Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Geni e malattie Ciò che rende possibile lo sviluppo di una malattia in un ospite e’ la presenza nel patogeno di uno o piu geni di patogenicità, di virulenza e di specificitaùà per quello specifico ospite I geni di virulenza in un patogeno sono usualmente specifici per uno o pochi tipi di piante-ospiti Tale specificità fa si che un patogeno sia in grado di attaccare un limitato numero di ospiti Dal punto di vista della pianta non suscettibile all’attacco del patogeno questa viene chiamata resistenza non ospite Tutte le piante hanno delle difese pre-esistenti e indotte che conferiscono resistenza contro la maggior parte dei patogeni

2
Come compaiono nuovi geni di resistenza o virulenza?
Ogni specie vegetale sembra essere suscettibile a un basso numero di patogeni (meno di 100 per pianta) Nonostante possano essere attaccate da diversi patogeni, possono esistere degli individui all’interno delle singole specie che presentano un elevato grado di resistenza e che quindi presentano solo sintomi minori Perche’ molte piante non vengono attaccate dai propri patogeni e perche’ molte di quelle attaccate non muoiono? Accade perche’ le piante attraverso l’evoluzione hanno acquisito uno o numerosi geni di resistenza che le proteggono dall’infezione o da sintomi piu’ severi Quando compare un nuovo gene di resistenza ad un patogeno la pianta diventa resistente a tutti I precedenti individui di patogeno Tali patogeni hanno di solito uno o piu geni di virulenza e se non compare nessun nuovo gene In risposta al gene di resistenza l’infezione non puo avvenire Come compaiono nuovi geni di resistenza o virulenza? Mutazione Ricombinazione Flusso genico Deriva genetica
 Nonostante possano essere attaccate da diversi patogeni, possono esistere degli individui all’interno delle singole specie che presentano un elevato grado di resistenza e che quindi presentano solo sintomi minori. Perche’ molte piante non vengono attaccate dai propri patogeni e perche’ molte di quelle attaccate non muoiono Accade perche’ le piante attraverso l’evoluzione hanno acquisito uno o numerosi geni di resistenza che le proteggono dall’infezione o da sintomi piu’ severi. Quando compare un nuovo gene di resistenza ad un patogeno la pianta diventa resistente a tutti I precedenti individui di patogeno. Tali patogeni hanno di solito uno o piu geni di virulenza e se non compare nessun nuovo gene In risposta al gene di resistenza l’infezione non puo avvenire. Come compaiono nuovi geni di resistenza o virulenza Mutazione. Ricombinazione. Flusso genico. Deriva genetica.")
3
Tipi di resistenza ai patogeni
Le piante possono essere resistenti a un patogeno perchè: Appartengono ad un gruppo tassonomico al di fuori di quello ospite di un patogeno (resistenza non ospite) Hanno dei geni di resistenza (geni R) diretti contro geni di avirulenza del patogeno (resistenza vera o gene per gene) Per vari motivi scappano o tollerano l’infezione (resistenza apparente) Comunque anche all’interno della stessa specie c’e una considerevole variabilita’ nei confronti della suscettibilita ad un patogeno e di virulenza da parte del patogeno stesso
 Hanno dei geni di resistenza (geni R) diretti contro geni di avirulenza del patogeno (resistenza vera o gene per gene) Per vari motivi scappano o tollerano l’infezione (resistenza apparente) Comunque anche all’interno della stessa specie c’e una considerevole variabilita’ nei confronti della suscettibilita ad un patogeno e di virulenza da parte del patogeno stesso.")
4
Types of Resistance Non-host resistance
i.e., most plants are not hosts to most pathogens Partial, quantitative, or horizontal resistance These are the types of resistance that are most influenced by the disease triangle. Generally controlled by many genes for many plant processes or structures Race-specific or vertical resistance This type of resistance is an “either/or” type of resistance. Generally controlled by a single gene

5
Resistenza apparente La resistenza apparente alla malattia in piante note per essere suscettibili nei confronti di un determinato patogeno e’ dovuta alla “fuga” dalla malattia o alla tolleranza alla malattia stessa La fuga: la pianta suscettibile non rimane infetta perche’ I 3 fattori necassari per lo sviluppo della malattia (ospite suscettibile, patogeno virulento, ambiente favorevole) non coincidono e non interagiscono per il tempo giusto o per una durata sufficiente La tolleranza: e’ l’abilita della pianta di produrre una buon raccolto anche quando attaccata da un patogeno ed e’ il risultato di caratteristiche specifiche ed ereditabili che consentono loro di sopportare l’infezione senza subire profonde alterazioni fisiologiche
 non coincidono e non interagiscono per il tempo giusto o per una durata sufficiente. La tolleranza: e’ l’abilita della pianta di produrre una buon raccolto anche quando attaccata da un patogeno ed e’ il risultato di caratteristiche specifiche ed ereditabili che consentono loro di sopportare l’infezione senza subire profonde alterazioni fisiologiche.")
6
Complete and Partial Resistance
There are two fundamentally different mechanisms of disease resistance. Complete resistance AKA vertical resistance Highly specific (race specific) Involves evolutionary genetic interaction (arms race) between host and one species of pathogen. QUALITATIVE Partial Resistance AKA horizontal resistance Not specific- confers resistance to a range of pathogens QUANTITATIVE
 Involves evolutionary genetic interaction (arms race) between host and one species of pathogen. QUALITATIVE. Partial Resistance. AKA horizontal resistance. Not specific- confers resistance to a range of pathogens. QUANTITATIVE.")
7
Complete and Partial Resistance
There are two fundamentally different mechanisms of disease resistance. Complete resistance Disease severity class Frequency % Partial resistance Disease severity class Frequency %

8
Gene-for-Gene theory of Complete Resistance
Pathogen has virulence (a) and avirulence (A) genes A a Plant has resistance gene RR rr If the pathogen has an Avirulence gene and the host a Resistance gene, then there is no infection
 and avirulence (A) genes. A. a. Plant has resistance gene. RR. rr. If the pathogen has an Avirulence gene and the host a Resistance gene, then there is no infection.")
9
Gene-for-gene interactions vertical resistance
Avirulence genes first identified by H.H. Flor in 1950’s in studies on rust of flax Pathogens with avr genes are not pathogenic on plants with R genes Both avr and R genes are dominant genes

10
Gene-for-Gene theory of Complete Resistance
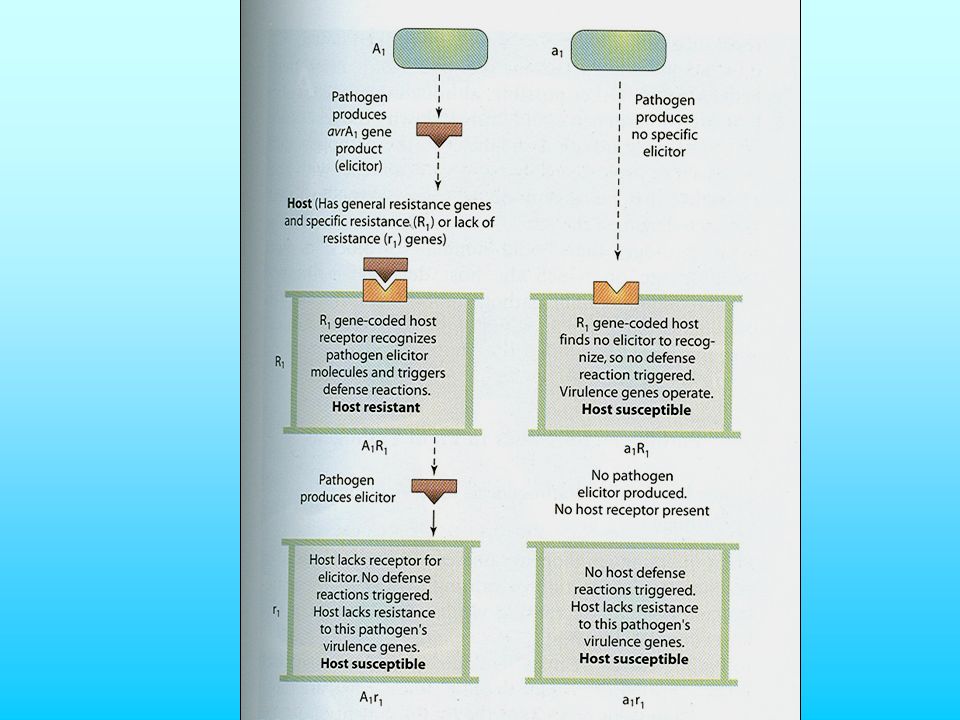
The Avirulence gene codes for an Elicitor molecule or protein controlling the synthesis of an elicitor. The Resistance gene codes for a receptor molecule which ‘recognises’ the Elicitor. A plant with the Resistance gene can detect the pathogen with the Avirulence gene. Once the pathogen has been detected, the plant responds to destroy the pathogen. Both the Resistance gene and the Avirulence gene are dominant

11
Gene-for-Gene theory of Complete Resistance
What is an elicitor? It is a molecule which induces any plant defence response. It can be a polypeptide coded for by the pathogen avirulence gene, a cell wall breakdown product or low-molecular weight metabolites. Not all elicitors are associated with gene-for-gene interactions. What do the Avirulence genes (avr genes) code for? They are very diverse! In bacteria, they seem to code for cytoplasmic enzymes involved in the synthesis of secreted elicitor. In fungi, some code for secreted proteins, some for fungal toxins. What does a resistance gene code for? The receptor for the specific elicitor associated with the interacting avr gene
 code for They are very diverse! In bacteria, they seem to code for cytoplasmic enzymes involved in the synthesis of secreted elicitor. In fungi, some code for secreted proteins, some for fungal toxins. What does a resistance gene code for The receptor for the specific elicitor associated with the interacting avr gene.")
12
Genetics of resistance:
Host cultivar Pathogen race Race-specific resistance: Resistance characterized by specific genetic interactions between host genotypes and pathogen genotypes. The ranking of cultivars from least to most resistant depends on the pathogen genotype (race) used. This type of resistance is commonly qualitative
 used. This type of resistance is commonly qualitative.")
13
Race-specific resistance is based on the Gene-for-Gene Hypothesis :
The product of a single gene in the host specifically recognizes the product of a gene in the pathogen. During an infection event, an interaction between these products results in incompatibility (i.e., a hypersensitive response and programmed cell death). ‘’
. ‘’")
14
Gene-for-Gene Hypothesis
Host: ‘R’–gene’= recognizer this gene produces a product that ‘recognizes’ usually an R-gene is a dominant gene ‘dominant’ means that receiving a copy of the gene from only one parent is sufficient to trigger the recognition response in the plant Pathogen: ‘AVR’ gene’= avirulence gene this gene produces an elicitor – ‘elicits the HR’ an AVR-gene is dominant dominant means that receiving a copy of the gene from only one parent is sufficient to elicit a recognition response in the host

15
Geni di patogenicita’ I patogeni hanno diversi geni che sono essenziali per causare malattia o per aumentare la virulenza I fattori di patogenicita’ sono il prodotto dei geni di patogenicita’ (pat) e di malattia (dsp) Questi geni includono quelli essenziali per il riconoscimento dell’ospite, per l’adesione ad esso e alle sue strutture, per la germinazione e la formazione di strutture di infezione, per la penetrazione e la colonizzazione dei tessuti vegetali I geni coinvolti nella sintesi e modificazione dei lipopolisaccaridi della parete dei batteri gram negativi condizionano il range di ospite Alcuni enzimi detti CWDE, alcune tossine, ormoni, polisaccaridi, proteinasi, siderofori e melanine sono prodotti dai patogeni e sono essenziali per causare infezione e sono quindi dei fattori di patogenicita’ Quindi I geni di patogenicita’ sono quei geni che rendono un particolare microrganismo un patogeno I prodotti dei geni di virulenza possono essere anche riconosciuti dall’ospite e alla lunga diventare fattori di avirulenza
 e di malattia (dsp) Questi geni includono quelli essenziali per il riconoscimento dell’ospite, per l’adesione ad esso e alle sue strutture, per la germinazione e la formazione di strutture di infezione, per la penetrazione e la colonizzazione dei tessuti vegetali. I geni coinvolti nella sintesi e modificazione dei lipopolisaccaridi della parete dei batteri gram negativi condizionano il range di ospite. Alcuni enzimi detti CWDE, alcune tossine, ormoni, polisaccaridi, proteinasi, siderofori e melanine sono prodotti dai patogeni e sono essenziali per causare infezione e sono quindi dei fattori di patogenicita’ Quindi I geni di patogenicita’ sono quei geni che rendono un particolare microrganismo un patogeno. I prodotti dei geni di virulenza possono essere anche riconosciuti dall’ospite e alla lunga diventare fattori di avirulenza.")
16
Geni di patogenicita’ nei funghi
Geni che controllano la produzione delle strutture di infezione Molti funghi producono appressori per penetrare la parete vegetale. Ad esempio l’appressorio di Magnaporthe grisea contiene melanina e I mutanti incapaci di sintetizzare iol pigmento non riescono a produrre la struttura di infezione. Un altro gene mpg1, che sintetizza per una idrofobina, e’ fondamentale per il funzioanmento dell’appressorio Geni che controllano I CWDE Cutinasi, famiglia multigenica Pectinasi, famiglia multigenica Geni che controllano la produzione di metaboliti secondari in risposta alle Fitoanticipine, sono prodotte dall’ospite. Sono glicosidi con proprieta simili al sapone che possono distruggere le membrane. La saponina, avenacina A1, e’ localizzata nell’ epidermide delle radici di avena. Il fungo Gaeumannomyces graminis infetta l’avena producendo l’enzima avenacina che degrada la saponina Fitoalessine, come la pisatina possono essere degradate dalla pisatina demetilasi che e’ prodotta dal fungo Nectria Geni che controllano la produzione di tossine Geni legati alla signal transduction La distruzione del gene PMK1 in M. grisea riduce la capacita’ di formare l’appressorio. Il gene CHK1 di Cochliobolus heterostrophus ha una patogenicità ridotta

17
Geni di patogenicita’ nei batteri
Adesione alla superficie della pianta Sistemi di secrezione 5 tipi di secrezione Secrezione di tossine (ABC transporter) Secrezione di proteine, enzimi, tossine e fattori di virulenza Il piu importanti per il patogeno, trasporta proteine effettrici nell cellula vegetale Trasporta macromolecole nella cellula dell’ospite, simili ai fattori di mobilizzazione plasmidica tra batteri Codifica per adesine CWDE Tossine batteriche EPS come fattori di patogenicita’ Sono fattori molto importanti sia diretttamente che indirettamente rappresentando una barriera di resistenza ai ROS Occlude le punteggiature areolate Fattori di conversione del fenotipo PhcA, un regolatore della trascrizione ce il prodotto dell’operone phcBRSQ che controlla I livelli di PhcA in dipendenza della densita’ cellulare. Celluel che contengono alti livelli di PhcA porducono molti fattori di virulenza (EPS, esoenzimi)se viene inattivato il ceppo diventa avirulento ma inizia a produrre PG, siderofori, Hrp, e proteine di mobilita’ Hrp genes Sono associati ai fattori avr e codificano per proteine dette piline che sono usate per costruire un sistema di trasporto di tipo 3 per rilasciare fattori avr.
 Secrezione di proteine, enzimi, tossine e fattori di virulenza. Il piu importanti per il patogeno, trasporta proteine effettrici nell cellula vegetale. Trasporta macromolecole nella cellula dell’ospite, simili ai fattori di mobilizzazione plasmidica tra batteri. Codifica per adesine. CWDE. Tossine batteriche. EPS come fattori di patogenicita’ Sono fattori molto importanti sia diretttamente che indirettamente rappresentando una barriera di resistenza ai ROS. Occlude le punteggiature areolate. Fattori di conversione del fenotipo. PhcA, un regolatore della trascrizione ce il prodotto dell’operone phcBRSQ che controlla I livelli di PhcA in dipendenza della densita’ cellulare. Celluel che contengono alti livelli di PhcA porducono molti fattori di virulenza (EPS, esoenzimi)se viene inattivato il ceppo diventa avirulento ma inizia a produrre PG, siderofori, Hrp, e proteine di mobilita’ Hrp genes. Sono associati ai fattori avr e codificano per proteine dette piline che sono usate per costruire un sistema di trasporto di tipo 3 per rilasciare fattori avr.")
18
Avirulence genes Can be extremely varied, almost anything can be an avirulence gene There are few common characteristics Avr genes tend to persist in populations Suggests an important role in pathogen physiology Sometimes they code for a virulence factor, but most have no role in pathogenicity

19
hrp genes Hrp genes (hypersensitive response and pathgenicity)
Found only in gram- bacteria Not avr genes but appear to be essential for the development of hypersensitive response and disease in non-resistant plants Appear to be involved in the type III secretion system of bacteria

20
Interazione avr-hrp-R
Il prodotto di avr1 induce un enzima extracellulare a produrre un elicitore che si muove liberamente attraverso la membrana batterica I prodotti di avr 2, e 4 così come le proteine effettore trasmesse attraverso il sistema di secrezione di tipi III si muovono attraverso I pori formati dalle proteine hrp e agiscono come elicitori I geni di patogenicità/malattia producono gli effettori

21
Resistance genes Of the genes characterized to data, the proteins coded for are quite similar Most have a leucine-rich domain (LRR) Resistance gene can be cytoplasmic or transmembrane proteins Once bound to the avr gene product, initiate the cascade leading to HR

23
Resistance or susceptibility
Gene for gene interaction -one gene Resistance or susceptibility genes in the plant R (resistant dominant) r (susceptible Recessive) A (avirulent dominant AR (-) Ar (+) Avirulence or virulence genes in the pathogen a (virulent recessive) aR (+) ar (+) Minus indicates an incompatible reaction (resistance) Plus indicates a compatible reaction (disease)
 r (susceptible. Recessive) A (avirulent. dominant. AR (-) Ar (+) Avirulence or virulence genes in the pathogen. a (virulent recessive) aR (+) ar (+) Minus indicates an incompatible reaction (resistance) Plus indicates a compatible reaction (disease)")
24
Resistance or susceptibility
Gene for gene interaction -two genes Resistance or susceptibility genes in the plant R1R2 R1r2 r1R2 r1r2 A1A2 A1a2 a1A2 a1a2 - + - + - + + Avirulence or virulence genes in the pathogen Minus indicates an incompatible reaction (resistance) Plus indicates a compatible reaction (disease)
 Plus indicates a compatible reaction (disease)")
25
A common test question:
For each of the boxes, indicate whether or not the races of a rust pathogen will succeed in infecting the host cultivars Avirulence (A) or lack-of-avirulence (? or ‘a’) genes possessed Race: A1A2A3A4 A1a2A3A4 a1A2A3A a1a2A3A4 a1a2a3a4 r1r2r3r4 R1r2r3r4 lack-of-resistance (? or ‘r’) genes in each cultivar Resistance (R) or r1R2r3r4 R1R2r3r4 R1R2r3R4 ‘R’, ‘A’ = active product present ‘?’, ‘a’, ‘r’ = active product absent
 or lack-of-avirulence ( or ‘a’) genes possessed. Race: A1A2A3A4 A1a2A3A4 a1A2A3A4 a1a2A3A4 a1a2a3a4. r1r2r3r4. R1r2r3r4. lack-of-resistance ( or ‘r’) genes in each cultivar. Resistance (R) or. r1R2r3r4. R1R2r3r4. R1R2r3R4. ‘R’, ‘A’ = active product present ‘ ’, ‘a’, ‘r’ = active product absent.")
26
A common test question:
For each of the boxes, indicate whether or not the races of a rust pathogen will succeed in infecting the host cultivars Avirulence (A) or lack-of-avirulence (? or ‘a’) genes possessed Race: A1A2A3A4 A1a2A3A4 a1A2A3A a1a2A3A4 a1a2a3a4 r1r2r3r4 R1r2r3r4 yes yes yes yes yes no no yes yes lack-of-resistance (? or ‘r’) genes in each cultivar Resistance (R) or yes r1R2r3r4 R1R2r3r4 no yes no yes yes no no no yes yes R1R2r3R4 no no no no yes ‘R’, ‘A’ = active product present ‘?’, ‘a’, ‘r’ = active product absent
 or lack-of-avirulence ( or ‘a’) genes possessed. Race: A1A2A3A4 A1a2A3A4 a1A2A3A4 a1a2A3A4 a1a2a3a4. r1r2r3r4. R1r2r3r4. yes. yes. yes. yes. yes. no. no. yes. yes. lack-of-resistance ( or ‘r’) genes in each cultivar. Resistance (R) or. yes. r1R2r3r4. R1R2r3r4. no. yes. no. yes. yes. no. no. no. yes. yes. R1R2r3R4. no. no. no. no. yes. ‘R’, ‘A’ = active product present ‘ ’, ‘a’, ‘r’ = active product absent.")
27
avr/R gene combinations to date
Tomato: avrD/Pto Pseudomonas syringae pv. tomato Tobacco: replicase proteins/N gene TMV, but also other biotrophs Rice: avrpi-ta/Pi-ta Magnaporthe grisea Tomato: Avr2, Avr3, Avr9/Cf2, Cf4, Cf9 Cladosporium fulva Arabidopsis: avrpm1-avrB/RPM1 Pseudomonas syringae pv maculicola and pv glycinea

28
Avirulence Gene Function – what is currently known
Avr Protein Pathogen Homology with and/or possible virulence function * Location in the plant cell Common domain for avirulence and virulence function Location of the matching R gene * AvrPphC Pseudomonas syringae pv. phaseolicola Interferes with the plant defense response NK ‡ NK NK AvrPphF Pseudomonas syringae pv. phaseolicola Interferes with the plant defense response NK NK NK AvrRpt2 Pseudomonas syringae pv. tomato Interferes with the plant defense response NK NK NK AvrRpm1 Pseudomonas syringae pv. maculicola Enhances growth Plasma membrane [ NK Plasma membrane AvrPto Pseudomonas syringae pv. tomato Enhances growth Degradation of resistance gene Plasma membrane No NK AvrBst Xanthomonas campestris pv. campestris Ubiquitin-like protease NK Yes NK AvrXa7 Xanthomonas oryzae pv. oryzae Transcriptional activation domain required for virulence Nucleus Yes NK AvrBs2 Xanthomonas campestris pv. vesicatoria Agrocinopine synthase (from Agrobacterium tumefaciens) NK Recognition region localized in the portion with highest homology to synthase NK AvrBs3 Xanthomonas campestris pv. glycinea Transcriptional activation domain required for virulence Nucleus Yes NK PthA Xanthomonas citri Transcriptional activation domain required for virulence/causes cell hyperplasia [53] Nucleus NK NK Avr9 Cladosporium fulvum NK Exoplasmic NK External plasma membrane
 NK Recognition region localized in the portion with highest homology to synthase. NK AvrBs3 Xanthomonas campestris pv. glycinea Transcriptional activation domain required for virulence Nucleus Yes NK PthA Xanthomonas citri Transcriptional activation domain required for virulence/causes cell hyperplasia [53] Nucleus NK NK Avr9 Cladosporium fulvum NK Exoplasmic NK External plasma membrane")
29
AvrRpm1 and avrB localise to the plant cell membrane.
Protoplasts from rpm1 plants were transformed with plasmid that express either avr-GFP or avrG2A-GFP constructs. Control transformations were with GFP alone. avrRpm1 N-terminal fatty acylation sequence avrRpm1 MGCVSSTSR Soc3a MGCSVSKKK Ca sensor G2A = glycine change to alanine Loss in avirulence

30
cloned resistance genes
Protein structure of cloned resistance genes C N Pto tomato; bacterial resistance Xa21 rice; bacterial resistance Hs1 sugar beet; nematode res. Cf9, Cf2 tomato; fungal resistance L6 flax; fungal resistance RPS2, RMP1 Arabidopsis; bac. res. N tomato; viral resistance Prf tomato; bacterial resitance Membrane anchor site Serine/threonine protein kinase domain Signal peptide Leucine-rich repeat Transmembrane domain Conserved motif Leucine zipper domain DNA binding site

31
Reazione R-avr

32
Hypothesized physical model for how R-gene and Avr-gene products interaction on plasma membrane of host cells Pathogen protein (AVR) interacts with host R-gene protein (Cf9), which triggers the cell death response (hypersensitivity) from pathogen = R-gene product From: MHAJ Joosten and PJGM de Wit Annual Review of Phytopathology Vol. 37:335

33
Pathogen ? Host is Rz,Rz or Rz,? ?,? P H AVRz,AVRz or AVRz, ? ?,?
Gene-for-Gene Hypothesis Host is Rz,Rz or Rz,? ?,? AVRz,AVRz or AVRz, ? ?,? H P ? Resistance Disease Pathogen Specific recognition in upper left box only

34
What if the pathogen is a necrotrophic fungus that produces a host selective toxin?
S,S or S,? ?, ? Tox, Tox or Tox, ? ?, ? Disease!! Pathogen P P ? H Resistant Gene-for-gene II Tox = gene for toxin S = gene for receptor Specific recognition in upper left box only ? ? ? H Resistant Resistant

35
How many races? Host cultivar Pathogen race Three cultivars, each with a different R gene yields 8 races (23) Ten cultivars, each with a different R gene could discern a possible 1024 races (210) !! The set of cultivars used to characterize races is called a ‘differential set’
 !! The set of cultivars used to characterize races is called a ‘differential set’")
36
L’interazione PG/PGIP
Le poligalatturonasi sono enzimi del patogeno in grado di idrolizzare la compenete pectica della lamella mediana Le PGIP, polygalcturonase inhibiting proteins, sono delle proteine LRR che legano le PG del patogeno Questo legame fa funzionare “a singhiozzo” le PG liberando in tal modo degli oligomeri di acido poligalatturonico gli OG La lunghezza degli OG non e’ casuale ma si l’interazione fra le 2 proteine crea 2 categorie di OG, I “lunghi”, monomeri di AG e I “corti”, residui Le 2 categorie di OG creati diventano degli elicitori endogeni di HR, ma non solo, dato che vengono creati anche durante il normale sviluppo della parete vegetale, servono anche come molecole segnale per reindirizzare lo sviluppo della pianta

37
Meccanismi responsabili del riconoscimento ospite-patogeno
Attualmente sono stati ipotizzati 4 meccanismi possibili: Il modello elicitore-recettore Il modello “dimero” Il modello “ion channel defense” Il modello sopressore-recettore

38
Modello elicitore-recettore
Questo modello suggerisce he almeno 2 gruppi di geni della pianta sono coinvolti: Il sensore che riconosce il patogeno L’effettore, e cioe I diversi geni di difesa della pianta Il gene avr del patogeno produce direttamente un elicitore oppure produce un enzima che rilascia un elicitore Il segnale e’ chiamato elicitore specifico Gli elicitori sintetizzati direttamente dal patogeno vengono chimati elicitori esogeni, se invece deriva da un prodotto di degradazione della pianta e’ endogeno Il sensore della pianta, il recettore, e’ geneticamente determinato da un gene razza-specifico di resistenza Dato che ad un fattore R corrisponde uno specifico fatore avr si parla di una coppia R-avr che detremina la resistenza razza-specifica Il recettore si trova presumibilmente sulla membrana plasmatica dove ha maggiore facilita’ di “incontrare” l’elicitore

39
Modello “dimero” Il problema del modello precedente e’ che per scatenare la resistenza, oltre ai geni R servono anche degli “effettori”, cioe’ altri geni che si accendono in conseguenza del legame recettore-elicitore Il modello a “dimero”invece si basa sul fatto che un singolo gene della pianta interagendo con un solo gene del patogeno scatena la reazione difensiva Si ha un riconoscimento tra molecola sensore e molecola segnale che non avviene nella membrana plasmatica ma nel citoplasma Questo dimero agisce come regolatore negativo, bloccando l’espressione dei geni che portano alla compatibilita’, agendo a livello trascrizionale Appena la compatibilita di base e’ bloccata, viene ri-accesa la resistenza di base Questo ultimo punto diverge dal modello elicitore-recettore. Infatti in questo caso non si parla di ri-attivazione di risposta difensiva ma piuttosto di nuovo meccanismo di difesa

40
Modello “ion channel defense”
Il modello si basa sul fatto che alcune proteine di membrana che funzionano come canali ionici abbiano la capacità di stimolare la resistenza razza-specifica Il modello dà una plausibile spiegazione di alcuni fenomeni osservati, tra cui la risposta ipersensibile, la morte cellulare in seguito ad HR, e l’attivazione di geni di difesa della pianta Un importante motivo per proporre tale modello nasce dall’evidenza che la prima reazione di una pianta soggetta a stress e’ l’alterazione dei potenziali di membrana in segutio a perdita di elettroliti Il riconoscimento tra un elicitore e la sua corrispondente proteina transmembrana agisce come stimolo per far aprire tale proteina legata a dei canali ionici Questa apertura porta ad un efflusso di ioni K+ e ad un influsso di ioni H+ e ad una conseguente depolarizzazione della membrana con occasionale distruzione della stessa Le sostanze liberate da tale evento stimolano la difesa nelle cellule vicine

41
Modello “ion channel defense”
Dopo il riconoscimento tra elictore e proteina di membrana quindi avviene questa depolarizzazione che come detto può provocare la distruzione del plasmalemma oppure genera una cascata di segnali In risposta a questi cambiamenti, infatti, si attivano le kinasi legate alla membrana, le fosfatasi, fosfolipasi e G protein Le kinasi attivano la NADPH ossidasi che trasformano l’O2 in anione superossido e altre specie attive dell’ossigeno I ROS agiscono direttamente sulla parete vegetale rinsaldandola, e attivano inoltre dei fattori di trascrizione che possono a loro volta attivare la PCD, l’HR e la sintesi di proteine ed enzimi di difesa Altre pathway specifiche vengono attivate e che portano alla produzione di lipoperossidi, acido jasmonico, etilene e acido salicilico Quest’ultimo e’ anche coinvolto nella diffusione sistemaica della resistenza attraverso la SAR

42
Modello “soppressore-recettore
Il modello propone che tutte le piante siano suscettibili all’attacco da qualsiasi patogeno, quindi tutte le piante esibiscono una compatibilita’ di base D’altronde tale fenomeno e’ contrastato da un elicitore generico prodotto da tutti I patogeni che porta alla resistenza di base Per colonizzare una particolare pianta il patogeno compatibile deve quindi produrre un soppressore di tale elicitore generico Come fa un patogeno ad essere riconosciuto? Una mutazione insorge nel soppressore ma non disturba il legame tra soppressore ed elicitore in tal modo non dà apparentemente nessuno svantaggio al patogeno D’altronde questa mutazione può portare ad una mutazione nel gene R che a questo punto e’ in grado di riconoscere l’elicitore nonstante la presenza del soppressore Quindi avvenendo il riconoscimento tra elicitore e gene R la reazione diventa incompatibile re

43
Model for the action of Xa21 (rice blight resistance gene)
Leucine-rich receptor Transmembrane domain Kinase Membrane Elicitor Signal transduction ([Ca2+], gene expression) Cell Wall Plant Cell
 Cell Wall. Plant Cell.")
44
The arms race explained
An avirulence genes mutates so that it’s product is no longer recognised by the host resistance gene. It therefore becomes a virulence gene relative to the host, and the pathogen can infect. The host resistance gene mutates to a version which can detect the elicitor produced by the new virulence gene.

45
Two General Categories of Defense:
Structural Characteristics Biochemical/Physiological Mechanisms Defense can take the form of both the above & may differ according to age of the plant, environment, stress, etc.

46
Plant Defense mechanisms – 2 types
A. Pre-existing B. Formed in Response to Infection

47
Getting inside the host is a critical
step is disease development Fungal pathogens use ‘contact and chemical tropisms’ to guide their way to site where infection is possible Pathogens use several tools to establish a parasitic relationship: enzymes, growth regulators, toxins Appreciate that pathogens are highly evolved to enter and establish a parasitic relationship with their host Use the timeline as is guide to when events relating to attack and counterattack take place

48
Plants respond immediately to a pathogen’s attempt to get inside :
Wound plug (papillae) formation as a result of needle injury:
 formation as a result of needle injury:")
49
TIME-LINE OF INFECTION
Outside of host Inside of host Fungal pathogen: Preinfection: Germination Germ tube search Appressorium formation Penetration peg Postinfection Haustorium formation (biotroph) Toxin formation (necrotroph) Detoxification of phytoalexins Reproduction Host: General induced Papillae Cork & lignin Systemic defenses: formation layers acquired resistance Specific recognition: Hypersensitivity Phytoalexins structural CONSTITUTIVE DEFENSES chemical
 Toxin formation (necrotroph) Detoxification of phytoalexins. Reproduction. Host: General induced Papillae Cork & lignin Systemic. defenses: formation layers acquired. resistance. Specific recognition: Hypersensitivity Phytoalexins. structural CONSTITUTIVE DEFENSES chemical.")
50
Host defenses Constitutive Induced
Structural Biochemical Structural Biochemical wax pre-formed cork layer hypersensitivity cuticle inhibitors: thickness abscission layer phytoalexins phenolics shape and others gums systemic acquired activity of resistance stomata papillae anti-microbial tyloses enzymes

51
Pre-existing Defenses
The first line of defense against a pathogen is the plant surface!! Trichomes – hairs- may prevent pathogen from reaching cell surface; or reduce water retention.

52
The leaf surface Epicuticular waxes: difficult to penetrate- lessens wetability. Cuticle: difficult to penetrate. Guard cells: shape, density and size- may make it difficult for pathogens to enter. e.g. wheat rust – some only open late in the day when rust can only invade during night or early morning. Thickness of cell outer wall – may prevent entry or spread of disease.

53
Pre-existing chemical defenses
Fungitoxic exudates Secreted by plants on their surface, catechol, coumarins and phenolics Intercellular inhibitors Phenolics compounds like tannins Other compounds like dienes (fatty acid-like compounds Fungal enzyme inhibitors, catechin Antifungal enzymes, glucanases, chitinases

54
Other pre-existing factors
Lack of recognition factors Pathogen fails to respond with critical processes Lack of receptor or sensitive site Pathogenicity factor has no effect Lack of essential substances Certain growth factors or sugars necessary for pathogen growth

55
Constitutive biochemical defense
Onion smudge In contrast to red and yellow onions, white onions do not contain significant quantities of certain phenolic chemicals (one is catechol). If present, these phenolics confer resistance to onion smudge disease. susceptible resistant
. If present, these phenolics confer resistance to onion smudge disease. susceptible. resistant.")
56
Induced structural defenses
Abscission layer Cork layer The formation of cork or abscission layers can limit the size of lesions, and consequently the extent of damage that can be caused by a single infection

57
Induced structural defenses
Tyloses are overgrowths of living cells that protrude via pits into xylem vessels blocking the vascular system. If they form abundantly and quickly, they can stop the spread of vascular wilt pathogens. Their formation is triggered by a ‘stress condition’.

58
d. Gums – deposition of gums (carbohydrate/protein) in intercellular spaces. Forms an impenetrable barrier. Pathogen is isolated and dies. Common in stone fruit.

59
Cellular Defense Structures
- changes in the cell wall - wall appositions/papillae Response to attempted invasion very fast. Composed of: a. Callose b. Cellulose c. Lignin – very resistant

60
- encasement – hyphae or haustoria are surrounded by thickened cell wall

61
Inducible plant defense mechanisms
Requires recognition of pathogen by plant A chemical specific recognition Pathogen elicitors, include toxins, carbohydrates, fatty acids, proteins, etc. Host plant receptors Often transmembrane proteins Results in defense response cascade

62
Necrotic structural defense response
Hypersensitive response is a structural response (also a biochemical response) a. Host is penetrated; pathogen contacts protoplast b. Nucleus migrates to site, disintegrates, cell membranes are disrupted, host cell dies. c. Cell browns and pathogen dies Important defense against obligate parasites, nematodes, some bacteria, some facultative saprobic fungi Usually, pathogen is dependent on living tissue; thus disease is limited.
 a. Host is penetrated; pathogen contacts protoplast. b. Nucleus migrates to site, disintegrates, cell membranes are disrupted, host cell dies. c. Cell browns and pathogen dies. Important defense against obligate parasites, nematodes, some bacteria, some facultative saprobic fungi. Usually, pathogen is dependent on living tissue; thus disease is limited.")
63
Necrotic structural defense response

64
Induced Metabolic Defenses: Horizontal resistance
Many of the same compounds associated with pre-existing defense response are important in inducible response The primary difference is that the presence and amount of such compounds is a result of interaction between pathogen and plant, i.e., a result of chemical signals…elicitors

65
Induced Metabolic Defenses: Horizontal resistance
Stimulated by elicitors 1. Host elicitors: cell wall components – glucose 2. Pathogen elicitors: cell wall components – beta-gucans, chitin 3. Heavy metal ions: Cu, Cd, Zn 4. Temperature induction – cold hardening, heat shock. Also other abiotic stresses. 1. Phytoalexins – are toxic substances produced only after stimulation by pathogenic microorganisms, chemical or mechanical injury such as insect feeding. Most inhibit growth of fungi, but some are active against bacteria and nematodes. Phaseollin, pisatin, glyceollin, gossypol, capsidiol

66
Induced Metabolic Defenses: Horizontal resistance
2. Increase Phenols Many different phenolic chemicals accumulate in cells and kill pathogens 3.Oxidize phenols to quinones: more toxic to microbes 4. Substrates resisting enzymes of the pathogen. Pectin is susceptible to enzymatic decay, but is more resistant if complexed with divalent cations (Ca or Mg)
")
67
Induced Metabolic Defenses: Horizontal resistance
5. Fungitoxic Cyanide-derivatives. HCN incorporated in wall linked by sugar molecule; when pathogen degrades wall release sulfur compounds and cyanide. Respiratory toxin. 6. Detoxification of Pathogen Toxins Fungal toxins that kill plant cells can be metabolized by enzymes produced by host 7. PR proteins – pathogenesis-related proteins Usually an antimicrobial enzyme: chitinases, beta-glucanases, proteinases, lysozymes

68
Hypersensitive Reaction/ Programmed Cell Death
In response to signals, evidence suggests that infected cells produce large quantities of extra-cellular superoxide and hydrogen peroxide which may 1/ damage the pathogen 2/ strengthen the cell walls Oxidative 3/ trigger/cause host cell death Burst Evidence is accumulating that host cell also undergo changes in gene expression which lead to cell death Programmed Cell Death

69
Plant Defense: Hypersensitive Response
Localized resistance mechanism due to recognition of a pathogen R (Resistance) genes from the host mediate recognition of effector proteins secreted from the bacterium Leads to rapid death of cells at the site of infection due to activation of defense genes
 genes from the host mediate recognition of effector proteins secreted from the bacterium. Leads to rapid death of cells at the site of infection due to activation of defense genes.")
70
Induced Metabolic Defenses:
Vertical resistance The hypersensitive response Many of the same compounds associated with hypersensitive response are associated with induced horizontal resistance. The primary difference here is that their production during HR is accompanied by programmed cell death…apoptosis

71
Induced Metabolic Defenses:
Vertical resistance Hypersensitive Reaction A type of programmed cell death – apoptosis. Invaded cells and some surrounding cells die. Characterized by: 1. recognition of pathogen avr genes by host resistance genes 2. increased ion movement across membranes 3. rapid burst of reactive oxygen species (ROS), superoxides and nitric oxide 4.phenolics, jasmonates and salicylic acid –signaling molecules 5.degradation of DNA, membranes disrupted, cell death. 6. Production of PR protein is induced, which further induces Systemic Acquired Resistance (SAR)
, superoxides and nitric oxide. 4.phenolics, jasmonates and salicylic acid –signaling molecules. 5.degradation of DNA, membranes disrupted, cell death. 6. Production of PR protein is induced, which further induces Systemic Acquired Resistance (SAR)")
72
L’HR e la PCD L’HR, che consiste in una morte cellulare localizzata molto rapida nel sito della tentata infezione, si trova in tutte le risposte di difesa: quelle mediate da uno o più geni R, nella resistenza non ospite e in molti casi di resistenza poligenica o quantitativa L’interazione tra un elicitore e il suo recettore porta immediatamente a eventi di signal transduction dovuto ad un rapido flusso di ioni transmembrana Questo flusso porta all’alcalinizzazione dell’apoplasto, alla formazione di ROS, alla produzione di NO, all’attivazione e all’attivazione di una cascate di segnale che coinvolge la pathway delle MAPK e alla fine ad una attivazione della trascrizione di geni di difesa Questa riprogrammazione trascrizionale risulta nella produzione e nel rilascio di composti antimicrobici o nella generazione di molecole segnale che possono agire a distanza instaurando così una risposta sistemica acquisita (SAR) L’HR non e’ sempre necessaria per la difesa. Se il sistema dei geni R riesce a confinare l’invasione allora la PCD non e’ piu’ necessaria. La PCD e’ comunque controlata da SA, JA, ET ed NO e dalla produzione di ROS. La distruzione del gene lsd1 (lesion simulating disease) porta alla generazione di piante che sono incapaci di bloccare la propagazione della PCD e quindi di confinarla al solo sito d’infezione. Le cellule delle piante Dlsd sono ipersensibili all’anione superossido, alla presenza del quale muoiono rapidamente (runaway cell death-rcd) Botrytis cinerea attacca a induce la rcd nelle cellule che presentano il gene lsd1. Infatti subito dopo l’infezione il fungo produce e rilascia H2O2 e superossido (convertito rapidamente dalla SOD in H2O2). Questi ROS attivano la HR e la PCD. In tal modo il fungo riesce a “simulare” l’HR per colonizzare I tessuti morti
 L’HR non e’ sempre necessaria per la difesa. Se il sistema dei geni R riesce a confinare l’invasione allora la PCD non e’ piu’ necessaria. La PCD e’ comunque controlata da SA, JA, ET ed NO e dalla produzione di ROS. La distruzione del gene lsd1 (lesion simulating disease) porta alla generazione di piante che sono incapaci di bloccare la propagazione della PCD e quindi di confinarla al solo sito d’infezione. Le cellule delle piante Dlsd sono ipersensibili all’anione superossido, alla presenza del quale muoiono rapidamente (runaway cell death-rcd) Botrytis cinerea attacca a induce la rcd nelle cellule che presentano il gene lsd1. Infatti subito dopo l’infezione il fungo produce e rilascia H2O2 e superossido (convertito rapidamente dalla SOD in H2O2). Questi ROS attivano la HR e la PCD. In tal modo il fungo riesce a simulare l’HR per colonizzare I tessuti morti.")
73
Hypersensitive response
Can be very localized, just a few cells and difficult to see, or it can involve a much larger area of the leaf surface Can lead to SAR

74
Local lesions – restricted HR

75
I ROS e le LOX La membrana cellulare e’ un sito attivo per l’induzione di meccansimi di difesa, infatti serve come ancora per I geni R che riconoscendo gli elicitori sono in grado di stimolare le risposte di difesa In molte combinazioni ospite-patogeno I cambiamenti che avvengono nella membrana svolgono un ruolo importante nella difesa Le più importanti risposte che avvengono a livello di membrana includono: Il rilascio di molecole importanti per la signal transduction sia nell’intorno della cellula che a livello sistemico Il rilascio e l’accumulo di ROS e di enzimi lipossigenasici L’attivazione di fenolossidasi In molte interazioni pianta-patogeno la prima risposta della pianta e’ la produzione di ROS. Queste specie reattive come l’anione superossido, l’idrossil radicale e il perossido d’idrogeno sono prodotti a partire dall’azione di una NADPH ossidasi di membrana dopo pochi secondi o minuti dal contatto con il patogeno o con un suo elicitore I ROS stimolano la perossidazione dei lipidi di membrana che se prodotti in alta concentrazione diventano tossici possono essere coinvolti nella morte HR La presenza dei ROS blocca comunque anche l’avanzamento del patogeno L’ossidazione delle membrane coinvolge l’attivazione di varie LOX che ossidano gli FFA rilasciati dalle membrane lipidiche Molti di questi LOOH vengono usati come precursori per la produzione di JA e di traumatina che agisce come trasduttore del segnale nelle reazioni di difesa

76
Le PR proteins Le proteine relative alla patogenesi (PR) sono proteine tossiche per il patogeno prodotte in grande quantità quando la pianta viene attaccata Si trovano sia a livello intra che intercellulare a livello di parete vegetale I diversi gruppi di PRP sono stati classificati in base alla funzione svolta, alle relazioni sierologiche, alle sequenze AA, al MW e ad altre proprieta’ Le PRP possono esssere o estremamente acide o basiche e quindi facilmente solubili e reattive Si conoscono almeno 14 classi di PRP, tra queste le PR1, (antioomiceti e antifungine), le PR2 (b-1,3-glucanasi), le PR3 (chitinasi), le PR4 (antifungine), le PR6 (inibitori di proteasi), le defensine, le thionine, I lisozimi, le proteine simili all’osmotina, le LOX, le proteine ricche in cys, le proteinasi, le chitosanasi e le perossidasi Ci sono molte isoforme di ciascuna PRP nelle varie piante I composti segnali che attivano la produzione e l’attivazione di PRP sono l’SA, l’ET, le xilanasi, la sistemina, il JA
 sono proteine tossiche per il patogeno prodotte in grande quantità quando la pianta viene attaccata. Si trovano sia a livello intra che intercellulare a livello di parete vegetale. I diversi gruppi di PRP sono stati classificati in base alla funzione svolta, alle relazioni sierologiche, alle sequenze AA, al MW e ad altre proprieta’ Le PRP possono esssere o estremamente acide o basiche e quindi facilmente solubili e reattive. Si conoscono almeno 14 classi di PRP, tra queste le PR1, (antioomiceti e antifungine), le PR2 (b-1,3-glucanasi), le PR3 (chitinasi), le PR4 (antifungine), le PR6 (inibitori di proteasi), le defensine, le thionine, I lisozimi, le proteine simili all’osmotina, le LOX, le proteine ricche in cys, le proteinasi, le chitosanasi e le perossidasi. Ci sono molte isoforme di ciascuna PRP nelle varie piante. I composti segnali che attivano la produzione e l’attivazione di PRP sono l’SA, l’ET, le xilanasi, la sistemina, il JA.")
77
Hypersensitivity- Specific Recognition
Some host cultivars can recognize certain pathogen isolates and trigger an HR whereas others can not. In this picture, the two inner cultivars are exhibiting an HR. Combinations of two or more HR-inducing recognition events can give rise to pathogen ‘races’ (more on this later). S HR HR S Localized cell death
. S HR HR S. Localized cell death.")
78
Specific recognition event triggers programmed cell death pathway
NO = Nitrous oxide

79
Timeline Review: Barley powdery mildew race A
is inoculated onto different host and nonhost plants: At what point on the timeline do the spores fail? 100 50 On susceptible barley On wheat Percentage of Conidia On resistant barley with HR- type resistance On tulip Hyphae Haustorium Germination Appressorium Peg extension

80
Induced biochemical defenses
Phytoalexins: antibiotics synthesized by and accumulated in plant host cells near those cells that have been attacked by pathogens Synthesized in healthy cells near an infection site – i.e., local induction Synthesis occurs all host cultivars but the rate (speed) of synthesis can vary among cultivars – an HR typically induces synthesis in nearby cells Most plants produce several phytoalexins (e.g., over 20 have been isolated from potato) Enzymes to detoxify phytoalexins have been observed in some fungal pathogens
 of synthesis can vary among cultivars – an HR typically induces synthesis in nearby cells. Most plants produce several phytoalexins. (e.g., over 20 have been isolated from potato) Enzymes to detoxify phytoalexins have been observed in some fungal pathogens.")
81
Phytoalexin molecules
Several hundred have been isolated and characterized from at least 15 plant families

82
Example of attack and counterattack
Fungal pathogen Host cell Phytoalexins are synthesized in response to pathogen attack Some pathogens counterattack producing enzymes that degrade the phytoalexin

83
Induced Resistance Develops after pre-inoculation of plants w/various biotic agents or after pre-treatment with various chemical or physical agents. It is non-specific since, regardless of the agent used as the inducer resistance is increased for a variety of pathogens. Local Acquired Resistance Systemic Acquired Resistance (SAR) Infection is localized, after several days, plant is resistant to attack in other areas. Usually resistance is expressed as small local lesions or complete resistance (no symptoms).
 Infection is localized, after several days, plant is resistant to attack in other areas. Usually resistance is expressed as small local lesions or complete resistance (no symptoms).")
84
Systemic Acquired Resistance
Inducer inoculation Local acquired resistance 3 days to months, then inoculate Systemic acquired resistance SAR- long-term resistance to a range of pathogens throughout plant caused by inoculation with inducer inoculum

85
Induced biochemical defenses
Systemic Acquired Resistance is the activation of defenses in distal, non-infected parts of the plant. The plant becomes more resistant to a secondary, challenge inoculation there 2 1 After a primary inoculation here

86
Systemic Acquired Resistance
SAR confers broad-based resistance to different pathogens. For example, primary inoculation with a fungal leaf spot pathogen reduces susceptibility of the host plant to other fungi as well as to bacterial and viral pathogens Salicylic acid (chemical related to aspirin) is part of signaling pathway involved in transmission of the defense response throughout the plant to produce SAR. This has lead to the development of synthetic chemicals that mimic the role of salicylic acid
 is part of signaling pathway involved in transmission of the defense response throughout the plant to produce SAR. This has lead to the development of synthetic chemicals that mimic the role of salicylic acid.")
87
Induction of Systemic Acquired Resistance
1- Production of H2O2 (plus antioxidants) Hydrogen peroxide has been associated secondary induction of SAR and direct toxic activity to invading pathogens 2 - Thickening of plant cell wall Production of phenolics (lignin, tannic acid) that strengthen walls and inhibit pathogen enzymes 3- Accumulation of pathogenesis related proteins “PR-proteins” enzymes that degrade fungal cell walls: chitinases, ß-1,3 Glucanases These enzymes accumulate in vacuole of plant cell. Upon attack, they directly degrade fungal cell walls. Indirectly, their action results in the release of fungal wall components that elicit additional defense reactions Sprayed inducer (activator) that mimics salicyclic acid
 Hydrogen peroxide has been associated. secondary induction of SAR and direct. toxic activity to invading pathogens. 2 - Thickening of plant cell wall. Production of phenolics (lignin, tannic acid) that strengthen walls and inhibit pathogen enzymes. 3- Accumulation of pathogenesis related proteins. PR-proteins enzymes that degrade fungal cell walls: chitinases, ß-1,3 Glucanases. These enzymes accumulate in vacuole of plant cell. Upon attack, they directly degrade fungal cell walls. Indirectly, their action results in the release of fungal wall components that elicit additional defense reactions. Sprayed inducer. (activator) that mimics. salicyclic acid.")
88
Plants can have more than one type of acquired resistance
Strains of bacteria living on roots Pathogens or synthetic chemicals

89
Plant Defense: Systemic Acquired Resistance
Following HR, increased resistance throughout the plant Due to increased expression of pathogenesis-related genes Long-lasting effect Salicylic acid is required for induction

90
Signal transduction nella SAR
Le difese indotte sono regolate da una rete di pathway di segnalazione interconnesse in cui I componenti primari sono l’acido salicilico (SA), l’acido jasmonico (JA), l’etilene (ET) e l’ossido nitrico (NO) La pianta reagisce all’attacco di un patogeno aumentando la produzione di queste sostanze mentre un distinto set di geni relativi alla difesa attiva nella resistenza gene per gene tenta di bloccare l’infezione L’acido salicilico reagisce con diverse proteine tra cui l’ascorbato perossidasi e la catalasi (enzimi scavengers dell’H2O2) e con una proteina plastidiale con attivita’ antiossidante La principale via di difesa stimolata dal SA e’ quella legata all’attivazione delle PR (pathogenesis related) proteins. Alcuni dei geni “accesi” dal SA attivano altri geni connessi con la sintesi di JA e di ET L’NO invece induce la sintesi di PR1 e dell’enzima PAL che e’ alla base della formazione di SA L’SA di solito non e’ richiesto nelle prime difese stimolate dai geni R ma piuttosto per la propagazione ed il mantenimento della risposta sistemica secondaria (SAR) Tutti gli ormoni (SA, JA, ET) si influenzano reciprocamente in modo positivo o negativo durante le risposte difensive generando un cosiddetto “crosstalk” Il crosstalk fornisce un potenziale regolativo per attivare diversi meccansimi di resistenza in varie combinazioni e svolge un ruolo nel determinare quale via difensiva attivare
, l’acido jasmonico (JA), l’etilene (ET) e l’ossido nitrico (NO) La pianta reagisce all’attacco di un patogeno aumentando la produzione di queste sostanze mentre un distinto set di geni relativi alla difesa attiva nella resistenza gene per gene tenta di bloccare l’infezione. L’acido salicilico reagisce con diverse proteine tra cui l’ascorbato perossidasi e la catalasi (enzimi scavengers dell’H2O2) e con una proteina plastidiale con attivita’ antiossidante. La principale via di difesa stimolata dal SA e’ quella legata all’attivazione delle PR (pathogenesis related) proteins. Alcuni dei geni accesi dal SA attivano altri geni connessi con la sintesi di JA e di ET. L’NO invece induce la sintesi di PR1 e dell’enzima PAL che e’ alla base della formazione di SA. L’SA di solito non e’ richiesto nelle prime difese stimolate dai geni R ma piuttosto per la propagazione ed il mantenimento della risposta sistemica secondaria (SAR) Tutti gli ormoni (SA, JA, ET) si influenzano reciprocamente in modo positivo o negativo durante le risposte difensive generando un cosiddetto crosstalk Il crosstalk fornisce un potenziale regolativo per attivare diversi meccansimi di resistenza in varie combinazioni e svolge un ruolo nel determinare quale via difensiva attivare.")
91
Salicylic Acid Colorless, crystalline organic carboxylic acid
Functions as a plant hormone All cells capable of synthesizing Released by cells with in favorable environmental conditions; allow survival at current metabolic level Acts by rapid local increases then rapid decreases in levels After survival threat has passed, returns to normal functioning Increases cell metabolism rate to take advantage of more advantageous nutrient and environmental conditions Sustained level of SA may occur if a cell reaches peak metabolic levels and may signal that resources can be turned to growth

92
Salicylic Acid (SA) COOH OH Accumulates in both local and systemic tissues (not the systemic signal) Removal of SA (as in nahG plants) prevents induction of SAR Analogs: INA or BTH
 prevents induction of SAR. Analogs: INA or BTH.")
93
Jasmonic Acid Methyl jasmonate catabolized from jasmonic acid, more volatile; may act as airborne signal messenger to neighboring undamaged plants Herbivore oral secretions contact damaged plant tissue & prompt release of volatile organic molecules that attract herbivore predators; methyl jasmonate may be involved

94
Plant Defense: Jasmonic Acid Pathway

95
Coronatine Phytotoxin produced by some plant-pathogenic bacteria
Mimics the action of methyl jasmonate (MeJA) in plants CFA - Polyketide coronafacic acid CMA - Cyclized amino acid; coronamic acid
 in plants. CFA - Polyketide coronafacic acid. CMA - Cyclized amino acid; coronamic acid.")
96
Previous study: Investigate SA/JA cross-talk
Conclusion: Not a simple mechanism where resistance to bacteria = susceptibility to caterpillar

97
Three ways plant hormones interact:
Mode I - Response Level -Hormones interact by regulating a common set of genes ie. Auxin response genes regulated by ethylene and gibberellins Many genes regulated by ethylene and JA signaling pathways Mode II - Signal Transduction Level -One component functions in both signaling pathways ie. JA/COR signaling affects plant response to both pathogen and herbivore Mode III - Biosynthesis Level -Production of one hormone alters in response to second hormone ie. SA negatively regulates JA

98
MeJA, CMA, CFA Methyl jasmonate (MeJA) is an inducer of the JA pathway
- COR is a MeJA mimic CMA and CFA are compounds that come together to form COR

99
MODEL Virulent P. syringae Avirulent P. syringae Disease HR JA/COR SA
SIS Disease Avirulent P. syringae HR SA SAR No disease T. ni susceptibility T. ni resistance

100
Mutants affecting SA synthesis
Elevated SA accumulation dnd1 (defense, no death 1): increased SA, but reduced HR, DND1 gene encodes cyclic-nucleotide-gated ion channel mpk4: constitutive SA accumulation edr1 (enhanced disease resistance 1): defective MAPKKK
: increased SA, but reduced HR, DND1 gene encodes cyclic-nucleotide-gated ion channel. mpk4: constitutive SA accumulation. edr1 (enhanced disease resistance 1): defective MAPKKK.")
101
Mutants affecting SA synthesis
reduced SA accumulation eds1 (enhanced disease susceptibility 1): lipase homolog pad4 (phytoalexin deficient 4): another lipase homolog sid1 and sid2 (salicylic acid induction-deficient): defects in chorismate pathway
: lipase homolog. pad4 (phytoalexin deficient 4): another lipase homolog. sid1 and sid2 (salicylic acid induction-deficient): defects in chorismate pathway.")
102
NPR1: non-expresser of PR genes
Also known as NIM1 or SAI1 Positive regulator of SAR Downstream of SA, upstream of PR genes npr1 mutants are susceptible to various pathogens Overexpression of NPR1 generates broad-spectrum resistance Unique, but similar to Iκ-B (negative regulator of immunity in animals)
")
103
NPR1 overexpression

104
Pathogen-Related (PR) Genes
Antimicrobial properties Many identified Categorized according to activity Examples PR-2 : beta-1,3-glucanase PR-3 : chitinase PR-12: defensin

105
SAR Avr R gene SA NPR1 PR-1 PR-2 PR-5 SAR

106
Structural features of NPR1
nim 1-2 npr 1-1 NLS S S BTB ARD 593 amino acids, 67 kD Two protein-protein interaction domains: BTB/POZ and Ankyrin repeats Contains NLS Multiple phosphorylation sites No DNA binding domain

107
TGA Factors Found to interact with NPR1 through yeast-two hybrid
bZIP transcription factors Six members in Arabidopsis (TGA1-6) Might be redundant Bind to as-1 element
 Might be redundant. Bind to as-1 element.")
108
NPR1-TGA2 interaction Direct visualisation

109
SAR Avr R gene SA TGA2 NPR1 PR-1 PR-2 PR-5 SAR

110
Current model Figure 6

111
SAR Avr R gene SA TGA2 NPR1 TGA5 PR-1 PR-2 PR-5 SAR SAR independent
resistance

112
Why are secretion-related proteins induced?
Hypothesis: A coordinated up-regulation of protein secretion machinery is required for the proper folding, modification, and transport of PR proteins.

113
Genetic Evidence Mutant analysis: Calnexin mutant PDI mutant
BiP2 mutant DAD1 mutant SEC61 mutant No visible phenotype Inability to transport PR proteins when induced by SA analog (S-methylester)
")
114
Conclusions NPR1 not only induces PR genes but also secretion-related genes These secretion-related genes are required for the efficient transport of PR proteins and SAR induction NPR1 interacts with a cis-element (TL1) to induce secretion-related genes separately from the PR gene element (as-1)
 to induce secretion-related genes separately from the PR gene element (as-1)")
115
Similarity with animals
1/ Resistance/avirulence gene interaction is analogous to animal antibodies- involves protein-protein binding is highly specific 2/ Oxidative burst is analogous to neutrophil action 3/ Programmed cell death is common to both plants and animals 4/ Systemic acquired resistance is like immunity? Convergent evolution!!!!

116
…per riassumere!

117
Powdery mildew disease
I funghi che causano questa malattia attaccano specie di piante appartenenti a piu di 1600 generi Le difese contro questi funghi sono ottenute grazie a dei geni R. Tra questi e’ stato studiato il gene Mla di orzo che conferisce la resistenza a 32 razze di Blumeria graminis f.sp. Hordei Mla e’ una proteina recettore con domini CC-NB-LRR capaci di riconoscere almeno 3 fattori AVR del fungo I geni Rar1 e SGT1 sono sotto il controllo di Mla e sono fondamentali per lo scatenamento delle risposte di difesa Rar1 e’ sotto il controllo di un altro fattore, RPW8, proteina che presenta dei domini transmembrana ed e’ coinvolta nella resistenza dell’orzo a diverse powdery mildews. RPW8 induce, anche attraverso il coinvolgimento di Rar1 e di acido salicilico, l’accumulo di ROS, la morte ipersensibile e l’attivazione di PR-1 proteins Si pensa che RPW8 agisca come fattore di compatibilita’ che rende possibile il trasporto di molti fattori di patogenicita’ all’interno dell’ospite. In tal modo RPW8 consente ai recettori intracellulari di riconoscere tali fattori e di attivare le risposte difensive Una caratteristica della resistenza dell’orzo mediata da Mlo a Blumeria e’ quello di essere capace di bloccare l’infezione anche dopo la penetrazione senza necessita’ della morte ipersensibile Il tentativo di penetrazione del fungo porta a rimodellare la parete cellulare intorno all’ifa di penetrazione e all’appresorio In tale sito si accumula no ROS e composti fenolici antimicrobici

118
Magnaporthe grisea Il conidio del patogeno germina rapidamente e tenta di penetrare la superficie fogliare, in 4 ore l’apice del tubo si allarga, la parete si inspessisce con l’apposizione di melanina Numerosi geni sono attivi durante questo processo, una idrofobina (MPG1) che aiuta l’appressorio a riconoscere le superfici idrofobiche della foglia; il gene Pth11 che sembra essere coinvolto nella signal transduction legata alla formazione dell’appressorio cosi come magB che codifica per una proteina Ga e la MAPK PMK1. PMK1 controlla in particolare la germinazione conidica attraverso la degradazione di glicogeno e lipidi che proprio grazie al segnale trasdotto da questa MAPK, vengono mobilizzati verso l’appresorio in formazione
 che aiuta l’appressorio a riconoscere le superfici idrofobiche della foglia; il gene Pth11 che sembra essere coinvolto nella signal transduction legata alla formazione dell’appressorio cosi come magB che codifica per una proteina Ga e la MAPK PMK1. PMK1 controlla in particolare la germinazione conidica attraverso la degradazione di glicogeno e lipidi che proprio grazie al segnale trasdotto da questa MAPK, vengono mobilizzati verso l’appresorio in formazione.")
119
Fusarium Fusarium oysporum consiste di piu di 120 formae specialis a seconda dell’ospite infettato. Ogni f.sp. e’ ulteriormente suddivisa in razze fisiologiche ognuna con un caratteristico pattern di virulenza su diverse varieta dell’ospite Il cambiamento di pH del suolo induce l’attivazione di un fattore di trascrizione che attiva geni specificamente attivi in condizioni di alcalinita’ ementre inibisce I geni attivi a pH acidi, alterando la crescita e le capacita di patogenesi I primi segnali di riconoscimento fungo-pianta includono il fattore di trascrizione CTF1b. Questo media l’attivazione di cut2 che codifica per una cutinasi che una volta attiva attacca la cutina della pianta rilasciando nell’ambiente monomeri di cutina. Questi stimolano l’attivazione di CTF1a che attiva a sua volta la cutinasi principale, cut1. L’adesione e la penetrazione nella radice sono sotto il controllo di una MAPK. Questa controlla l’attivazione sequenziale di molti CWDE che vengono poi rilasciati anche all’interno dei vasi infetti Una volta dentro la pianta il fungo entra in contatto con sostanze anttimicrobiche preesistenti come fitoanticipine e fitoalessine. In particolare il pomodoro produce la saponina a-tomatina, la patata a-calconina e a-solanina. Il fungo produce degli enzimi in grado di tagliare queste molecole in composti non tossici Il fungo e’ anche in grado di degradare le fitoalessine attraverso degli enzimi demetilasici Pda2 e PdaL e H Le varie specie di Fusarium sono anche in grado di produrre molte sostanze tossiche come ad esempio le fumonisine. Queste sono in grado di interferire con la sintesi delle cere superficiali e inducono apoptosi.

120
Targets for Improvement
For example, rice Targets for improvement in rice production fall into three categories Biotic constraints- (pests and diseases) Weeds, Fungi (e.g. Blast), Bacteria (e.g. Blight), Viruses (e.g. Rice yellow mottle virus), Insects (e.g. Brown plant hopper), Nematodes (e.g. Cyst-knot nematode) Abiotic constraints (adverse physical environment) Drought, Nutrient availability, Salinity Cold, Flooding Yield and quality Plant morphology, Photosynthetic efficiency, Nitrogen fixation, Carbon partitioning, Aroma
 Weeds, Fungi (e.g. Blast), Bacteria (e.g. Blight), Viruses (e.g. Rice yellow mottle virus), Insects (e.g. Brown plant hopper), Nematodes (e.g. Cyst-knot nematode) Abiotic constraints (adverse physical environment) Drought, Nutrient availability, Salinity Cold, Flooding. Yield and quality. Plant morphology, Photosynthetic efficiency, Nitrogen fixation, Carbon partitioning, Aroma.")
121
Marker Assisted Selection in Disease Resistance
Resistance genes can be selected for by screening with the disease. So, conventional breeding can produce resistant varieties. But, resistance genes break-down. The disease organism mutates to overcome them (in 2-3 years). If there were several resistance genes, the disease organism would take very much longer to overcome all resistance genes (in fact it is virtually impossible). But, you can’t select for say 3 resistance genes conventionally- you can’t tell the difference between 1 gene and 2 or 3 by phenotype. But if you select for markers linked to the resistance genes, you can introduce multiple resistance genes.
. If there were several resistance genes, the disease organism would take very much longer to overcome all resistance genes (in fact it is virtually impossible). But, you can’t select for say 3 resistance genes conventionally- you can’t tell the difference between 1 gene and 2 or 3 by phenotype. But if you select for markers linked to the resistance genes, you can introduce multiple resistance genes.")
122
Elite variety with multiple resistance genes
Marker Assisted Selection in Disease Resistance Selectable markers Elite variety Donor Donor Donor 3 Multiple crosses followed by backcrossing with selection for markers at every stage Elite variety with multiple resistance genes

123
Vantaggi e svantaggi nel breeding della resistenza orizzontale e verticale
La resistenza può essere ottenuta incorporando uno, pochi o molti geni di resistenza in una varietà. Alcuni di questi geni possono controllare alcuni importanti steps nello sviluppo della malattia e quindi svolgono un ruolo importante nella resistenza La resistenza verticale e’ utile contro specifici patogeni o specifiche razze di patogeno E’ maggiormente efficace quando E’ incorporata in specie annuali, facile da migliorare come I cereali E’ diretta contro patogeni che non si riproducono e diffondono rapidamente come Fusarium o contro patogeni che non mutano frequentemente come Puccinia Consistono di geni R “forti” che conferiscono una resistenza completa e a lungo termine La popolazione ospite non consiste di una varieta uniforme geneticamente Se una sola di queste condizioni non si verifica la resistenza diventa di breve durata e compaiono nuove razze di patogeno che bypassano le difese dell’ospite La resistenza orizzontale Conferisce una resistenza parziale ma di più lunga durata Tale resistenza coinvolge più processi fisiologici dell’ospite che agiscono come meccansimi di difesa e che sono degli ostacoli impossibili da superare per il patogeno Le piante con la resistenza orizzontale rimangono resistenti più a lungo ma con minore intensita’

124
Multilinee e piramidalizzazione
Le varieta’ con la resistenza verticale sono spesso attaccate all’improvviso da nuove razze di patogeno e questo può portare a delle vere e proprie epidemie Per evitare questo svantaggio, cioe’ l’inefficienza della resistenza affidata a pochi geni, vengono oggi usate le multilinee attraverso la piramidalizzazione Le multilinee sono una miscela di varieta’ individuali (linee e cultivar) che sono agronomicamente simili ma che differiscono nei loro geni di resistenza La piramidalizzazione consiste nell’uso di varietà che sono derivate dall’incrocio di molte varietà che contengono geni R diversi e poi selezionando da queste solo quelle che contengono una miscela di tutti I geni R Le multilinee e la piramidalizzazione sono stati sviluppate come metodiche soprattutto nella lotta contro le ruggini dei cereali, ma il loro uso sta decisamente aumentando anche in altri sistemi pianta-patogeno
 che sono agronomicamente simili ma che differiscono nei loro geni di resistenza. La piramidalizzazione consiste nell’uso di varietà che sono derivate dall’incrocio di molte varietà che contengono geni R diversi e poi selezionando da queste solo quelle che contengono una miscela di tutti I geni R. Le multilinee e la piramidalizzazione sono stati sviluppate come metodiche soprattutto nella lotta contro le ruggini dei cereali, ma il loro uso sta decisamente aumentando anche in altri sistemi pianta-patogeno.")
125
Genetic transformations
Agrobacterium transformations Direct transfer methods for transformation Transformation cassettes From transformed cells to plants The use of transformed plants in research Mutants Transposon Transposon and T-DNA tagging

126
Genetic Engineering of Plants-
Agrobacterium transformation- The bacteria Agrobacterium tumefaciens causes galls or tumors on plants Ti Plasmid (tumor inducing) Genomic DNA T-DNA (transfer) Foreign DNA T-DNA (transfer) Restrict and ligate together Re-introduce recombinant DNA
 Genomic DNA. T-DNA. (transfer) Foreign DNA. T-DNA. (transfer) Restrict and ligate together. Re-introduce recombinant DNA.")
127
Agrobacterium transformation 2
Whole T-DNA transferred randomly into plant chromosome Infect plant with recombinant agrobacterium Grow up transformed plants from single cells

128
“GENETIC ENGINEERING” without AGROBACTERIUM
All involve getting DNA directly across the plasma membrane Shock of protoplasts Micro-injection Biolistics

129
Allows transgenic cells to be selected from
Transformation constructs or cassettes Genes of interest Promoter Selectable (marker) gene Gene of interest T-DNA T-DNA Promoter e.g. Cauliflower Mosaic Virus 35S RNA gene promoter (CAM 35S) Selectable marker-gene e.g. antibiotic resistance or herbicide resistance Allows transgenic cells to be selected from non-transgenic
 gene. Gene of interest. T-DNA. T-DNA. Promoter. e.g. Cauliflower. Mosaic Virus 35S RNA gene promoter. (CAM 35S) Selectable marker-gene. e.g. antibiotic resistance or herbicide resistance. Allows transgenic cells to be selected from. non-transgenic.")
130
From transformed cells to plants
Plant cells are grown as a callus of undifferentiated cells on agar plates transformation After transformation, cells grown on selective media (e.g. containing antibiotic) Untransformed cells die selection Transfer to tube with hormones Cells containing transgenes grow into plantlets
 Untransformed cells die. selection. Transfer to tube with hormones. Cells containing transgenes grow into plantlets.")
131
Transposons and T-DNA tagging
Transposons have only been found in a few plants (e.g. Maize, Antirrhium). But, they can be introduced by transformation. The Ac transposon has been introduced to tobacco, Arabidopsis, potato, tomato, bean and rice. Mutations using transposons or T-DNA (both of which insert randomly into nuclear DNA) are produced by transformation methods described earlier. Large numbers of plants are screened for an observable phenotype (e.g. lack of response to light). Screen Identify mutated gene
. But, they can be introduced by transformation. The Ac transposon has been introduced to tobacco, Arabidopsis, potato, tomato, bean and rice. Mutations using transposons or T-DNA (both of which insert randomly into nuclear DNA) are produced by transformation methods described earlier. Large numbers of plants are screened for an observable phenotype (e.g. lack of response to light). Screen. Identify mutated. gene.")
132
Transposons and T-DNA tagging
The gene into which the insert has occurred can be recovered by PCR Insertion (Transpososn or T-DNA) Mutated ORF Restrict Ligate PCR amplify using primers homologous to and facing out of insert
 Mutated ORF. Restrict. Ligate. PCR amplify using primers homologous to and facing out of insert.")
133
In Arabidopsis, you can get a mutant of any gene you want
Transposons and T-DNA tagging In Arabidopsis and maize, large collections of Tansposon and T-DNA tagged mutants have been developed over the last few years. In Arabidopsis, you can get a mutant of any gene you want They represent a fantastic tool for the discovery of plant genes and gene function.

134
Da cellule singole si possono far crescere piante intere
Se una pianta viene incisa sotto la ferita si ha una proliferazione cellulare detta callo Se un frammento di callo giovane è rimosso e posto in un terreno di coltura contenente i nutrienti e gli ormoni appropriati In seguito il callo si differenzierà in germogli e radici e alla fine sarà prodotta un intera pianta con fiori Il differenziamento del callo dipende dal rapporto tra 2 ormoni, le citochinine e le auxine. Quando il rapporto aux/cit è alto si sviluppano radici…

135
Rigenerazione di piante da protoplasti

136
Trasformazione di cellule vegetali
Le cellule vegetali mature sono circondate da una parete cellulare costituita da cellulosa, emicellulosa, pectina e lignina e quindi non sono molto idonee alla captazione del DNA La parete cellulare può essere rimossa mediante degli enzimi litici I protoplasti, cellule vegetali senza parete, possono acquisire macromolecole come il DNA e sono capaci di rigenerare piante intere

137
Esempio di trasformazione
Da foglie di tabacco vengono prelevati dei dischi fogliari Le cellule al margine del disco cominciano a rigenerare e se vengono messe in presenza di A. tumefaciens possono essere facilmente trasformate Le cellule che recano il plasmide vengono selezionate coltivandole nel terreno stimolatore di germogli e in presenza di un antibiotico appropriato quale la kanamicina (NPTII) e il cefotaxime che serve per bloccare la proliferazione del A. tumefaciens
 e il cefotaxime che serve per bloccare la proliferazione del A. tumefaciens.")
138
Rigenerazione di dischi fogliari

139
Altri metodi per inserire DNA in pianta
Virus I virus si sono adattati nell’evoluzione a trasferire il proprio genoma negli organismi ospite Possono essere usati sia su dicotiledoni che su monocotiledoni Contengono 2 filamenti di DNA a singola elica il DNA A (contiene geni per la costruzione del capside) e il DNA B (contiene i geni per la virulenza)
 e il DNA B (contiene i geni per la virulenza)")
140
Infezione sistemica

141
Altri metodi per inserire DNA in pianta
Elletroporazione Un’alta concentrazione di DNA plasmidico contenente un gene clonato viene aggiunta a una sospensione di protoplasti e il miscuglio viene sottoposto a shock con campo elettrico che va da 200 a 600 V/cm In seguito a elettroporazione i protoplasti vengono fatti crescere in colture tissutali per una o due settimane prima di iniziare la selezione dei cloni positivi

142
Altri metodi per inserire DNA in pianta
Gene Gun Sferette di tungsteno rivestite di DNA vengono sparate direttamente nelle cellule vegetali. I bersagli finora scelti includono Sospensioni di cellule embrionali Foglie intattte Semi Il bombardamento può produrre organuli transgenici Cloroplasti mitocondri

143
Gene Gun-1

144
Gene Gun-2

Presentazioni simili















 | doi: /213399a0>")



