Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Secrezione pancreatica dal pancreas esocrino

2
Il pancreas è una ghiandola a funzione mista che si trova parallelo e dietro allo stomaco con una struttura simile a quella delle ghiandole salivari. Il pancreas esocrino secerne tutti gli enzimi necessari alla digestione di grassi, carboidrati e proteine. Secerne inoltre una componente acquosa ricca in bicarbonato.

6
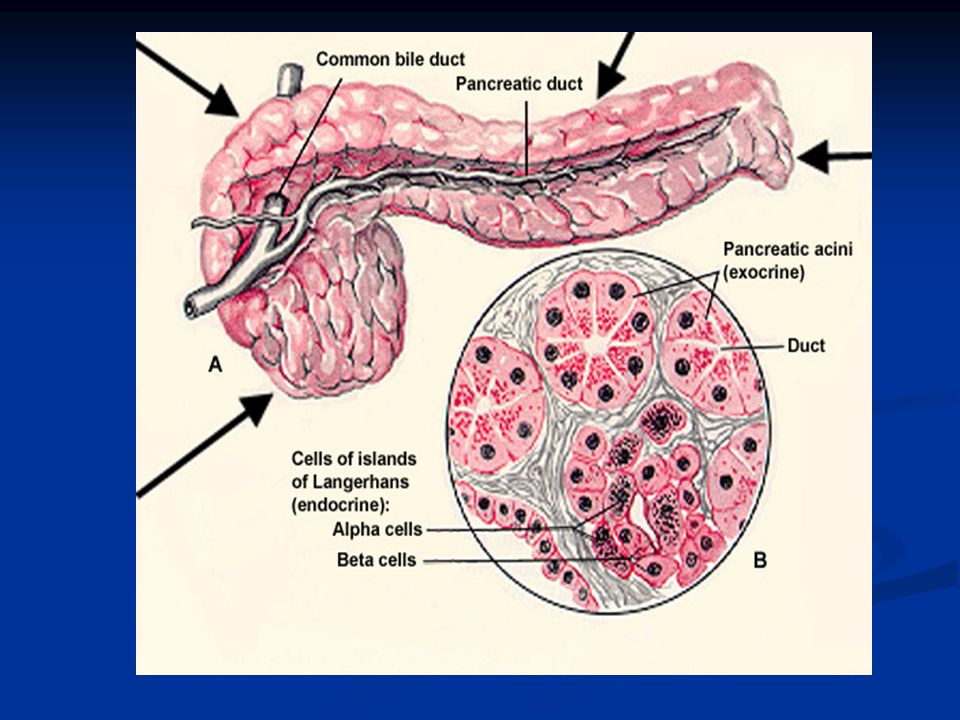
Il pancreas è costituito al 98% da acini che secernono la componente enzimatica con funzione digestiva, mentre il restante 2% è costituito dalle isole del Langerhans che formano la porzione endocrina del pancreas. All’interno degli isolotti si trovano 3 diversi tipi cellulari che secernono: Cellule α →glucagone Cellule β →insulina Cellule δ →somatostatina

7
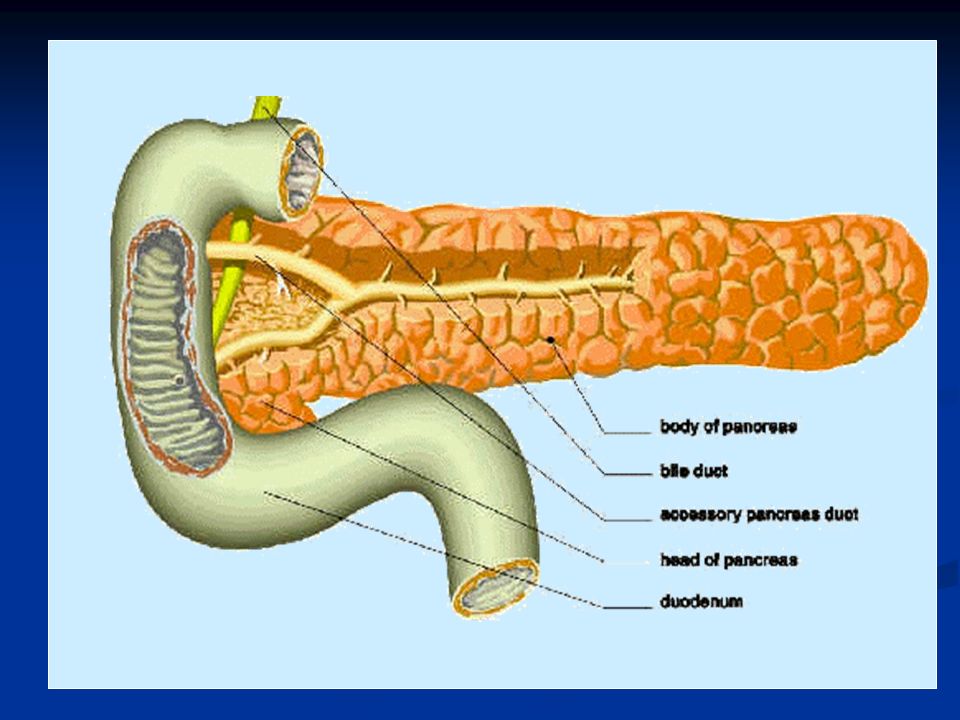
Il prodotto della secrezione esocrina passa attraverso il dotto pancreatico che si unisce al dotto biliare per riversare il contenuto nel duodeno attraverso la papilla di Vater che chiude lo sfintere di Oddi. Lo stimolo per la secrezione esocrina del pancreas è la presenza di cibo nel duodeno e le caratteristiche del succo pancreatico dipendono in buona misura dal tipo di cibo presente nel chimo.

8
Struttura del pancreas esocrino. Il succo pancreatico viene riversato in dotti di calibro via via crescente e infine nel dotto pancreatico che sfocia nel duodeno.

9
Enzimi pancreatici Il pancreas secerne tutti gli enzimi necessari alla digestione di proteine, carboidrati e grassi. Inoltre secerne una componente acquosa che contiene bicarbonato che ha la funzione di neutralizzare il chimo acido proveniente dallo stomaco. Il totale ammonta a circa 1000ml al giorno. I più importanti enzimi proteolitici sono: Tripsina (il più abbondante) ChimotripsinaCarbossipeptidasi Elastasi e nucleasi (meno importanti) Separa alcuni peptidi in singoli aa Non rilasciano singoli aa ma piccoli peptidi
 ChimotripsinaCarbossipeptidasi Elastasi e nucleasi (meno importanti) Separa alcuni peptidi in singoli aa Non rilasciano singoli aa ma piccoli peptidi.")
10
Questi enzimi sono secreti in forma inattiva dal pancreas (tripsinogeno, chimotripsinogeno, procarbossipeptidasi) e sono attivati solo nel tratto intestinale. Un enzima detto enterochinasi attiva la tripsina e questa a sua volta attiva gli altri enzimi proteolitici. Questo è importante perché altrimenti questi enzimi digerirebbero le cellule degli acini: per questo motivo le cellule pancreatiche secernono anche un enzima tripsina inibitore.

11
Fra gli enzimi per la digestione dei grassi ricordiamo: lipasi pancreatiche che idrolizzano i grassi neutri in acidi grassi e monogliceridi colesterolo esterasi che idrolizza gli esteri del colesterolo fosfolipasi che separano gli acidi grassi dai fosfolipidi

12
Fra gli enzimi per la digestione dei carboidrati ricordiamo l’amilasi pancreatica che idrolizza amido, glicogeno e gli altri zuccheri a formare disaccaridi e qualche trisaccaride.

13
La componente acquosa del succo pancreatico è costituita da acqua e bicarbonato ed è secreta dalle cellule epiteliali dei dotti pancreatici. In generale gli stimoli per la secrezione della componente enzimatica e della componente acquosa sono differenti. Ad alti livelli di secrezione la concentrazione di bicarbonato nel plasma si aggira sui 145 mM. Secrezione di bicarbonato

14
La secrezione di Na + insieme a HCO 3 - serve per mantenere l’elettroneutralità nel lume Scambio dei cloruri

15
Ingresso di CO 2 che forma H + e HCO 3 - con AC Ingresso di CO 2 che forma H + e HCO 3 - con AC HCO 3 - è scambiato con Cl - e finisce nel lume (trasporto attivo secondario) HCO 3 - è scambiato con Cl - e finisce nel lume (trasporto attivo secondario) H + è scambiato con Na + ; Na + deve poi passare nel lume. Viene drenata anche dall’acqua per via transcellulare H + è scambiato con Na + ; Na + deve poi passare nel lume. Viene drenata anche dall’acqua per via transcellulare

16
Gli stimoli principali per la secrezione pancreatica sono quelli presentati nella diapositiva seguente. La secrezione pancreatica deriva dall’azione combinata di questi tre stimoli che si potenziano a vicenda come avviene per la secrezione gastrica.

17
ACh: rilasciata dal nervo vago; agisce sulla componente enzimatica Colecistochinina (CCK): secreta dalla mucosa duodenale e digiunale all’ingresso del cibo; agisce sulla componente enzimatica Secretina: secreta sempre da mucosa duodenale e digiunale quando arriva cibo molto acido dallo stomaco; agisce sulla componente acquosa
: secreta dalla mucosa duodenale e digiunale all’ingresso del cibo; agisce sulla componente enzimatica Secretina: secreta sempre da mucosa duodenale e digiunale quando arriva cibo molto acido dallo stomaco; agisce sulla componente acquosa")
18
Il chimo acido induce rilascio di secretina dalle cellule S duodenali Secretina e CCK sono assorbite nel torrente sanguigno La secretina induce il rilascio della componente acquosa del succo pancreatico; CCK induce il rilascio della componente enzimatica ACh induce il rilascio della componente enzimatica negli acini

19
Fase cefalica: gli stessi segnali che stimolano la secrezione gastrica, determinano il rilascio di ACh a livello pancreatico. Questo determina circa il 20% della produzione di enzimi, mentre la componente acquosa è esigua. Il risultato è che la componente enzimatica arriva al duodeno in piccole quantità perché non c’è flusso di liquido.

20
Fase gastrica: secrezione di un ulteriore 5-10% di componente enzimatica sotto stimolazione vagale. Anche qui molto poco arriva al duodeno perché la componente acquosa è ancora scarsa. Fase intestinale: quando il chimo arriva nel duodeno si ha produzione di secretina che stimola fortemente la secrezione della componente acquosa e anche di CCK che stimola ulteriormente la secrezione della componente enzimatica.

21
Cellule S della mucosa duodenale e digiunale duodenale e digiunale Prosecretina Secretina (27aa) Chimo acido che entra nel duodeno con pH < 4 duodeno con pH < 4 Incremento della secrezione della componente acquosa della componente acquosa contenente bicarbonato
 Chimo acido che entra nel duodeno con pH < 4 duodeno con pH < 4 Incremento della secrezione della componente acquosa della componente acquosa contenente bicarbonato")
22
HCl + NaHCO 3 NaCl + H 2 CO 3 CO 2 + H 2 O eliminata a livello polmonare polmonare pH ideale per gli enzimi pancreatici Quando il pH nel duodeno arriva intorno a 3.0 sono secrete grandi quantità di secretina e quindi di ioni bicarbonato che neutralizzano l’acido.

23
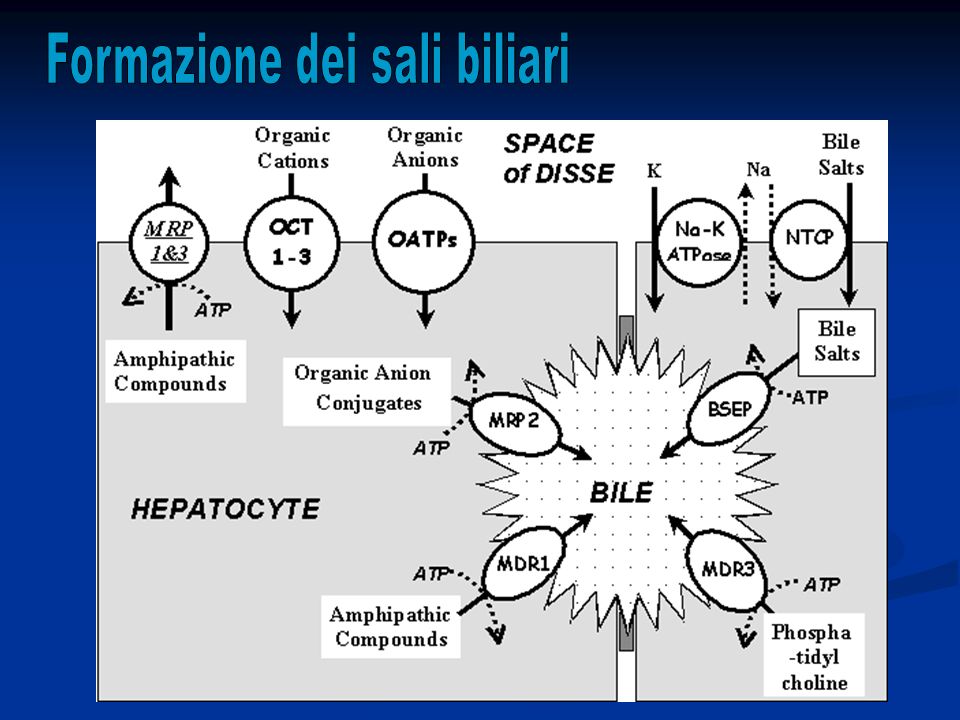
Il fegato ha molteplici funzioni che interessano vari aspetti del metabolismo corporeo. Fra queste funzioni, direttamente correlata alla funzione digestiva è la capacità di secernere bile, normalmente circa 600-1200 ml/die.

24
La bile ha una funzione fondamentale nel metabolismo dei grassi, grazie alla sua composizione in acidi biliari perché: Emulsionano grosse particelle di grasso in piccole particelle che possono essere attaccate dalle lipasi Favoriscono il trasporto e l‘assorbimento dei grassi a livello della mucosa intestinale Inoltre la bile è un sistema di escrezione di prodotti di rifiuto dal sangue, quali bilirubina ed eccessi di colesterolo sintetizzato a livello epatico.

25
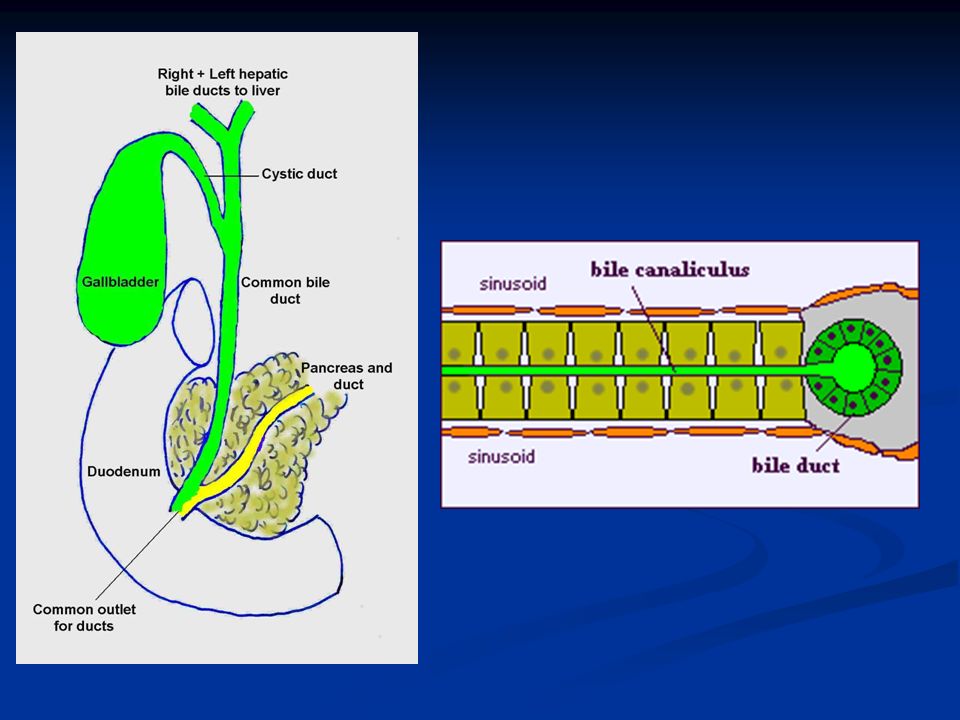
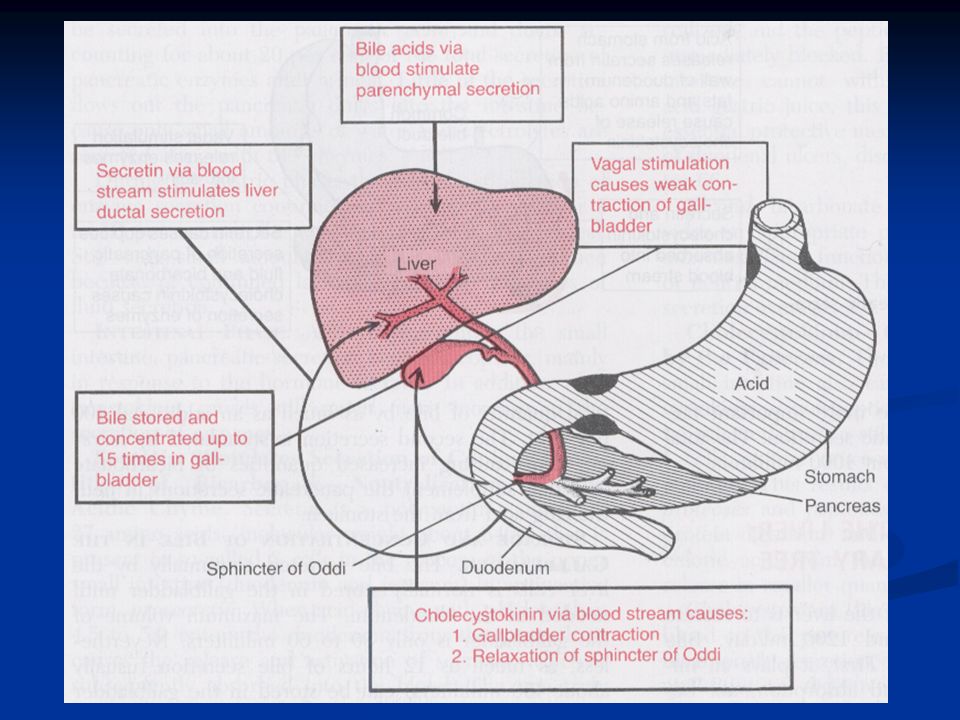
La bile è secreta in due steps: I.La parte iniziale è secreta dagli epatociti: questa contiene grandi quantità di acidi biliari, colesterolo e altri costituenti organici. Viene secreta nei canalicoli biliari. II.La bile poi fluisce perifericamente verso i setti interlobulari dove i canalicoli si gettano nei dotti biliari terminali fino ad arrivare al dotto epatico e al dotto biliare comune da dove la bile passa direttamente nel duodeno o viene riversata, attraverso il dotto cistico, nella cistifellea.

26
Attraversando i dotti biliari le cellule epiteliali dei dotti secernono una soluzione acquosa con Na + e HCO 3 -. Questa attività secretiva è stimolata dalla secretina.

27
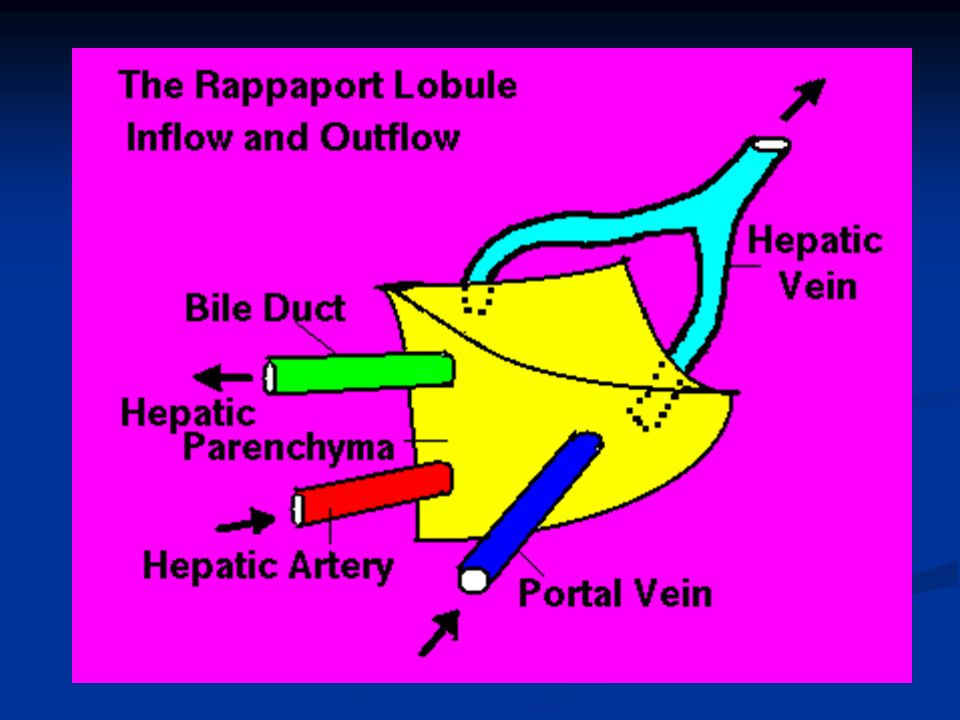
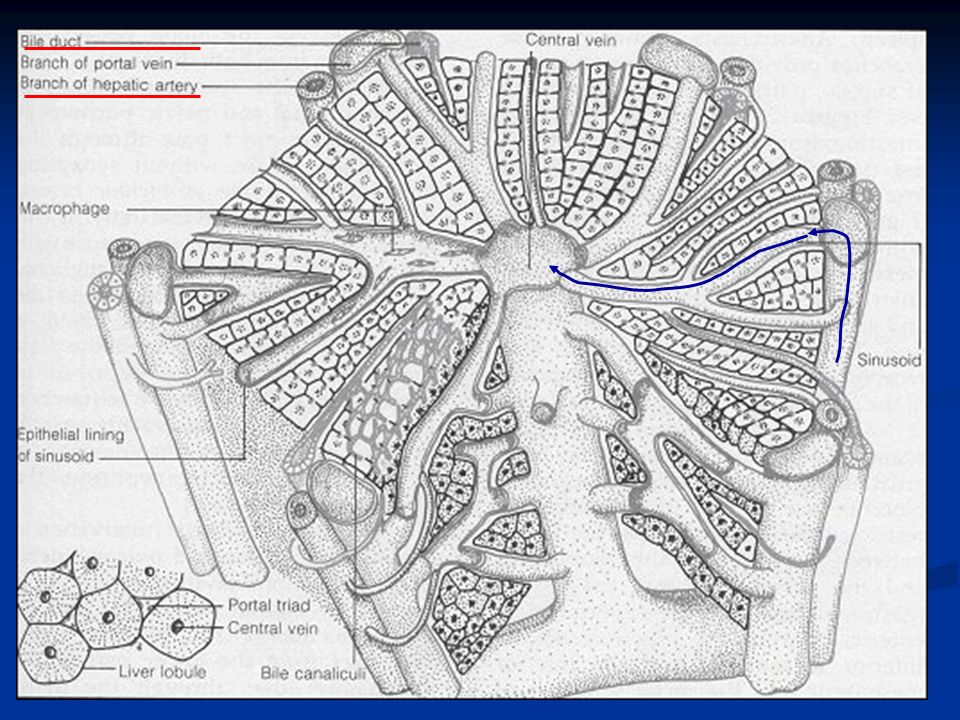
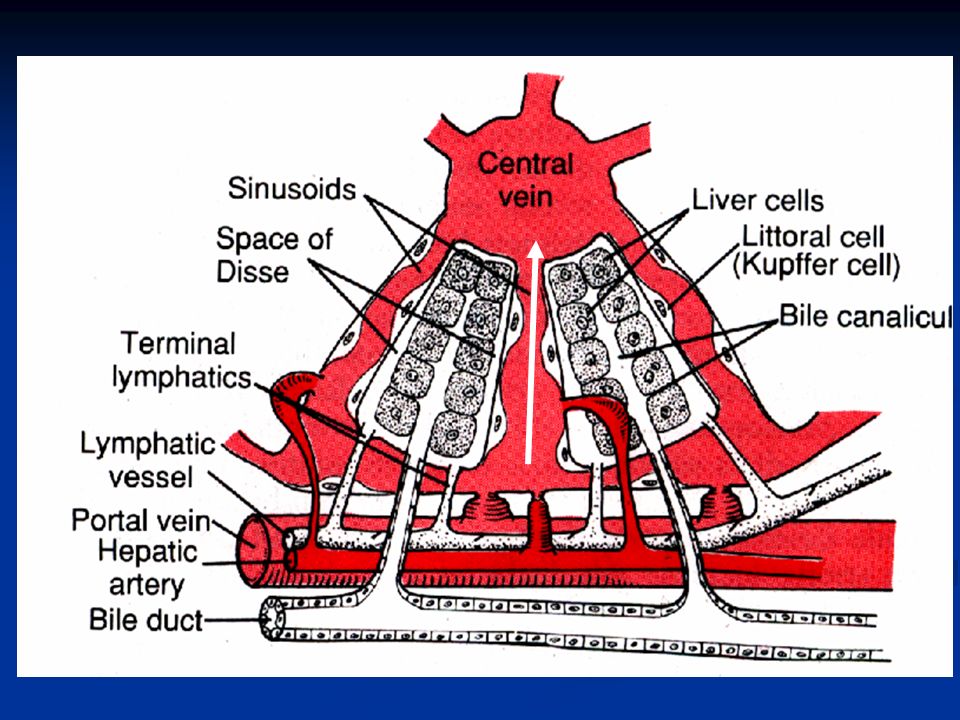
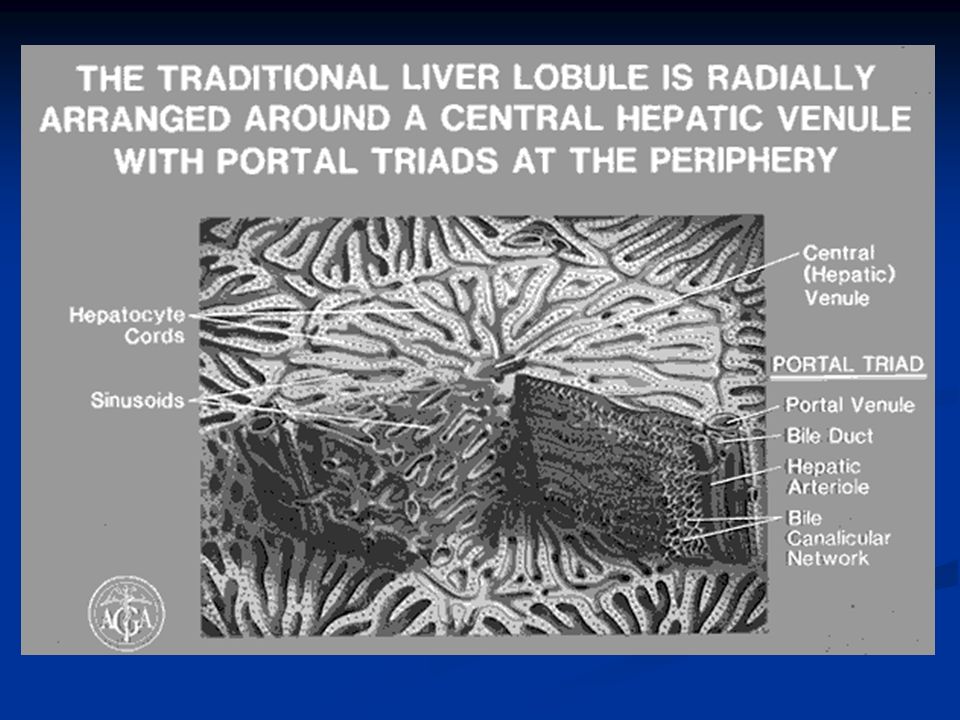
Nel fegato ci sono circa 50.000-100.000 lobuli, ciascuno dei quali è 0.8-2mm di diametro e diversi mm di lunghezza. Ogni lobulo è organizzato intorno ad una vena centrale che si svuota nelle vene epatiche e poi nella cava inferiore. In direzione centrifuga si diramano diverse piastre formate ciascuna da due linee di epatociti e tra le cellule adiacenti decorrono canalicoli biliari che si svuotano nei dotti biliari al margine del lobulo.

28
Al margine dei lobuli si trova anche un’arteriola epatica e una venula portale (triade portale) che porta il sangue refluo da milza, stomaco e intestino. Questo sangue passa nei sinusoidi epatici che si trovano fra una piastra e l’altra e da qui alla vena centrale. Nelle piastre oltre agli epatociti troviamo altri due tipi cellulari: Cellule endoteliali Cellule di Kupffer o cellule reticoloendoteliali: queste sono macrofagi destinate a fagocitare materiale particolato o batteri per impedire che passino in circolo.

31
a= vena centrale b= epatociti c= sinusoidi d= arteriole (da arteria epatica) e= vaso linfatico f= venule dalla vena porta
 e= vaso linfatico f= venule dalla vena porta")
35
La bile secreta dagli epatociti è generalmente immagazzinata nella cistifellea fino al pasto successivo, con una capacità volumetrica di circa 30- 60 ml. Tuttavia è possibile mantenere la secrezione di 12 ore (più o meno 450 ml) grazie al fatto che nella cistifellea acqua ed alcuni elettroliti sono assorbiti dalla mucosa e quindi la bile risulta fortemente concentrata in termini di sali biliari, colesterolo, lecitina e bilirubina. Questo riassorbimento avviene per trasporto attivo di Na + attraverso l’epitelio seguito da assorbimento secondario di Cl - e quindi di acqua.
 grazie al fatto che nella cistifellea acqua ed alcuni elettroliti sono assorbiti dalla mucosa e quindi la bile risulta fortemente concentrata in termini di sali biliari, colesterolo, lecitina e bilirubina. Questo riassorbimento avviene per trasporto attivo di Na + attraverso l’epitelio seguito da assorbimento secondario di Cl - e quindi di acqua..")
36
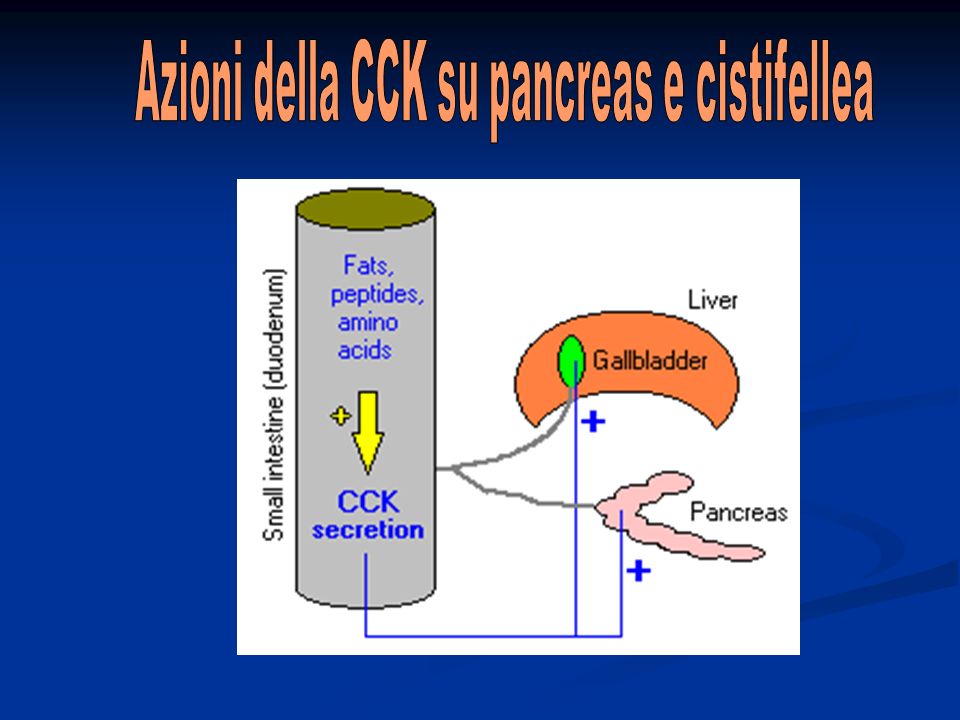
Quando il cibo comincia ad essere digerito nel tratto gi superiore, la cistifellea inizia a svuotarsi per aumento delle contrazioni della parete. Lo stimolo principale per iniziare queste contrazioni è la CCK che viene rilasciata quando sostanze grasse passano nel duodeno.

37
Bile epatica Bile della cistifellea Sali biliari* 35mM310mM Bilirubina* 0.8mM3.2mM Colesterolo* 3mM25mM Acidi grassi 0.12gm/dl0.3-.2gm/dl Lecitina* 1mM10mM Na + 165mM135mM Cl - 100mM25mM

40
Affinché la bile passi nel duodeno insieme al succo pancreatico è necessario che lo sfintere di Oddi della papilla si rilasci. I fattori principali che stimolano il rilasciamento sono:

41
A. CCK, oltre a promuovere le contrazioni della cistifellea (stimolo principale) induce anche un rilasciamento dello sfintere che però è del tutto insufficiente. B. Le contrazioni stesse della parete della cistifellea interessano anche i muscoli dello sfintere che inizia a rilasciarsi, ma anche questo stimolo è insufficiente. C. Lo stimolo principale è invece il rilassamento indotto dal propagarsi delle onde peristaltiche intestinali a livello del duodeno
 induce anche un rilasciamento dello sfintere che però è del tutto insufficiente. B. Le contrazioni stesse della parete della cistifellea interessano anche i muscoli dello sfintere che inizia a rilasciarsi, ma anche questo stimolo è insufficiente. C. Lo stimolo principale è invece il rilassamento indotto dal propagarsi delle onde peristaltiche intestinali a livello del duodeno.")
43
Il precursore dei sali biliari è il colesterolo che è assunto con la dieta o è sintetizzato a livello epatico. Il colesterolo è insolubile in acqua, ma a livello epatico è complessato con lecitina e sali biliari a formare micelle e solubilizzato. Il colesterolo è convertito in acido colico o chenodesossicolico. Questi acidi si combinano con glicina o taurina (in minor quantità) formando glico- e tauro-coniugati i cui sali sono secreti nella bile.
 formando glico- e tauro-coniugati i cui sali sono secreti nella bile..")
44
A livello duodenale i sali biliari contenuti nella bile hanno due importanti effetti: Emulsione di particelle di grassi che vengono ridotti in minuscole parti facilmente attaccabili dalle lipasiEmulsione di particelle di grassi che vengono ridotti in minuscole parti facilmente attaccabili dalle lipasi I sali biliari aiutano l’assorbimento di acidi grassi, monogliceridi, colesterolo e altri lipidi nel tratto intestinale formando micelle solubili.I sali biliari aiutano l’assorbimento di acidi grassi, monogliceridi, colesterolo e altri lipidi nel tratto intestinale formando micelle solubili.

45
Circa il 94% dei sali biliari sono riassorbiti a livello intestinale e riportati al fegato attraverso la circolazione portale. Il rimanente 6% è riformato nel fegato a partire dal colesterolo.

46
94% 6%

47
Nella cistifellea in seguito a concentrazione della bile può succedere che il colesterolo precipiti a formare i calcoli. I motivi che portano a questo sono: Eccessivo riassorbimento di acqua dalla bile Eccessivo riassorbimento di lecitina e sali biliari Eccessiva secrezione di colesterolo nella bile Infiammazione dell’epitelio della cistifellea

48
Intestino tenue: Secrezione di muco dalle ghiandole del Brunner situate nei primi centimetri del duodeno (fra piloro e papilla di Vater). È un muco alcalino a funzione lubrificante e protettiva dall’acidità del chimo. La secrezione è indotta da stimoli chimici irritanti della mucosa, da secretina, da stimolazione vagale.

49
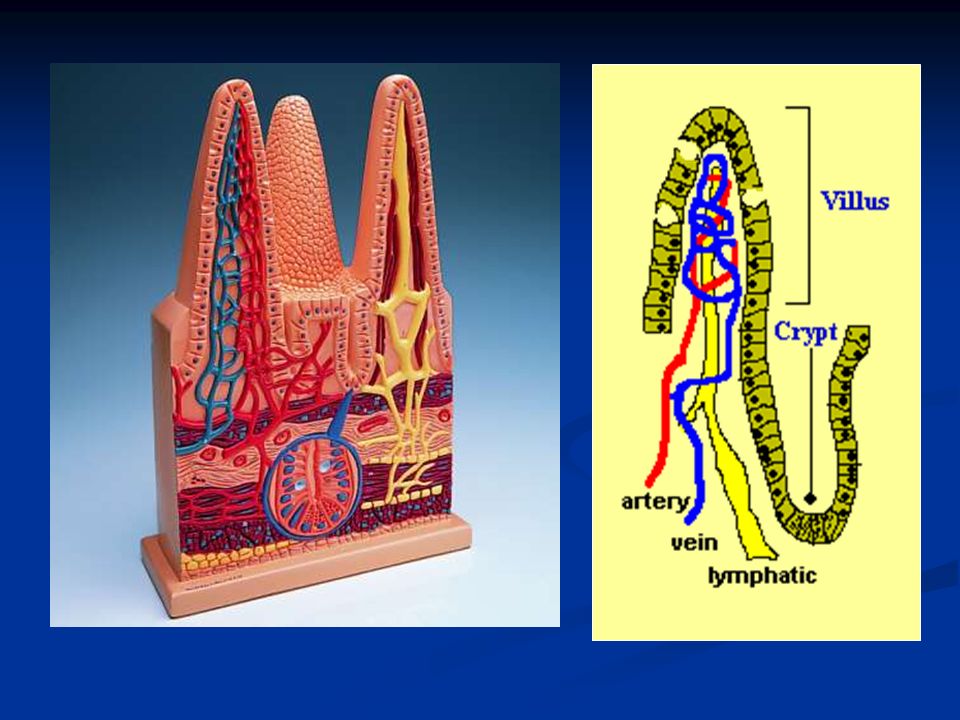
Per tutta la lunghezza del piccolo intestino troviamo le cripte di Lieberkühn che si trovano fra i villi intestinali e secernono muco e grandi quantità di acqua ed elettroliti. Questa acqua ed elettroliti sono poi riassorbiti dai villi; questa circolazione è utile per favorire l’ assorbimento di sostanze dal chimo.

50
Cellule di Paneth a funzione antimicrobica per l’intestino tenue. Secernono α-difensine contro batteri, funghi e anche alcuni virus. Cellule staminali che sono precursori degli enterociti Enterociti

51
Inoltre bisogna ricordare che gli enterociti della mucosa intestinale contengono enzimi digestivi fra cui: varie peptidasi: digestione di piccoli peptidi in singoli aa saccarasi, maltasi, isomaltasi, lattasi lipasi intestinale per separare i grassi neutri in glicerolo e acidi grassi

52
Intestino crasso: La mucosa dell’intestino crasso non ha villi, ma è anch’essa caratterizzata dalla presenza delle cripte di Lieberkühn. La secrezione è quasi esclusivamente mucosa a funzione protettiva e lubrificante in seguito a stimoli meccanici e irritativi. Non ci sono enzimi digestivi.

53
Digestione e assorbimento dei nutrienti

54
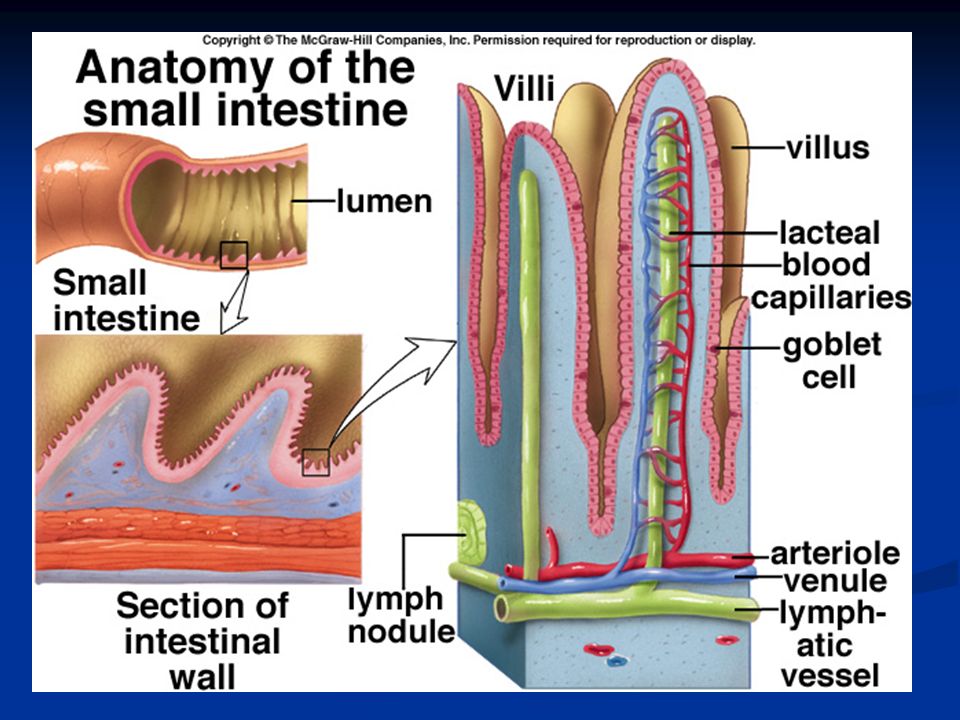
La quantità di fluido assorbita giornalmente a livello del tratto gi è di circa 8-9 litri, di cui la maggior parte è assorbita nel piccolo intestino. Valvulae conniventes: estendono la superficie di assorbimento di tre volte Villi intestinali: protrudono dalle valvulae conniventes aumentando la superficie di assorbimento di 10 volte Orletto a spazzola degli enterociti:sono circa 1000 microvilli per ogni cellula e aumentano la superficie di assorbimento di altre 20 volte.

57
CE= cellule epiteliali (enterociti) B= orletto a spazzola degli enterociti G= cellule di Goblet che secrenono muco
 B= orletto a spazzola degli enterociti G= cellule di Goblet che secrenono muco")
58
Suddivisione dei carboidrati I carboidrati sono suddivisi in tre categorie principali che sono: Monosaccaridi Disaccaridi Polisaccaridi

59
Digestione ed assorbimento dei carboidrati Lo scopo della digestione dei carboidrati è quello di liberare monosaccaridi dai disaccaridi e dai polisaccaridi complessi. Il processo inizia in bocca ad opera dell’amilasi salivare che funziona per valori di pH tra 6.6 e 6.8. A livello gastrico pertanto la sua azione cessa. La digestione dei carboidrati prosegue a livello duodenale ad opera dell’α-amilasi pancreatica: questi enzimi scindono i legami α,1-4 formando maltosio, maltotriosi, α-destrine e tracce di glucosio. Queste reazioni avvengono nel lume intestinale.

60
Vediamo quali sono i principali disaccaridi e le unità monomeriche che li costituiscono. DisaccarideUnità monomeriche LattosioGlucosio + Galattosio SaccarosioGlucosio + Fruttosio MaltosioGlucosio + Glucosio

61
Le rimanenti reazioni che portano alla formazione di monosaccaridi avvengono sulla superficie della parete intestinale dove troviamo disaccaridasi (maltasi, saccarasi, lattasi e isomaltasi) per la scissione dei disaccaridi che sono associate alla membrana plasmatica degli enterociti. Queste disaccaridasi, se presenti in quantità insufficienti per la digestione dei disaccaridi, generano situazioni di intolleranza che si manifestano con vomito, diarrea ed altri sintomi.

62
Fra queste la più nota intolleranza è quella per il lattosio dovuto ad una deficienza della lattasi, che è stata ampiamente riscontrata in tutte le popolazioni. La lattasi viene sintetizzata a partire dalla vita fetale e raggiunge livelli massimi alla nascita per poi diminuire progressivamente secondo fattori legati soprattutto a cause genetiche e non alla quantità di lattosio presente nella dieta come comunemente si crede.

63
Vediamo i meccanismi dell’assimilazione. I punti principali sono: La digestione enzimatica finale che libera monosaccaridi è dovuta ad enzimi che sono legati alla membrana plasmatica degli enterociti. Glucosio e galattosio sono trasportati per trasporto attivo e competono fra loro per l’utilizzo del trasportatore. Il fruttosio viene assorbito per diffusione facilitata. Il glucosio derivato dalla digestione di amido e lattosio è assorbito nel piccolo intestino solo per co- trasporto con Na +.

64
Assorbimento del glucosio: trasporto attraverso l’epitelio intestinale L’assorbimento del glucosio avviene dal lume attraverso l’epitelio e poi nel sangue. Il trasportatore che veicola glucosio e galattosio nell’enterocita è il trasportatore per gli esosi Na-dipendente, noto come SGLUT-1 che trasporta sia il glucosio che il sodio. Questo meccanismo prevede una serie di cambiamenti conformazionali che possono essere così schematizzati:

65
Il trasportatore è inizialmente rivolto verso il lume e può legare Na +, ma non glucosioIl trasportatore è inizialmente rivolto verso il lume e può legare Na +, ma non glucosio Il sodio si lega inducendo un cambiamento conformazionale che apre una tasca capace di legare il glucosioIl sodio si lega inducendo un cambiamento conformazionale che apre una tasca capace di legare il glucosio Il glucosio si lega ed il trasportatore si orienta nella membrana in modo che le tasche che alloggiano Na + e glucosio guardino verso l’interno della cellulaIl glucosio si lega ed il trasportatore si orienta nella membrana in modo che le tasche che alloggiano Na + e glucosio guardino verso l’interno della cellula Il sodio è rilasciato nel citoplasma causando un’instabilità del legame con il glucosioIl sodio è rilasciato nel citoplasma causando un’instabilità del legame con il glucosio Il glucosio è rilasciato ed il trasportatore riacquista la sua configurazione originale.Il glucosio è rilasciato ed il trasportatore riacquista la sua configurazione originale.

66
Out (lume) In (enterocita) Na + Glucosio Out (lume) In (enterocita) SGLUT-1
 In (enterocita) Na + Glucosio Out (lume) In (enterocita) SGLUT-1")
67
Assorbimento di glucosio e galattosio attraverso la mucosa del piccolo intestino SGLUT-1 Galattosio o Glucosio Trasportatore mobile del glucosio ENTEROCITA Glucosio GLUT 5 Glucosio capillare ATPADP Na + K+K+ K+K+ LUMESANGUE

68
Lipidi Esiste una gran varietà di lipidi che includono: Trigliceridi: formati da tre catene di acidi grassi legati a glicerolo Fosfolipidi: composti da acido fosfatidico (glicerolo + 2 acidi grassi + fosfato) Steroidi: derivati del colesterolo Colesterolo: non contiene acidi grassi, ma ha un anello sterolico che conferisce caratteristiche chimiche e fisiche simili a quelle dei lipidi
 Steroidi: derivati del colesterolo Colesterolo: non contiene acidi grassi, ma ha un anello sterolico che conferisce caratteristiche chimiche e fisiche simili a quelle dei lipidi")
69
Siccome la maggior parte dei grassi utilizzati per ricavare energia nell’organismo deriva dai trigliceridi (bersaglio principale nella riduzione calorica), vediamone la struttura base I tre acidi grassi più diffusi sono: Acido stearico: 18 atomi di C, saturo Acido oleico: 18 atomi di C, ha un doppio legame C=C Acido palmitico: 16 atomi di C, saturo
, vediamone la struttura base I tre acidi grassi più diffusi sono: Acido stearico: 18 atomi di C, saturo Acido oleico: 18 atomi di C, ha un doppio legame C=C Acido palmitico: 16 atomi di C, saturo")
70
I lipidi più diffusi negli alimenti sono i trigliceridi. Insieme a questi troviamo fosfolipidi, colesterolo e esteri del colesterolo. I trigliceridi iniziano ad essere digeriti, ma in maniera del tutto insignificante, a livello gastrico dove troviamo una lipasi gastrica. È responsabile meno del 10% della digestione dei lipidi.

71
Il primo passo nella digestione dei lipidi è la frammentazione dei globuli di grasso in piccole particelle che possano essere attaccate dalle lipasi del succo pancreatico. Questa azione emulsionante è fornita dagli acidi biliari, ma soprattutto dalla lecitina presente nella bile. Queste sono molecole caratterizzate da una parte polare solubile in acqua e da una grossa parte liposolubile.

72
La parte liposolubile si lega ai globuli di grasso e in presenza di movimenti di rimescolamento i globuli di grasso si frammentano in piccolissime particelle (emulsione) che possono essere attaccate dagli enzimi digestivi. Questi sono fortemente idrosolubili e quindi possono attaccare i globuli di grasso solo in superficie. L’enzima più importante è la lipasi pancreatica che viene secreta nella componente enzimatica del succo pancreatico in grandissime quantità. Esiste anche una lipasi enterica a livello degli enterociti, ma è di scarsa importanza. I trigliceridi sono scissi in acidi grassi liberi e 2-monogliceridi.

73
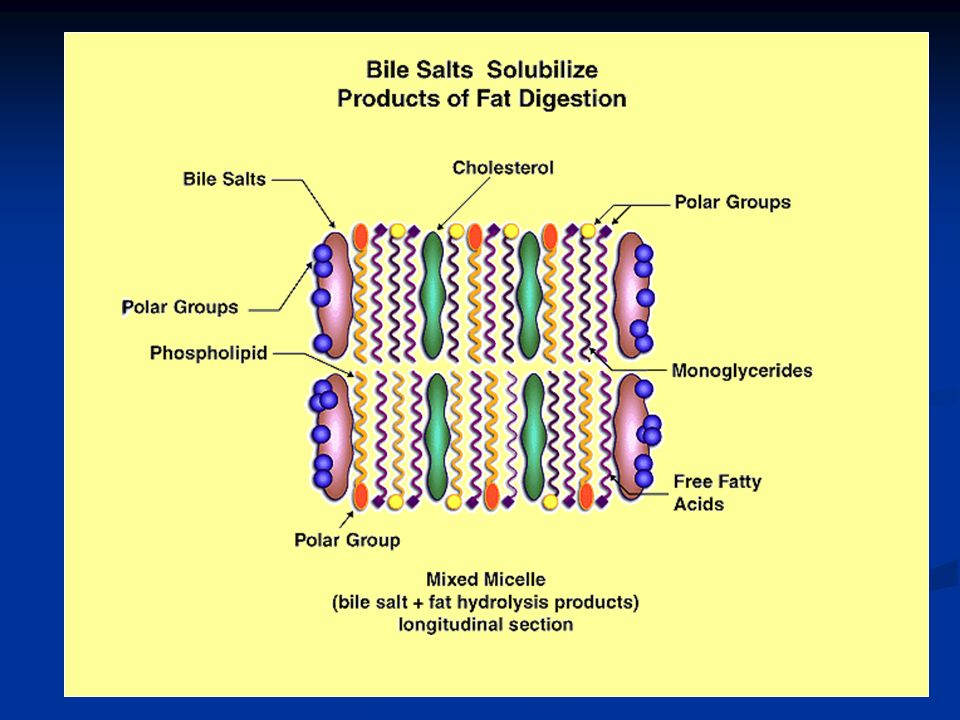
L’idrolisi dei trigliceridi è un processo facilmente reversibile, per cui, una volta digeriti è necessario allontanare gli acidi grassi liberi e i 2-monogliceridi, affinché la digestione possa continuare. I sali biliari provvedono a questo formando le micelle, strutture formate da 20-40 molecole di sali biliari in cui le teste polari dei sali sono rivolte verso l’esterno, mentre la parte liposolubile si lega ai prodotti della digestione lipidica e si rivolge verso l’interno. La concentrazione degli acidi grassi liberi e dei 2-monogliceridi diminuisce e la digestione può continuare.

75
Queste micelle funzionano anche da vettori per trasportare i prodotti della digestione verso l’orletto a spazzola degli enterociti. Quando i prodotti della digestione sono assorbiti a livello dell’orletto a spazzola, i sali biliari tornano nel chimo per questo meccanismo di trasporto.

76
Quando gli acidi grassi e i monogliceridi sono portati a contatto con la membrana plasmatica degli enterociti, questi si sciolgono nella membrana uscendo dalle micelle e liberando i sali biliari che tornano nel chimo ad inglobare nuovo materiale. All’interno dell’enterocita nel reticolo endoplasmatico liscio si formano nuovi trigliceridi che si aggregano. Successivamente nell’apparato del Golgi questi aggregati inglobano anche colesterolo e fosfolipidi. I fosfolipidi tendono a formare micelle in cui i trigliceridi restano inglobati: queste strutture sono liberate dal Golgi e si sciolgono nell’interno della cellula e per esocitosi fuoriescono dalla membrana basolaterale passando nel vaso linfatico sotto forma di chilomicroni.

77
I chilomicroni si riversano quindi nel torrente linfatico fino al dotto toracico per finire nella vena succlavia sx del collo e da qui al sangue. Circa l’80-90% dei grassi sono assorbiti in questo modo.

78
Le apoproteine sono fondamentali per l’esocitosi dei granuli

79
Digestione Si possono identificare tre processi principali: Intraluminale: si svolge nel duodeno e nel digiuno superiore ad opera delle lipasi pancreatiche stimolate dalla CCK. È fondamentale l’azione emulsionante della bile. La CCK stimola anche il rilascio di bile e, in parte, il rilasciamento dello sfintere di Oddi. Mucosale: la micella interagisce con la membrana plasmatica. Gli acidi grassi liberi e i monogliceridi sono trasportati attraverso la membrana del microvillo con trasporto passivo.

80
Secretoria: i chilomicroni formati sono veicolati per esocitosi attraverso la porzione laterale della cellula mucosale nello spazio extracellulare ed entrano nel sistema linfatico.

81
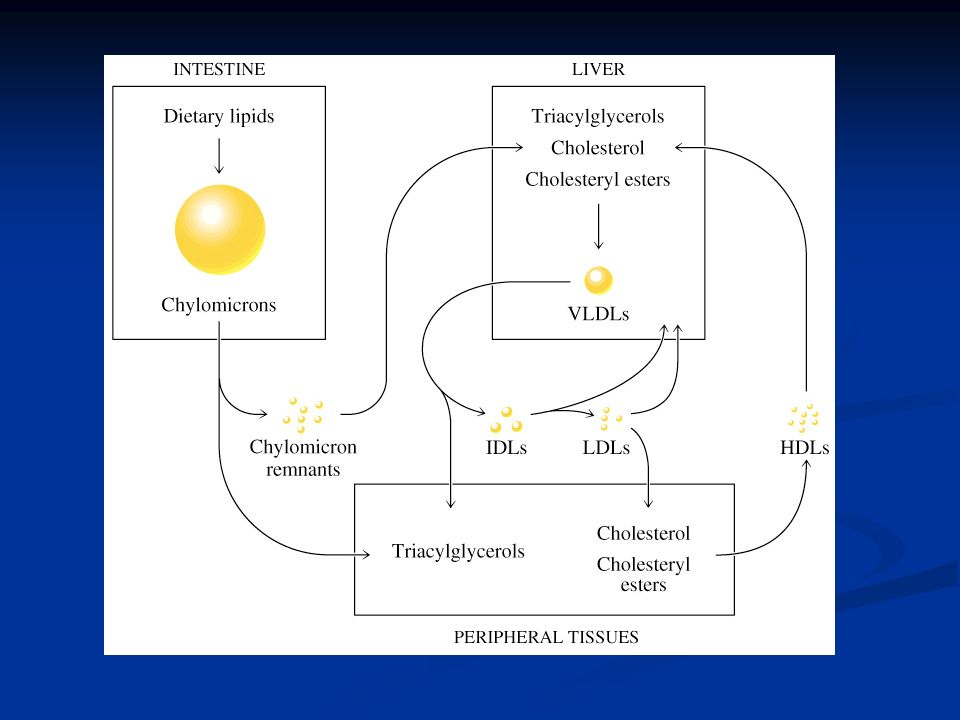
Lipoproteine I trigliceridi e gli esteri del colesterolo sono trasportati nel sangue dalle lipoproteine, complessi ad alto peso molecolare, che sono formate negli epatociti e negli enterociti. Le lipoproteine sono formate in buona parte da fosfolipidi che hanno la parte idrofilica orientata verso il sangue e la parte idrofobica verso l’interno della struttura. Il cuore (core) della struttura è formato da colesterolo. Esistono diversi tipi di lipoproteine che sono:
 della struttura è formato da colesterolo. Esistono diversi tipi di lipoproteine che sono:.")
82
Chilomicroni: assorbimento di trigliceridi e colesterolo assunti con la dieta derivanti dal processo digestivo Chilomicroni: assorbimento di trigliceridi e colesterolo assunti con la dieta derivanti dal processo digestivo Very-low-density-lipoproteins (VLDL): prodotte a livello epatico trasportano il colesterolo endogeno e quello derivato dai chilomicroni Very-low-density-lipoproteins (VLDL): prodotte a livello epatico trasportano il colesterolo endogeno e quello derivato dai chilomicroni Low-density-lipoproteins (LDL): prodotto metabolico delle VLDL. Perdita di trigliceridi ad opera della lipoproteina lipasi e aumento percentuale del colestrolo. Low-density-lipoproteins (LDL): prodotto metabolico delle VLDL. Perdita di trigliceridi ad opera della lipoproteina lipasi e aumento percentuale del colestrolo. High-density-lipoproteins (HDL): prodotte a livello epatico. High-density-lipoproteins (HDL): prodotte a livello epatico.
: prodotto metabolico delle VLDL. Perdita di trigliceridi ad opera della lipoproteina lipasi e aumento percentuale del colestrolo. High-density-lipoproteins (HDL): prodotte a livello epatico. High-density-lipoproteins (HDL): prodotte a livello epatico..")
83
Lipoproteina: core formato da colesterolo esterificato e trigliceridi. Shell formata da fosfolipidi e apoproteine con funzioni enzimatiche e recettoriali

84
HDL or high-density lipoprotein and LDL, low-density lipoprotein. HDL, known as good cholesterol, helps move cholesterol back to the liver for removal from the bloodstream. LDL, referred to as the bad cholesterol, helps cholesterol stick to artery walls. HDL: apoproteine A-I e A-II LDL: apoproteina B-100

85
Chilomicroni: nel circolo linfatico ed ematico si arricchiscono di specifiche apoproteine (Apo-E, Apo-C I, II, III), cedute dalle HDL. Sotto l’azione della lipoproteina lipasi presente sull’endotelio capillare del tessuto adiposo, sotto regolazione dell’insulina, la quantità dei trigliceridi è ridotta del 90%. I chilomicroni privati dei trigliceridi, sono captati dagli epatociti, che riconoscono la Apo-E, per recuperare i resti di colesterolo.

86
LDL: derivano dalle VLDL, che sono il meccanismo di trasporto per i lipidi sintetizzati a livello epatico. Sono ricche in colesterolo che trasportano ai vari distretti corporei entrando nelle cellule. Le cellule che hanno bisogno di colesterolo espongono il recettore B-100 che lega Apo-E.

87
HDL: Nell’uomo il colesterolo immesso in circolo dall’intestino (chilomicroni) e dal fegato (VLDL) supera le richieste per la produzione degli ormoni steroidei per cui una buona parte del colesterolo viene ricondotta al fegato e da qui passa alla bile: trasporto inverso del colesterolo. Ciò è possibile grazie alle HDL (lipoproteine ad alta densità), secrete dall’intestino e dal fegato. Le HDL diffondono facilmente nell’interstizio e vengono a contatto con le cellule dell’intero organismo. E’ possibile che le cellule esprimano un recettore per le HDL quando hanno colesterolo da smaltire.
, secrete dall’intestino e dal fegato. Le HDL diffondono facilmente nell’interstizio e vengono a contatto con le cellule dell’intero organismo. E’ possibile che le cellule esprimano un recettore per le HDL quando hanno colesterolo da smaltire..")
89
Normalmente sono controllati, con normali prelievi di sangue, i livelli di colesterolo e LDL. Nel caso in cui il recettore per la apoproteina B-100 venga meno espresso, il colesterolo legato alle LDL si accumula nel plasma aumentando moltissimo il rischio di infarto. chilomicroniLDLVLDLHDL

90
Le proteine della dieta sono costituite da catene di aminoacidi legati fra loro da legami peptidici.

91
Le caratteristiche di ciascuna proteina sono determinate dalla sequenza di aa e dai legami che intercorrono fra essi. La digestione delle proteine inizia a livello gastrico ad opera delle pepsine, che sono attive in range di pH fra 2 e 3 e si inattivano per pH superiori a 5. Queste pepsine sono in grado di digerire il collagene, costituente principale del tessuto connettivo intercellulare della carne. Quando il collagene è digerito gli enzimi proteolitici possono meglio attaccare le proteine cellulari. Le pepsine iniziano solo il processo digestivo e garantiscono solo il 10-20% della digestione proteica.

92
La maggior parte della digestione proteica avviene a livello intestinale grazie al succo pancreatico. All’uscita dello stomaco si trovano peptoni e grossi polipeptidi. All’ingresso del duodeno gli enzimi tripsina, chimotripsina, carbossipeptidasi e proelastasi attaccano subito questi composti. Tripsina e chimotripsina rompono i polipeptidi in piccoli peptidi. La tripsina taglia sul carbossile degli aa basici, Lys e Arg; la chimotripsina taglia sul carbossile di Phe, Tyr, Trp altri grandi amminoacidi apolari La carbossipolipeptidasi estrae da questi peptidi aa singoli. La proelastasi è convertita in elastasi che digerisce le fibre di elastina che tengono insieme la carne.

93
Solo una piccola parte delle proteine è digerita in singoli aa. La maggior parte rimane sotto forma di dipeptidi, tripeptidi e peptidi più grandi. La digestione dei peptidi avviene ad opera delle peptidasi degli enterociti della mucosa duodenale e digiunale. La membrana cellulare dei microvilli è ricca di peptidasi che protrudono verso l’esterno, dove vengono in contatto con il chimo intestinale.

94
Fra queste ricordiamo la aminopolipeptidasi e varie dipeptidasi che formano rispettivamente tri- e dipeptidi e singoli aa. Questi composti sono tutti trasportati facilmente attraverso la membrana del microvillo all’interno dell’enterocita. Sono poi le peptidasi dell’enterocita a spezzare gli ultimi legami peptidici e formare gli aa che passano nel sangue attraverso la membrana baso-laterale. Più del 99% del materiale assorbito è costituito da aa. Solo pochissimi tri- e dipeptidi e qualche molecola di piccole proteine passano nel sangue. Queste possono però essere sufficienti a causare serie reazioni immunitarie.

95
Il trasporto di singoli aa e di di- e tri-peptidi attraverso la membrana dell’enterocita avviene soprattutto nell’intestino tenue, grazie a meccanismi di co- trasporto con il Na + come già visto per glucosio e galattosio. Vale lo stesso meccanismo di legame e rotazione del trasportatore in membrana. Si sfrutta il gradiente per il sodio diretto verso l’interno della cellula. Si parla di co-trasporto o trasporto attivo secondario di aa o peptidi.

96
Formazione delle feci

97
Abbiamo detto che attraverso la valvola ileo-cecale passano circa 1500ml al giorno di materiale. La maggior parte di acqua ed elettroliti sono riassorbiti, tanto che nelle feci si eliminano al giorno solo 100ml di acqua e 5mEq/l di Na + e Cl -. La prima metà dell’intestino crasso ha funzione di riassorbimento. Ha infatti capacità di riassorbimento attivo di sodio e il potenziale elettronegativo che si crea determina riassorbimento anche di cloro. Le giunzioni fra una cellula e l’altra sono molto più serrate che nel tenue e questo impedisce fenomeni di back-leak presenti invece nell’intestino tenue. L’assorbimento di acqua avviene per gradiente osmotico.

98
storageassorbimento

99
L’intestino crasso può arrivare ad assorbire 5-7l di fluido al giorno. Se la quantità di materiale che entra attraverso la valvola ileo-cecale o la secrezione intestinale eccedono questo valore, si va incontro a fenomeni di diarrea. Per esempio la tossina colerica o tossine batteriche determinano un eccesso di secrezione da parte delle cripte del Lieberkühn che risulta in una severa diarrea talvolta anche letale.

100
Nella prima parte del colon sono presenti numerosi batteri capaci di digerire la cellulosa fornendo quindi un piccolo ulteriore aumento dell’apporto calorico giornaliero. Altre sostanze che dipendono dall’attività batterica sono la vitamina K, la vitamina B 12, tiamina, riboflavina e vari gas che contribuiscono al flatus nel colon (CO 2, CH 4 ).
..")
101
Le feci sono così formate: i. 75% acqua ii. 25% materiale solido che comprende: - 30% batteri morti - 10-20% grassi - 10-20% materiale inorganico - 2-3% proteine - 30% materiale non digerito e componenti secchi dei succhi digestivi come i sali biliari Il colore è determinato da stercobilina e urobilina e l’odore prodotto dall’azione batterica dovuto a sostanze quali H 2 S,mercaptano, indolo.

Presentazioni simili



















.>")