Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il cancro come “malattia metabolica”
produrre energia con risorse limitate e favorire percorsi metabolici che possano sostenere la proliferazione cellulare Effetto Warburg IPOSSIA TUMORALE: caratteristica metabolica principale delle cellule tumorali Attivazione del fattore trascrizionale HIF-1 effetto Warburg”, è considerato alla base della progressione tumorale. Inizialmente lo stesso Warburg aveva collegato tale alterazione ad un danno irreversibile a carico dei mitocondri, ipotesi sostenuta da una ridotta espressione di una subunità dell’enzima ATP-sintasi e dalla diminuita attività del complesso del citocromo c ossidasi.La proliferazione cellulare causa ipossia e le cellule si adattano a questa condizione attivando il fattore trascrizionale

2
Effetto Warburg Switch metabolico si riduce l’uso del ciclo degli acidi tricarbossilici ed aumenta l’attività della glicolisi anaerobica. Elevato consumo di glucosio e produzione di acido lattico Warburg osservò in cellule tumorali elevato consumo di glucosio e produzione di acido lattico .L’osservazione non ebbe seguito per molti anni, anche perché la scoperta di oncogeni e oncosoppressori ha deviato in altra direzione l’interesse scientifico. SCIENCE VOL MAY 2009

3
Le cellule cancerose sono metabolicamente più attive e captano più glucosio delle cellule normali
Tomografia a emissione di positroni (PET) somministrazione per via endovenosa al paziente di molecole marcate con radioisotopi che emettono positroni; tra queste la più utilizzata è il fluoro-desossi-glucosio (18F-FDG). Eccesso di FDG è escreto con urine SCIENCE VOL 324, 22 MAY 2009
 somministrazione per via endovenosa al paziente di molecole marcate con radioisotopi che emettono positroni; tra queste la più utilizzata è il fluoro-desossi-glucosio (18F-FDG). Eccesso di FDG è escreto con urine. SCIENCE VOL 324, 22 MAY")
4
Cell Metabolism 7, January 2008
In una cellula che non è in attiva proliferazione, l’acido piruvico entra nel ciclo TCA e la produzione di ATP è legata alla fosforilazione ossidativa. In una cellula in attiva proliferazione, invece, si ha aumento di glicolisi e produzione di lattato che serve a rigenerare il nad* per la glicolisi. Warburg aveva collegato tale alterazione ad un danno irreversibile a carico dei mitocondri Cell Metabolism 7, January 2008

5
Fermentazione lattica
Glicolisi

6
Produzione di molecole necessarie alla crescita
Perché avviene lo switch metabolico? -Utilizzo del nutriente più abbondante per produrre ATP -Possibilità di utilizzare intermedi della glicolisi a scopi biosintetici -Possibilità di utilizzare intermedi del Ciclo degli acidi tricarbossilici a scopi biosintetici Produzione di molecole necessarie alla crescita e divisione cellulare

7
Metabolismo glucosio La velocità della glicolisi è aumentata di circa 10 volte in cellule tumorali. Glucosio fornisce carbonio per i processi anabolici, energia, NADPH e ribosio-5-fosfato (via del pentoso fosfato) Attivazione lattico deidrogenasi Inibizione piruvato chinasi SCIENCE VOL 324, 22 MAY 2009
 Attivazione lattico deidrogenasi. Inibizione piruvato chinasi. SCIENCE VOL 324, 22 MAY")
8
Fermentazione lattica

9
Trasferimento dell’acetil CoA dal mitocondrio al citosol
Citrato liasi Citrato sintasi Malato deidrogenasi Enzima malico

10
Negli epatociti e negli adipociti il NADPH citosolico
ENZIMA MALICO Negli epatociti e negli adipociti il NADPH citosolico è generato dall’enzima malico Determina la sintesi di piruvato che rientra nei mitocondri

12
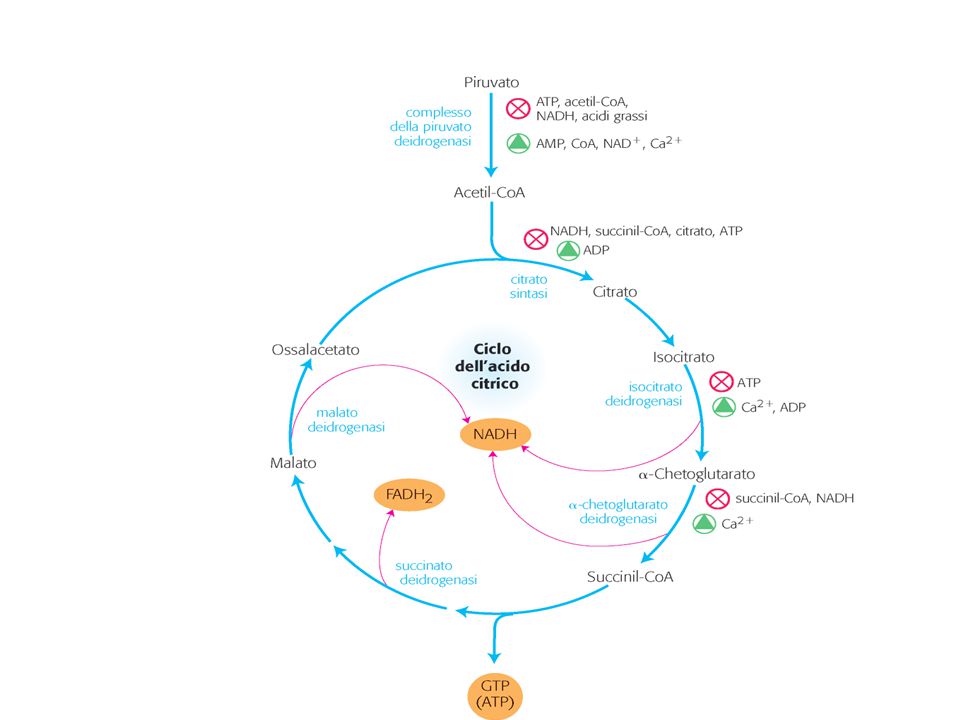
Funzioni biosintetiche del ciclo degli acidi tricarbossilici

13
Metabolismo glutammina
Glutammina fornisce N per sintesi purine e pirimidine, intermedi del ciclo degli acidi tricarbossilici (TCA cycle) Funzione anaplerotica Cell Metabolism 7, January 2008
 Funzione anaplerotica. Cell Metabolism 7, January")
14
TRANSAMINAZIONE DEAMINAZIONE OSSIDATIVA ac.glutammico

15
e risposte al microambiente tumorale
Il fenotipo metabolico di una cellula tumorale deriva da mutazioni genetiche e risposte al microambiente tumorale Oncogeni e oncosoppressori possono influenzare lo switch tra glicolisi anaerobica e ciclo degli acidi tricarbossilici (modulando l’espressione di specifici enzimi) e aumentare la captazione di glucosio e di glutammina. Il destino metabolico del glucosio nelle cellule tumorali contribuisce al fenotipo tumorale Vie di trasduzione alterate modificano il metabolismo cellulare The metabolic phenotype of tumour cells is controlled by intrinsic genetic mutations and external responses to the tumour microenvironment. Oncogenic signalling pathways controlling growth and survival are often activated by the loss of tumour suppressors (such as p53) or the activation of oncoproteins (such as PI3K). The resulting altered signalling modifies cellular metabolism to match the requirements of cell division. Abnormal microenvironmental conditions such as hypoxia, low pH and/or nutrient deprivation elicit responses from tumour cells, including autophagy, which further affect metabolic activity. These adaptations optimize tumour cell metabolism for proliferation by providing appropriate levels of energy in the form of ATP, biosynthetic capacity and the maintenance of balanced redox status. Nature Reviews Cancer vol.11, February 2011
 e aumentare la captazione di glucosio e di glutammina. Il destino metabolico del glucosio nelle cellule tumorali contribuisce al fenotipo tumorale. Vie di trasduzione alterate modificano il metabolismo cellulare. The metabolic phenotype of tumour cells is controlled by intrinsic genetic mutations and external responses to the tumour microenvironment. Oncogenic signalling pathways controlling growth and survival are often activated by the loss of tumour suppressors (such as p53) or the activation of oncoproteins (such as PI3K). The resulting altered signalling modifies cellular metabolism to match the requirements of cell division. Abnormal microenvironmental conditions such as hypoxia, low pH and/or nutrient deprivation elicit responses from tumour cells, including autophagy, which further affect metabolic activity. These adaptations optimize tumour cell metabolism for proliferation by providing appropriate levels of energy in the form of ATP, biosynthetic capacity and the maintenance of balanced redox status. Nature Reviews Cancer vol.11, February")
16
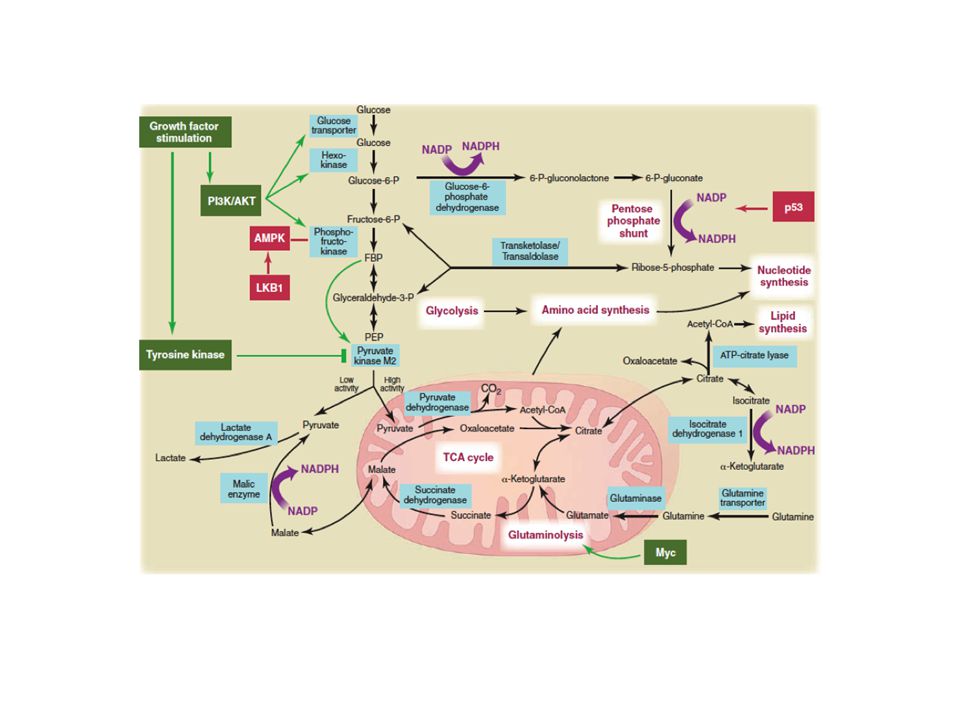
Signaling Network to Regulate Metabolism in Proliferating Cells
Constitutive activation of the PI3K/Akt/mTOR pathway can occur in tumors due to mutation of tumor suppressors, or by other mechanisms. Metabolic effects include: enhanced uptake of glucose and essential amino acids and protein translation. Translation of HIF-1a is enhanced. The transcription factor Myc increases expression of many metabolic enzymes, including glycolytic enzymes, LDH-A, and several enzymes required for nucleotide biosynthesis. The model shows some of the prominent aspects of metabolism in proliferating cells, including glycolysis; lactate production; the use of TCA cycle intermediates as macromolecular precursors; and the biosynthesis of proteins, nucleotides, and lipids. The PI3K/Akt/mTOR pathway, HIF-1a, and Myc participate in various facets of this metabolic phenotype. The binding of a growth factor (GF) to its surface receptor brings about activation of PI3K and the serine/threonine kinases Akt and mTOR. Cell Metabolism 7, January 2008
 to its surface receptor brings about activation of PI3K and the serine/threonine kinases Akt and mTOR. Cell Metabolism 7, January")
18
Effetti della Piruvato chinasi su glicolisi e via del pentoso fosfato
Pyruvate kinase isoform M2 (PKM2) is present in very few types of proliferating normal cells but is present at high levels in cancer cells. PKM2 catalyses the rate-limiting step of glycolysis, controlling the conversion of phosphoenolpyruvate (PEP) to pyruvate, and thus ATP generation. By slowing the passage of metabolites through glycolysis, PKM2 promotes the shuttling of these substrates through the pentose phosphate pathway (PPP) and other alternative pathways so that large quantities of reduced nicotinamide adenine dinucleotide phosphate (NADPH) and other macromolecules are produced. These molecules are required for macromolecule biosynthesis and the maintenance of redox balance that is needed to support the rapid cell division that occurs within a tumour. Nature Reviews Cancer vol.11, February 2011
 is present in very few types of proliferating normal cells but is present at high levels in cancer cells. PKM2 catalyses the rate-limiting step of glycolysis, controlling the conversion of phosphoenolpyruvate (PEP) to pyruvate, and thus ATP generation. By slowing the passage of metabolites through glycolysis, PKM2 promotes the shuttling of these substrates through the pentose phosphate pathway (PPP) and other alternative pathways so that large quantities of reduced nicotinamide adenine dinucleotide phosphate (NADPH) and other macromolecules are produced. These molecules. are required for macromolecule biosynthesis and the maintenance of redox balance that is needed to support the rapid cell division that occurs within a tumour. Nature Reviews Cancer vol.11, February")
20
PKM2 Phosphorylates Histone H3 and Promotes Gene Transcription and Tumorigenesis
Cell 150, 685–696, August 17, 2012 Tumor-specific pyruvate kinase M2 (PKM2) is essential for the Warburg effect. In addition to its well established role in aerobic glycolysis, PKM2 directly regulates gene transcription. In seguito ad attivazione di Epidermal growth factor receptor, che è presente in molti tumori umani, si osserva la traslocazione di PMK2 nel nucleo PKM2 Regulates Gene Expression by H3-T11 Phosphorylation. EGFR activation results in nuclear translocation of PKM2 that binds to gene promoter regions, where PKM2 phosphorylates H3-T11, leading to HDAC3 disassociation from the promoters and subsequent acetylation of histone H3, transcription of genes (c-Myc, cyclin D1), cell-cycle progression, and cell proliferation.
 is essential for the Warburg effect. In addition to its well established. role in aerobic glycolysis, PKM2 directly. regulates gene transcription. In seguito ad attivazione di Epidermal growth factor receptor, che è presente in molti tumori umani, si osserva la traslocazione di PMK2 nel nucleo. PKM2 Regulates Gene Expression by H3-T11 Phosphorylation. EGFR activation results in nuclear translocation of PKM2 that binds to gene. promoter regions, where PKM2 phosphorylates H3-T11, leading to HDAC3. disassociation from the promoters and subsequent acetylation of histone H3, transcription of genes (c-Myc, cyclin D1), cell-cycle progression, and cell proliferation.")
21
Controllo del potenziale redox in cellule tumorali
The production of two of the most abundant antioxidants, NADPH and GS), has been shown to be modulated in cancers. Pyruvate kinase isoform M2 (PKM2), which is overexpressed in many cancer cells, can divert metabolic precursors away from glycolysis and into the pentose phosphate pathway (PPP) to produce NADPH. NADP-dependent isocitrate dehydrogenase 1 (IDH1, IDH2) and malic enzyme 1(ME1) also contribute to NADPH production. MYC increases glutamine uptake and glutaminolysis, driving the de novo synthesis of GSH. Additionally, MYC contributes to NADPH production by promoting the expression of PKM2. Together, NADPH and GSH control increased levels of reactive oxygen species (ROS) driven by increased cancer cell proliferation. Nature Reviews Cancer vol.11, February 2011
, has been shown to be modulated in cancers. Pyruvate kinase isoform M2 (PKM2), which is overexpressed in. many cancer cells, can divert metabolic precursors away from glycolysis and into the pentose phosphate pathway (PPP) to produce NADPH. NADP-dependent isocitrate dehydrogenase 1 (IDH1, IDH2) and malic enzyme 1(ME1) also contribute to NADPH production. MYC increases glutamine uptake and glutaminolysis, driving the de novo synthesis of GSH. Additionally, MYC contributes to NADPH production by promoting the expression of PKM2. Together, NADPH and GSH control increased levels of reactive oxygen species (ROS) driven by increased cancer cell proliferation. Nature Reviews Cancer vol.11, February")
22
Riduzione di alfa-chetoglutarato a 2-idrossiglutarato Oncometabolita?
Mutazioni di IDH1 sono state riscontrate in gliomi e leucemie mieloidi acute. Riduzione di alfa-chetoglutarato a 2-idrossiglutarato Oncometabolita? Nature Reviews Cancer vol.11, February 2011

23
L’ipossia è una condizione diffusa nei tumori
- Molti tumori umani hanno valori di pO2 bassi - Una ipossia severa non è normalmente presente in tessuti normali, ma esiste spesso nei tumori - Il tumore prolifera più rapidamente di cellule normali - La capacità di adattarsi all’ipossia è essenziale per la crescita tumorale

24
Complesso eterodimerico HIF-1
In ipossia le cellule attivano un tipico programma di espressione genica regolato principalmente dal fattore di trascrizione indotto da ipossia HIF-1 Superfamiglia dei fattori trascrizionali eucariotici con dominio basic-helix-loop-helix (bHLH). Complesso eterodimerico HIF-1 subunità α (120 kDa) regolata dalla concentrazione di ossigeno subunità β (91/94 kDa) costitutivamente espressa
. Complesso eterodimerico HIF-1. subunità α (120 kDa) regolata dalla concentrazione di ossigeno. subunità β (91/94 kDa) costitutivamente espressa.")
25
Struttura di HIF-1 prolinnnnnnnnnnnnn
Il dominio bHLH e il dominio PAS (Per/ARNT/Sim) sono coinvolti nel legame con il DNA e nella dimerizzazione. Il dominio ODD (dominio di degradazione ossigeno dipendente) contiene due residui di prolina regolatori riconosciuti ed idrossilati in normossia da prolil-idrossilasi (PHD). I domini TAD (domini di transattivazione) interagiscono con i coattivatori trascrizionali (CBP/p300) e contengono un residuo di asparagina che viene idrossilato in normossia
 sono coinvolti nel legame con il DNA e nella dimerizzazione. Il dominio ODD (dominio di degradazione ossigeno dipendente) contiene due residui di prolina regolatori riconosciuti ed idrossilati in normossia da prolil-idrossilasi (PHD). I domini TAD (domini di transattivazione) interagiscono con i coattivatori trascrizionali (CBP/p300) e contengono un residuo di asparagina che viene idrossilato in normossia.")
26
PHDs sono degli enzimi prolina idrossilasi la cui attività dipende dalla presenza di O2, Fe (2+). PHD2 idrossila HIF-1α a livello di due residui di prolina (dominio ODD) promuovendone il reclutamento da parte di pVHL pVHL modulo di riconoscimento di una E3-ubiquitina ligasi responsabile della ubiquitinazione di HIF-1α e successiva degradazione per via proteosomica. The researchers, therefore, created a new version of VHL that does not stop working when oxygen is scarce. Introducing this newly engineered version of VHL into mice that had kidney tumours dramatically reduced levels of HIF, caused tumours to regress and limited the formation of new blood vessels within the tumours. Oxygen-independent degradation of HIF1-a via bioengineered VHLtumour suppressor complex (EMBO Mol.Med. 2009)
")
27
IPOSSIA o in mancanza di
NORMOSSIA la subunità a viene continuamente degradata perchè una prolil-idrossilasi (PH) attivata da O2 e Fe idrossila uno specifico residuo di prolina. Tale residuo viene riconosciuto dalla proteina di Von Hippel Lindau (VHL) che media l’aggiunta di ubiquitina ad opera di ubiquitin-ligasi (UL) e la degradazione della subunità α nel proteasoma. IPOSSIA o in mancanza di VHL ciò non avviene; entrambe le subunità di HIF sono presenti e i geni dell’ipossia vengono trascritti.
 attivata da O2 e Fe. idrossila uno specifico residuo. di prolina. Tale residuo viene. riconosciuto dalla proteina. di Von Hippel Lindau (VHL) che media l’aggiunta di. ubiquitina ad opera di. ubiquitin-ligasi (UL) e la degradazione della subunità α. nel proteasoma. IPOSSIA o in mancanza di. VHL ciò non avviene; entrambe le subunità di HIF sono presenti e i geni dell’ipossia vengono trascritti.")
28
Traslocazione nucleare
Stabilizzazione Traslocazione nucleare Formazione eterodimero Attivazione trascrizionale FIH-1 è una asparagina idrossilasi sensibile a O2, 2-oxoglutarato e Fe che blocca il legame con CBP/p300 (un cofattore che permette l’attivazione della trascrizione da parte di HIF) Fattore inibente HIF (FIH) è una asparagina idrossilasi, dipendente da O2 a-chetiglutarato e Fe: L’idrossilazione blocca l’interazione con CBP/p300 Prolil-idrossilasi (PHD): Km per O mM Fattore inibente HIF (FIH): Km per O2 90mM
 Fattore inibente HIF (FIH) è una asparagina idrossilasi, dipendente da O2. a-chetiglutarato e Fe: L’idrossilazione blocca l’interazione con CBP/p300. Prolil-idrossilasi (PHD): Km per O2 230 mM. Fattore inibente HIF (FIH): Km per O2 90mM.")
29
In condizioni di ipossia HIF-1a non viene degradato ma trasloca nel nucleo e dimerizza con HIF1-b. L’eterodimero interagisce con CBP/p300, riconosce e si lega alla sequenza nota come elemento regolatore dell’ipossia (HRE) presente sul promotore di geni indotti dall’ipossia, attivandone la loro trascrizione.
 presente sul promotore di geni indotti dall’ipossia, attivandone la loro trascrizione..")
30
Ruolo di HIF nel metabolismo
Trasportatori del glucosio GLUT1, GLUT3 Enzimi della glicolisi Piruvato deidrogenasi chinasi 1 (PDK1) Trasportatori di acidi monocarbossilici e di H+ Anidrasi Carbonica 9
 Trasportatori di acidi monocarbossilici e di H+ Anidrasi Carbonica 9.")
31
HIF-1 Attiva la piruvato deidrogenasi chinasi 1 (PDK1)
PDK1 fosforila la piruvato deidrogenasi inattivandola Ciò impedisce la conversione del piruvato in Acetil CoA e l’utilizzazione del piruvato da parte dei mitocondri. Contributo di HIF all’effetto Warburg Myc, che è attivato nel 40% dei tumori umani, coopera con HIF-1 nell’attivazione trascrizionale di PDK1

32
Metabolismo del Glucosio
Processi biologici Geni Angiogenesi Inibitore dell’attivatore del Plasminogeno tissutale (PAI-1), fattore di crescita dell’endotelio vascolare (VEGF), recettore FLT- 1 del VEGF Proliferazione cellulare e sopravvivenza CMET: recettore tirosina chinasico in grado di promuovere la proliferazione, la migrazione, ed altri meccanismi cellulari. ciclina G2, endotelina 1, eritropoietina, eme- ossigenasi 1 , fattore di crescita insulin-simile 2 (IGF-2), proteine leganti l’IGF (IGFBP -1,-2,-3), ossido nitricosintasi 2, p21, Microambiente extracellulare Anidrasi Carbonica 9, collagenedi tipo V (ccl), prolil-4-idrossilasi Metabolismo del Glucosio Trasportatori del glucosio GLUT1 , GLUT3 Enzimi glicolitici aldolasi A, aldolasi C, gliceraldeide-3-fosfato deidrogenasi, enolasi, esochinasi, lattato deidrogenasi, fosfofruttochinasi fosfogliceratochinasi 1 , piruvato chinasi, triosofosfato isomerasi Omeostasi del ferro Ceruloplasmina, transferrina, recettore della transferrina Altri fattori di trascrizione DEC1 , p35srg, recettore per gli attivatori dei perossisomialfa (PPARalfa) Tabella 1‑1: Geni target di HIF-1(Semenza 2002)
, fattore di crescita dell’endotelio vascolare (VEGF), recettore FLT- 1 del VEGF. Proliferazione cellulare e sopravvivenza. CMET: recettore tirosina chinasico in grado di promuovere la proliferazione, la migrazione, ed altri meccanismi cellulari. ciclina G2, endotelina 1, eritropoietina, eme- ossigenasi 1 , fattore di crescita insulin-simile 2 (IGF-2), proteine leganti l’IGF (IGFBP -1,-2,-3), ossido nitricosintasi 2, p21, Microambiente. extracellulare. Anidrasi Carbonica 9, collagenedi tipo V (ccl), prolil-4-idrossilasi. Metabolismo del Glucosio. Trasportatori del glucosio GLUT1 , GLUT3. Enzimi glicolitici aldolasi A, aldolasi C, gliceraldeide-3-fosfato deidrogenasi, enolasi, esochinasi, lattato deidrogenasi, fosfofruttochinasi fosfogliceratochinasi 1 , piruvato chinasi, triosofosfato isomerasi. Omeostasi del ferro. Ceruloplasmina, transferrina, recettore della transferrina. Altri fattori di trascrizione. DEC1 , p35srg, recettore per gli attivatori dei perossisomialfa (PPARalfa) Tabella 1‑1: Geni target di HIF-1(Semenza 2002)")
Presentazioni simili