEC (1)
Molti fattori di crescita si legano alle proteine della MEC e debbono essere considerati come componenti della MEC. E’ vero che molti fattori di crescita e secreti si legano ai GAGs, e specialmente ai GAGs ad eparan solfato. Tuttavia, molti fattori di crescita si possono legare a domini specifici della proteine della matrice: La fbronectina si può legare specificamente a diversi tipi di fattori di crescita (VEGF, HGF, PDGF, ecc.) I domini von Willebrand factor C (VWC/chordin) e follistatin che si trovano in molte proteine legano le “bone morphogenetic proteins» (BMPs), Il TGF si lega specificamente a domini TB delle «Latent transforming growth-factor β- binding proteins” (LTBPs) che a loro volta si legano alle fibrilline e a matrici ricche di fibronectina. Importante: mutazioni nelle fibrilline influenzano la regolazione della funzione del TGF-β nel sindrome di marfan e altre patologie Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 I domini von Willebrand factor C (VWC/chordin) e follistatin che si trovano in molte proteine legano le bone morphogenetic proteins» (BMPs), Il TGF si lega specificamente a domini TB delle «Latent transforming growth-factor β- binding proteins (LTBPs) che a loro volta si legano alle fibrilline e a matrici ricche di fibronectina. Importante: mutazioni nelle fibrilline influenzano la regolazione della funzione del TGF-β nel sindrome di marfan e altre patologie. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
50
Fattori di crescita e secreti legati alla MEC (2)
La MEC può fungere da serbatoio di fattori di crescita. Vi sono molti esempi di questo per le chemochine e per molti dei più importanti fattori di differenziamento (es. VEGFs, Wnt, Hhs, BMPs, FGFs) Tali fattori formano gradienti che controllano la formazione dei diversi pattern tissutali durante lo sviluppo embrionale; I gradienti sono fortemente influenzati dal legame alla MEC. [Chemochine: piccole citochine (molecole di segnalamento) secrete dalle cellule. Il nome deriva dalla loro capacità di indurre la chemotassi in cellule vicine responsive.] Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 Tali fattori formano gradienti che controllano la formazione dei diversi pattern tissutali durante lo sviluppo embrionale; I gradienti sono fortemente influenzati dal legame alla MEC. [Chemochine: piccole citochine (molecole di segnalamento) secrete dalle cellule. Il nome deriva dalla loro capacità di indurre la chemotassi in cellule vicine responsive.] Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
51
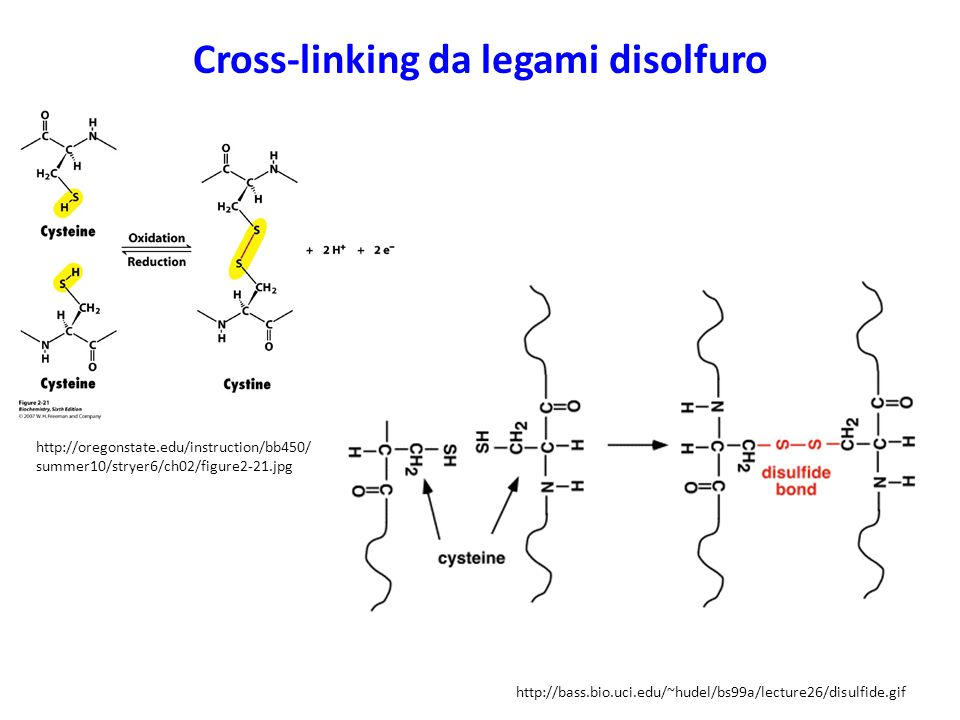
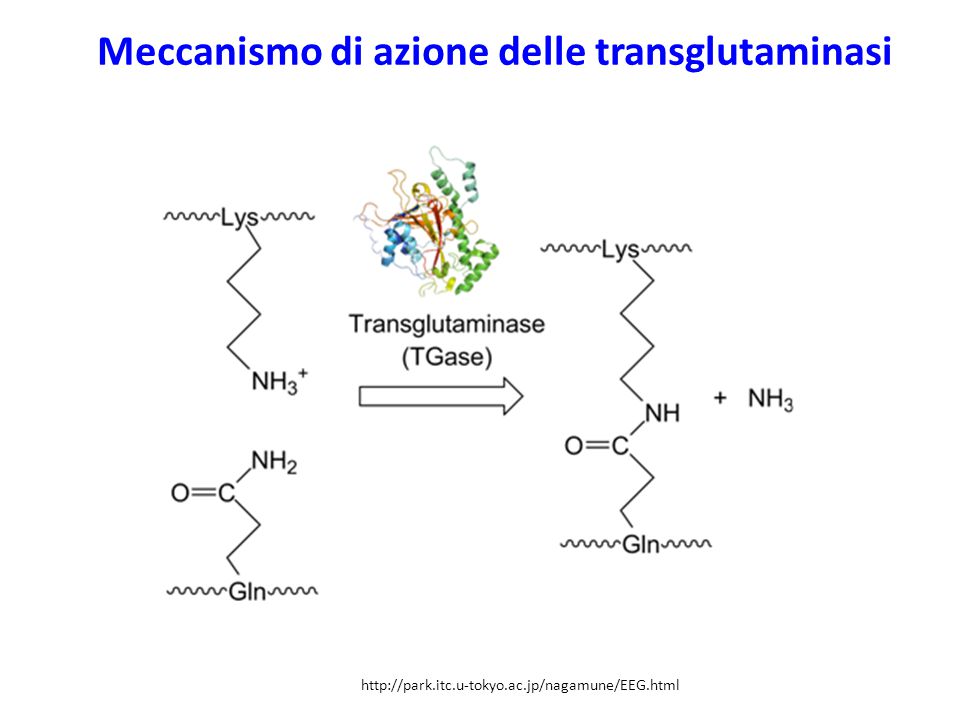
Agenti che modificano la struttura e la funzione della MEC (1)
Le proteine della MEC e le fibrille in cui si assemblano sono in seguito spesso modificate in modo significativo. I collageni subiscono «cross-links» mediante legami disolfuro (S-S) o derivati dall’azione di transglutaminasi o lisil ossidasi e idrossilasi.: Le laminine e altre proteine della membrana basale anche esse subiscono «cross-links» mediante legami disolfuro. Idem per la fibronectina che inoltre subisce ulteriore processamento ad uno stato caratterizzato da insolubilità in desossicolato (DOC). Anche la fibronectina e altre proteine della MEC sono substrati per la transglutaminasi 2 che contribuisce indubbiamente per l’insolubilità della MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 o derivati dall’azione di transglutaminasi o lisil ossidasi e idrossilasi.: Le laminine e altre proteine della membrana basale anche esse subiscono «cross-links» mediante legami disolfuro. Idem per la fibronectina che inoltre subisce ulteriore processamento ad uno stato caratterizzato da insolubilità in desossicolato (DOC). Anche la fibronectina e altre proteine della MEC sono substrati per la transglutaminasi 2 che contribuisce indubbiamente per l’insolubilità della MEC. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
52
Cross-linking da legami disolfuro
53
Meccanismo di azione delle transglutaminasi
54
Agenti che modificano la struttura e la funzione della MEC (2)
Anche gli enzimi proteolitici modificano la MEC: Ad es le pro-peptidasi dei procollageni sono necessarie per processare i collageni in modo che essi possano polimerizzare. I collageni e altre proteine della MEC sono inoltre substrati per le metalloproteinasi della matrice (MMPs) e per le proteasi ADAMTs («A Desintegrin and a Metalloproteinase with a Thrombospondin domain»). Molti altri enzimi proteolitici (elastasi, catepsine, diverse serina esterasi proteasi, ecc.) possono intervenire su diverse proteine della MEC. Queste proteasi sono coinvolte nel turn-over della MEC e inoltre probabilmente contribuiscono a rilasciare fattori di crescita legati alla matrice oppure ad esporre attività criptiche della MEC, incluso il rilascio di inibitori dell’angiogenesi. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a
 e per le proteasi ADAMTs («A Desintegrin and a Metalloproteinase with a Thrombospondin domain»). Molti altri enzimi proteolitici (elastasi, catepsine, diverse serina esterasi proteasi, ecc.) possono intervenire su diverse proteine della MEC. Queste proteasi sono coinvolte nel turn-over della MEC e inoltre probabilmente contribuiscono a rilasciare fattori di crescita legati alla matrice oppure ad esporre attività criptiche della MEC, incluso il rilascio di inibitori dell’angiogenesi. Hynes RO, Naba A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb Perspect Biol Jan 1;4(1):a")
55
Struttura a domini delle MMPs

56
Agenti che modificano la struttura e la funzione della MEC (3)
Allo stesso modo, enzimi che degradano i GAGs, quali ad esempio le ialuronidasi, le eparanasi e le solfatasi, possono alterare le proprietà dei PGs della matrice. Il rimodellamento della MEC da parte di queste attività ha effetti estremamente importanti nello sviluppo embrionale e nelle patologie.

57
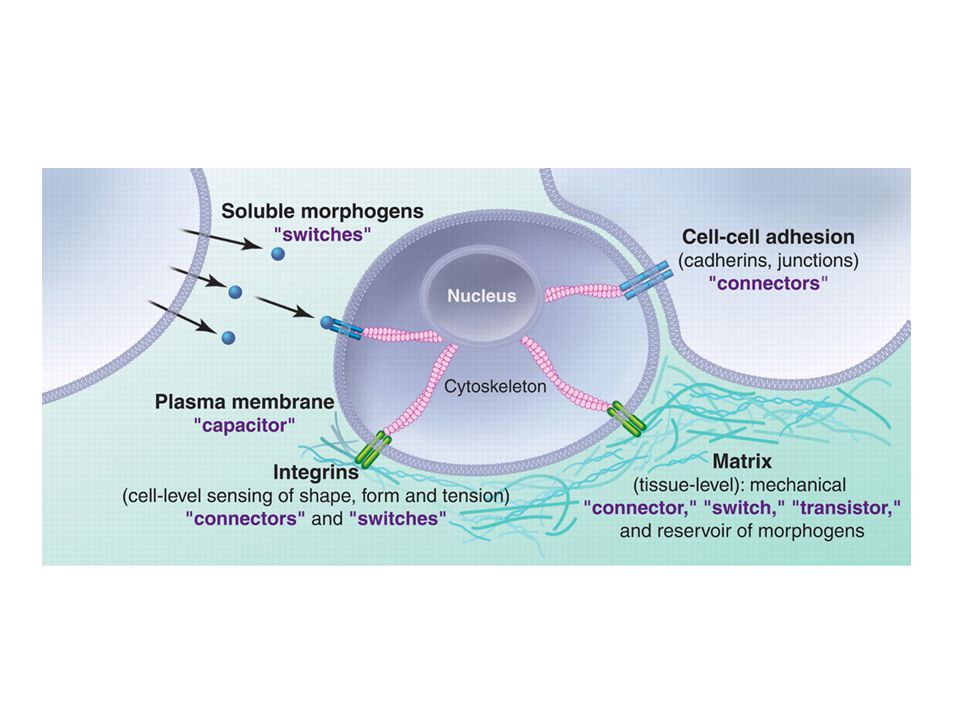
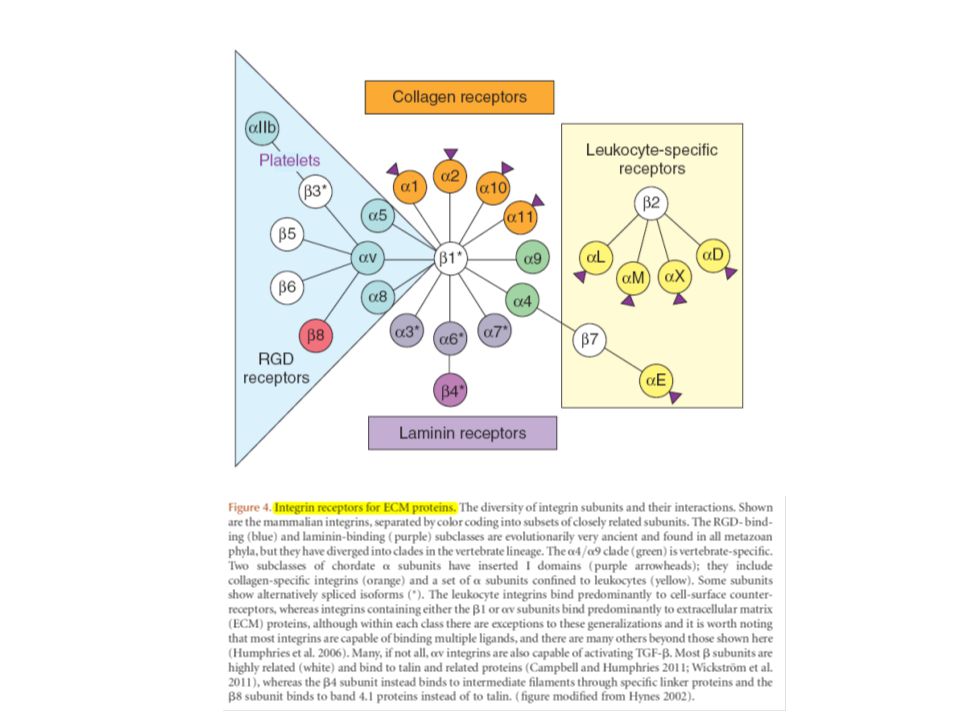
Recettori cellulari per la MEC (integrine e non solo) (1)
La MEC influenza il comportamento cellulare tramite recettori specifici. I principali recettori sono la famiglia delle integrine, che comprende 24 eterodimeri αβ. Un altro recettore per le proteine della MEC è il distroglicano, che si lega alla laminina, agrina e perlecano nelle membrane basali ed anche alle neurexine transmembrana. Ciascuno di questi ligandi del distroglicano contengono domini LamG che si legano al distroglicano in modo dipendente dalla glicosilazione, probabilmente mediante legame alle catene di carboidrati laterali del distroglicano. Mutazioni nel distroglicano o nelle sue proteine associate nella membrana o nel citoscheletro (o nella laminina) possono produrre varie forme di distrofia muscolare, a causa della perdita del collegamento transmembrana alla membrana basale che circonda le cellule muscolari.
 possono produrre varie forme di distrofia muscolare, a causa della perdita del collegamento transmembrana alla membrana basale che circonda le cellule muscolari.")
59
Recettori cellulari per la MEC (integrine e non solo) (2)
Altri recettori cellulari per la MEC includono: La GPVI sulle piastrine e le tirosina chinasi «Discoidin domain receptor» (DDR), tutti recettori per i collageni. Il complesso GPIb/V/IX, che forma un recettore per il fattore di von Willebrand nelle piastrine CD44, che si lega all’ialuronato ed è espresso in molte cellule. Oltre a collegarsi ai ligandi extracellulari questi recettori per la matrice forniscono collegamenti transmembrana al citoscheletro e alle vie di trasduzione del segnale. I domini citoplasmatici dei recettori per la MEC assemblano grandi complessi dinamici di proteine che regolano l’assemblaggio del citoscheletro ed all’interno delle cellule e attivano molte cascate di segnalazione Questo è vero non solo per le integrine ma anche per gli altri tipi di recettore.
, tutti recettori per i collageni. Il complesso GPIb/V/IX, che forma un recettore per il fattore di von Willebrand nelle piastrine. CD44, che si lega all’ialuronato ed è espresso in molte cellule. Oltre a collegarsi ai ligandi extracellulari questi recettori per la matrice forniscono collegamenti transmembrana al citoscheletro e alle vie di trasduzione del segnale. I domini citoplasmatici dei recettori per la MEC assemblano grandi complessi dinamici di proteine che regolano l’assemblaggio del citoscheletro ed all’interno delle cellule e attivano molte cascate di segnalazione. Questo è vero non solo per le integrine ma anche per gli altri tipi di recettore.")
60
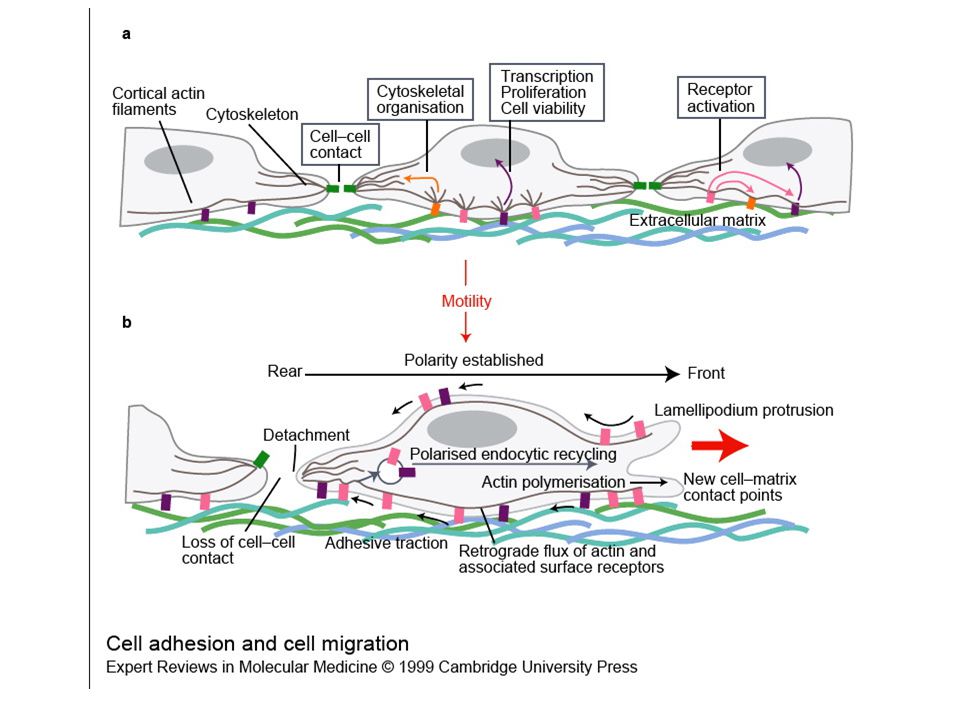
Le componenti extracellulari dell’adesione cellulare
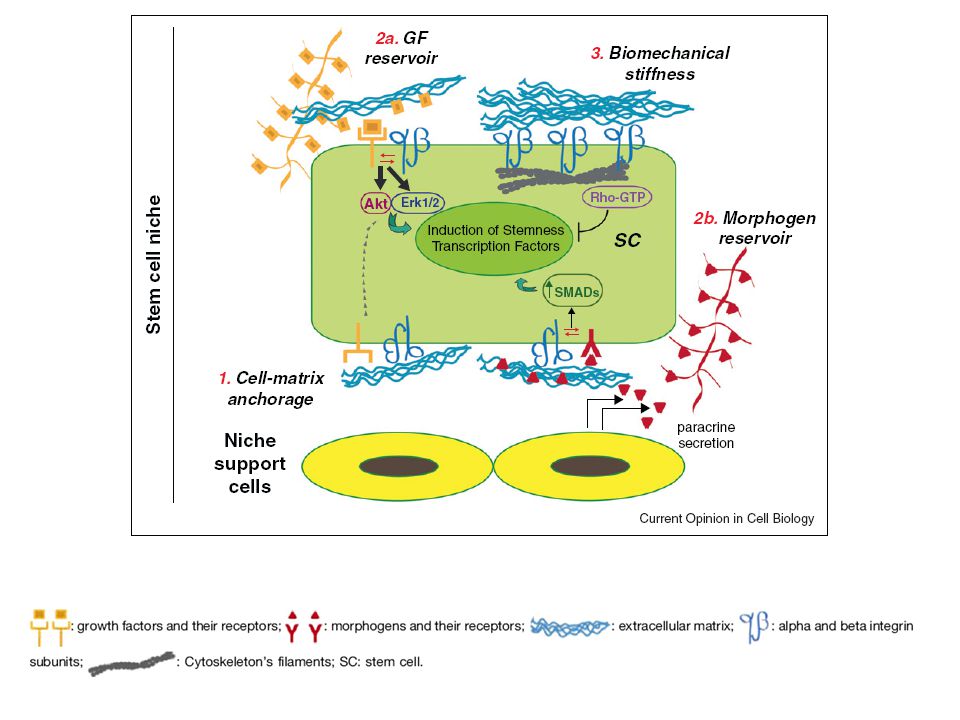
In vitro, l’adesione, l’espansione e la migrazione delle cellule richiedono interazioni tra la cellula e la MEC. Il grado di espansione dipende dalla concentrazione delle componenti della MEC, ossia del numero di contatti focali stabiliti. Quando una cellula incontra quantità limitate di sostanze adesive, essa tende a migrare verso regioni dove la concentrazione è sufficiente per permettere alle cellule di espandersi. Sembra che la migrazione abbia luogo quando le interazioni cellula-matrice sono poche in numero ma tali da conferire la trazione necessaria per la migrazione: L’aumento del numero di punti di adesione cellulare immobilizza la cellula. Si ritiene che meccanismi simili abbiano luogo durante lo sviluppo embrionale, i processi di riparo e la disseminazione metastatica. Smith & Wood, 1999

Presentazioni simili


















>")
 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")

, specializzato per la distribuzione di: gas.>")
>")