Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Sequenza-struttura-funzione
diverse combinazioni dei 20 aa G P Y W I V R T A C D Varietà di strutture e funzioni F S Q L E K N M H ASSOCIARE A CIASCUNA PROTEINA DI CUI CONOSCIAMO LA SEQUENZA UNA O PIU’ SPECIFICHE FUNZIONI A LIVELLO MOLECOLARE

2
Codice strutturale ESPERIMENTO DI ANFINSEN
NELLA SEQUENZA PRIMARIA E’ SCRITTA LA STRUTTURA DI UNA PROTEINA PROTEINE CON SEQ. PRIMARIE SIMILI TENDONO AD AVERE STRUTTURE 3D SIMILI CONFRONTO TRA GENOMI → i geni essenziali (ciclo cell, sviluppo embrionale, signalling) soggetti a poca variabilità
 soggetti a poca variabilità.")
3
Omologia, Identità, Similarità
Similarità –termine generico che indica una somiglianza che può essere dovuta a omologia, ma anche alla convergenza evolutiva. E’ un parametro quantitativo (posso quantificarla in %). Omologia – implica un’origine evolutiva comune e spesso anche similarità. E’ un parametro qualitativo. Prima di Darwin l’omologia era definita morfologicamente Con la genetica si sa che è dovuta a geni (antenati) comuni Due sequenze sono omologhe se sono simili e hanno un antenato in comune Se due sequenze sono omologhe la loro struttura 3D sarà conservata
. Omologia – implica un’origine evolutiva comune e spesso anche similarità. E’ un parametro qualitativo. Prima di Darwin l’omologia era definita morfologicamente. Con la genetica si sa che è dovuta a geni (antenati) comuni. Due sequenze sono omologhe se sono simili e hanno un antenato in comune. Se due sequenze sono omologhe la loro struttura 3D sarà conservata.")
4
Similarità dovuta ad evoluzione dallo stesso antenato comune
L’esistenza di similarità in brevi tratti di sequenze può essere casuale Similarità sufficientemente estesa implicano un’omologia (anche se non abbiamo una prova diretta di un comune antenato) Di conseguenza è possibile misurare il grado di significatività della similarità OMOLOGIA OMOPLASIA Similarità dovuta ad evoluzione dallo stesso antenato comune Similarità dovuta a convergenza evolutiva – derivano da linee diverse in seguito ad es. a una pressione selettiva simile
 Di conseguenza è possibile misurare il grado di significatività della similarità. OMOLOGIA OMOPLASIA. Similarità dovuta ad evoluzione dallo stesso antenato comune. Similarità dovuta a convergenza evolutiva – derivano da linee diverse in seguito ad es. a una pressione selettiva simile.")
5
Ortologhi/Paraloghi Mediante confronti di geni simili tra genomi diversi, e di geni simili dello stesso genoma, si può stabilire se due geni sono ortologhi o paraloghi, e da qui ricostruire la probabile storia evolutiva Proteine o geni omologhi possono appartenere alla stessa specie o a specie diverse e possono avere origine in seguito a eventi di Speciazione/Duplicazione

6
Ortologhi/Paraloghi Si può ipotizzare che i vari geni “simili” tra loro che troviamo nelle diverse specie, lo siano perché “parenti”, ovvero discendenti dallo stesso/i gene/i in specie antenate (speciazione) o nella stessa specie (duplicazione) Due sequenze (sia DNA, sia RNA, sia proteine) per cui possiamo fare questa ipotesi – basandoci sulla loro similarità – sono dette sequenze omologhe
 o nella stessa specie (duplicazione) Due sequenze (sia DNA, sia RNA, sia proteine) per cui possiamo fare questa ipotesi – basandoci sulla loro similarità – sono dette sequenze omologhe.")
7
Che cosa diversifica nel tempo due sequenze omologhe?
I 3 meccanismi più comuni attraverso i quali le sequenze mutano nel corso dell’evoluzione sono: Sostituzione Delezione Inserzione Inserzione e delezione sono una l’opposto dell’altra e vengono indicate con il termine indels

8
Sukkar Europa, circa 700 dC Sugar Suiker Sucre Zucker Zuckre Sokker
Zucchero Açucar Azucar Sakari Sukkar Europa, circa 700 dC

9
L’evoluzione delle parole
Tutte le parole delle lingue moderne che indicano lo “zucchero” discendono da una parola antenata comune Tutte dalla stessa (“sukkar” - parola usata dagli arabi), alcune da un’antenata più vicina nel tempo (“zuckre” in Francia)
, alcune da un’antenata più vicina nel tempo ( zuckre in Francia)")
10
L’evoluzione molecolare
Ipotesi: tutte le specie esistenti discendono da specie “antenate” Nel corso dell’evoluzione, da una specie possono nascere una o più specie diverse (speciazione) Allora, dovremmo essere in grado di trovare “tracce” dell’evoluzione... nel DNA! Se una specie discende da un’altra, allora il suo DNA è simile a quello della specie “antenata”, con - ovviamente - delle variazioni Più la speciazione è vicina nel tempo, più il DNA (in sequenza) è “simile”
 Allora, dovremmo essere in grado di trovare tracce dell’evoluzione... nel DNA! Se una specie discende da un’altra, allora il suo DNA è simile a quello della specie antenata , con - ovviamente - delle variazioni. Più la speciazione è vicina nel tempo, più il DNA (in sequenza) è simile")
11
L’albero della vita

12
Il - “vero” - albero della vita

13
L’evoluzione delle parole
Immaginiamo di non conoscere le parole “antenate” dello zucchero, e di doverci chiedere se due parole moderne in due lingue differenti sono “simili” tra loro SUGAR SUGR SUCR SUCRE

14
Allineamento S U G A R - S U C - R E
L’”allineamento” è un modo di rappresentare schematicamente i legami evolutivi tra due o più parole (o sequenze), indicando sostituzioni, inserzioni e delezioni S U G A R - S U C - R E Sostituzione (mutazione) Inserzioni (delezioni)
, indicando sostituzioni, inserzioni e delezioni. S U G A R - S U C - R E. Sostituzione. (mutazione) Inserzioni. (delezioni)")
15
Allineamento (multiplo)
S U G - A R - S U C – - R E Z U C K E R - S O K K E R - A Z U C - A R - S A K - A R I A ç U C - A R - - S U C(K)A R -
A R -")
16
SSH_UOMO -MLLLARCLLLVLVSSLLVCSGLACGPGRGFGKRRHPKKLTPLAYKQFIPNVAEKTLGAS
SSH_TOPO MLLLLARCFLVILASSLLVCPGLACGPGRGFGKRRHPKKLTPLAYKQFIPNVAEKTLGAS :******:*::*.******.*************************************** SSH_UOMO GRYEGKISRNSERFKELTPNYNPDIIFKDEENTGADRLMTQRCKDKLNALAISVMNQWPG SSH_TOPO GRYEGKITRNSERFKELTPNYNPDIIFKDEENTGADRLMTQRCKDKLNALAISVMNQWPG *******:**************************************************** SSH_UOMO VKLRVTEGWDEDGHHSEESLHYEGRAVDITTSDRDRSKYGMLARLAVEAGFDWVYYESKA SSH_TOPO VKLRVTEGWDEDGHHSEESLHYEGRAVDITTSDRDRSKYGMLARLAVEAGFDWVYYESKA ************************************************************ SSH_UOMO HIHCSVKAENSVAAKSGGCFPGSATVHLEQGGTKLVKDLSPGDRVLAADDQGRLLYSDFL SSH_TOPO HIHCSVKAENSVAAKSGGCFPGSATVHLEQGGTKLVKDLRPGDRVLAADDQGRLLYSDFL *************************************** ******************** SSH_UOMO TFLDRDDGAKKVFYVIETREPRERLLLTAAHLLFVAPHNDSATGEPEASSGSGPPSGGAL SSH_TOPO TFLDRDEGAKKVFYVIETLEPRERLLLTAAHLLFVAPHND SGPTPG--- ******:*********** ********************* ***..* SSH_UOMO GPRALFASRVRPGQRVYVVAERDGDRRLLPAAVHSVTLSEEAAGAYAPLTAQGTILINRV SSH_TOPO PSALFASRVRPGQRVYVVAERGGDRRLLPAAVHSVTLREEEAGAYAPLTAHGTILINRV * *******************.*************** ** *********:******** SSH_UOMO LASCYAVIEEHSWAHRAFAPFRLAHALLAALAPARTDRGGDSGGGDRGGGGGRVALTAPG SSH_TOPO LASCYAVIEEHSWAHRAFAPFRLAHALLAALAPARTD GGGGGSIP-AAQS ************************************* ***** :. :* . SSH_UOMO AADAPGAGATAGIHWYSQLLYQIGTWLLDSEALHPLGMAVKSS SSH_TOPO ATEARGAEPTAGIHWYSQLLYHIGTWLLDSETMHPLGMAVKSS *::* ** .************:*********::**********

17
ALLINEAMENTO DI SEQUENZE
A COPPIE AGTTTGAATGTTTTGTGTGAAAGGAGTATACCATGAGATGAGATGACCACCAATCATTTC ||||||||||||||||||| |||||||| ||| | |||||| ||||||||||||||||| AGTTTGAATGTTTTGTGTGTGAGGAGTATTCCAAGGGATGAGTTGACCACCAATCATTTC MULTIPLO KFKHHLKEHLRIHSGEKPFECPNCKKRFSHSGSYSSHMSSKKCISLILVNGRNRALLKTl KYKHHLKEHLRIHSGEKPYECPNCKKRFSHSGSYSSHISSKKCIGLISVNGRMRNNIKT- KFKHHLKEHVRIHSGEKPFGCDNCGKRFSHSGSFSSHMTSKKCISMGLKLNNNRALLKRl KFKHHLKEHIRIHSGEKPFECQQCHKRFSHSGSYSSHMSSKKCV KYKHHLKEHLRIHSGEKPYECPNCKKRFSHSGSYSSHISSKKCISLIPVNGRPRTGLKTs

18
Allineamento GLOBALE o LOCALE
GLOBALE considera la similarita’ tra due sequenze in tutta la loro lunghezza (da N- a C-terminale) LOCALE considera solo specifiche REGIONI simili tra alcune parti delle sequenze in analisi (solo regioni a ↑ densità di similarità generando più sub-allineamenti) Global alignment LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||. | | | .| .| || || | || TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHKAG Local alignment LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||||||||.|||| TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHK
 LOCALE considera solo specifiche REGIONI simili tra alcune parti delle sequenze in analisi (solo regioni a ↑ densità di similarità generando più sub-allineamenti) Global alignment. LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||. | | | .| .| || || | || TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHKAG. Local alignment. LTGARDWEDIPLWTDWDIEQESDFKTRAFGTANCHK ||||||||.|||| TGIPLWTDWDLEQESDNSCNTDHYTREWGTMNAHK.")
19
ALLINEAMENTI DI SEQUENZA
Per confrontare delle sequenze queste devono essere allineate ALLINEAMENTO: procedura per confrontare 2 o più sequenze residuo per residuo in modo da massimizzare la similarità tra esse e ridurre il numero di operazioni da effettuare per convertirle l’una nell’altra. E’ volto a stabilire una relazione biunivoca tra le coppie di residui delle sequenze considerate. L’allineamento di sequenze è strumento indispensabile per: CONFRONTO tra due sequenze; RICERCA DI SEQ SIMILI a una in esame NELLE BANCHE DATI; Determinazione di PATTERN e DOMINI CONSERVATI; PREDIZIONE DI STRUTTURA 3D; Stimare L’APPARTENENZA a UN CERTO FOLD; COSTRUIRE UN ALBERO FILOGENETICO: PREDIZIONE DI STRUTTURA SECONDARIA

20
ALLINEAMENTO DI SEQUENZA
PER ESEGUIRE UN ALLINEAMENTO DI SEQUENZA SONO NECESSARI ESSENZIALMENTE 3 STRUMENTI: Avere a disposizione una MATRICE DI SOSTITUZIONE. La matrice definisce la il GRADO di SIMILARITA’ tra amminoacidi; Avere a disposizione un ALGORITMO DI ALLINEAMENTO cercando di massimizzare il punteggio dato dalla matrice e valutando quanti gap (interruzioni) inserire; Avere a disposizione per evitare allineamenti senza senso una PENALITA’ per l’introduzione dei GAP. I GAP riflettono inserzioni/delezioni avvenute durante l’evoluzione LLTTVRNN LLTTVRNN LLVRNN LL--VRNN
 inserire; Avere a disposizione per evitare allineamenti senza senso una PENALITA’ per l’introduzione dei GAP. I GAP riflettono inserzioni/delezioni avvenute durante l’evoluzione. LLTTVRNN LLTTVRNN. LLVRNN LL--VRNN.")
21
Similarità e distanza Esistono due modi per misurare il grado di omologia tra due sequenze: Calcolare la similarità contando i match Calcolare la distanza contando mismatch e indels Similarità elevata ↔ bassa distanza Due sequenze identiche hanno una distanza pari a zero

22
SIMILARITA’ DI SEQUENZA
Nel punteggio di similarità di sequenza si tiene conto del fatto che gli amminoacidi a confronto in ogni posizione siano simili, differenti o identici e di una penalità per i gap. PER DEFINIRE LA SIMILARITA’ TRA LE DUE SEQUENZE SI USANO MATRICI BASATE SU PRESUPPOSTI DIVERSI: identità/non identità; Caratteristiche chimico-fisiche degli aa; Basate sul codice genetico: valutare quante mutazioni fare in una tripletta per passare da un aa a un altro. (se ad es. si cambia un solo nucleotide la sostituzione la sostituzione sarà meno penalizzata perché si tratta di evento probabile nel corso dell’evoluzione) Basate su criteri evolutivi estrapolati da confronto di sequenze di proteine omologhe (MATRICI BLOSUM E PAM) 2 penalità per i gap (apertura (fisso), estensione (lunghezza dipendente))
 Basate su criteri evolutivi estrapolati da confronto di sequenze di proteine omologhe (MATRICI BLOSUM E PAM) 2 penalità per i gap (apertura (fisso), estensione (lunghezza dipendente))")
23
Penalizzazioni degli indels
Generalmente si usano funzioni del tipo “lineare” wx = gx o, più frequentemente, di tipo “affine” wx = g + rx oppure wx = g + r(x—1) dove g è il punteggio di penalizzazione per l’apertura, r per l’elongazione e x la lunghezza dell’indel
 dove g è il punteggio di penalizzazione per l’apertura, r per l’elongazione e x la lunghezza dell’indel.")
24
Matrici di sostituzione
Le matrici di sostituzione tengono conto dei criteri di similarità tra aminoacidi Comprendono 210 valori: 20 (sulla diagonale) relativi al punteggio dell’appaiamento di ciascun aminoacido con se stesso 190 relativi a tutte le possibili sostituzioni aminoacidiche I 190 valori sono riportati anche nella loro parte speculare in modo che queste matrici hanno un formato 20 x 20 valori Le matrici di sostituzione più semplici considerano solo il criterio di identità e sono costituite da valori 0 o 1 Altre matrici considerano la similarità chimica tra gli aminoacidi o il numero minimo di mutazioni per passare da un codon all’altro e attribuiscono un punteggio alle diverse sostituzioni
 relativi al punteggio dell’appaiamento di ciascun aminoacido con se stesso. 190 relativi a tutte le possibili sostituzioni aminoacidiche. I 190 valori sono riportati anche nella loro parte speculare in modo che queste matrici hanno un formato 20 x 20 valori. Le matrici di sostituzione più semplici considerano solo il criterio di identità e sono costituite da valori 0 o 1. Altre matrici considerano la similarità chimica tra gli aminoacidi o il numero minimo di mutazioni per passare da un codon all’altro e attribuiscono un punteggio alle diverse sostituzioni.")
25
Similarità chimico-fisica
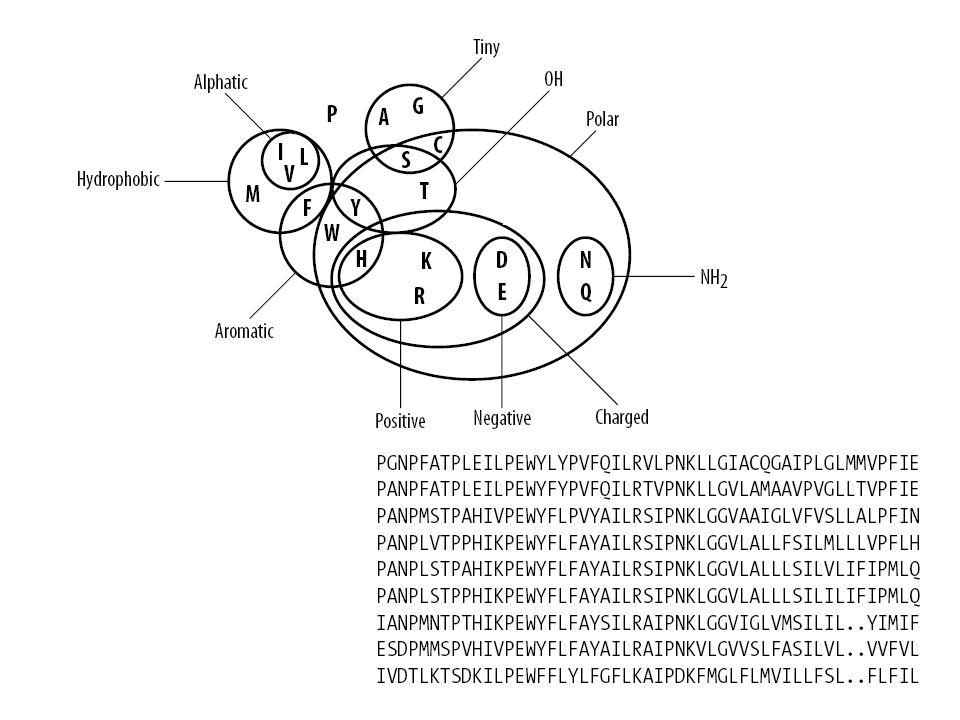
Gli aminoacidi possono essere raggruppati in base alle caratteristiche fisico-chimiche delle loro catene laterali. Su questa base un aminoacido può essere definito simile ad un altro R K basici I V F L idrofobici G A poco ingombro sterico R K H D E carichi R K N Q polari

27
Similarità definita da criteri “genetici”
Quando sostituisco un nucleotide all’interno di una regione codificante, l’effetto dipende dalla sua posizione all’interno del codone CUU (Leu/L)Leucine CUC (Leu/L)Leucine CUA (Leu/L)Leucine CUG (Leu/L)Leucine In questo caso, cambiando il terzo nucleotide, non cambia nulla (mutazioni “silenti”) AUU (Ile/I) Isoleucine AUC (Ile/I) Isoleucine AUA (Ile/I) Isoleucine AUG (Met/M) Methionine, Start In questo caso, cambiando il primo nucleotide, ottengo amminoacidi non troppo differenti dalla leucina... il codice genetico sembra essere “robusto”, ma..
Leucine CUC (Leu/L)Leucine CUA (Leu/L)Leucine CUG (Leu/L)Leucine. In questo caso, cambiando il terzo nucleotide, non cambia nulla (mutazioni silenti ) AUU (Ile/I) Isoleucine AUC (Ile/I) Isoleucine AUA (Ile/I) Isoleucine AUG (Met/M) Methionine, Start. In questo caso, cambiando il primo nucleotide, ottengo amminoacidi non troppo differenti dalla leucina... il codice genetico sembra essere robusto , ma..")
28
Mutazioni deleterie E V
La mutazione di un nucleotide nel sesto codone dell’emoglobina-beta (una delle sequenze che vanno a costituire l’emoglobina), causa la sostituzione dell’amminoacido codificato, problemi nella struttura dell’ emoglobina stessa, e il cambiamento di forma dei globuli rossi, risultando nell’anemia falciforme
, causa la. sostituzione dell’amminoacido codificato, problemi nella struttura dell’ emoglobina stessa, e il cambiamento di forma dei globuli rossi, risultando. nell’anemia falciforme.")
29
Quale matrice PAM conviene utilizzare?
In generale per due sequenze filogeneticamente vicine è meglio utilizzare una matrice PAM a basso indice e viceversa In assenza di informazioni si utilizzano PAM40, PAM120 e PAM 250 PAM250 individua similarità del 20% PAM120 individua similarità del 40% PAM80 individua similarità del 50% PAM60 individua similarità del 60% 27 marzo 2017

30
L’utilizzo della matrice di similarita’ appropriata per ciascuna analisi e’ cruciale per avere buoni risultati. Infatti relazioni importanti da un punto di vista biologico possono essere indicate da anche molto debole similarità. Sequenze poco divergenti molto divergenti BLOSUM80 BLOSUM62 BLOSUM45 PAM1 PAM120 PAM250

31
Confronto tra matrici PAM e BLOSUM
Le matrici PAM sono basate su un modello evolutivo Le matrici BLOSUM sono basate su famiglie di proteine Le matrici PAM sono basate su un allineamento globale Le matrici BLOSUM sono basate su un allineamento locale Le matrici PAM sono utili per studi evolutivi sulle proteine Le matrici BLOSUM sono fatte per individuare le regioni conservate

Presentazioni simili











>")






 Algoritmi di allineamento 2) Algoritmi di ricerca in database>")
