Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Corso di biochimica 1 Prof. Giuseppina Pitari
L’emoglobina II Corso di biochimica 1 Prof. Giuseppina Pitari

2
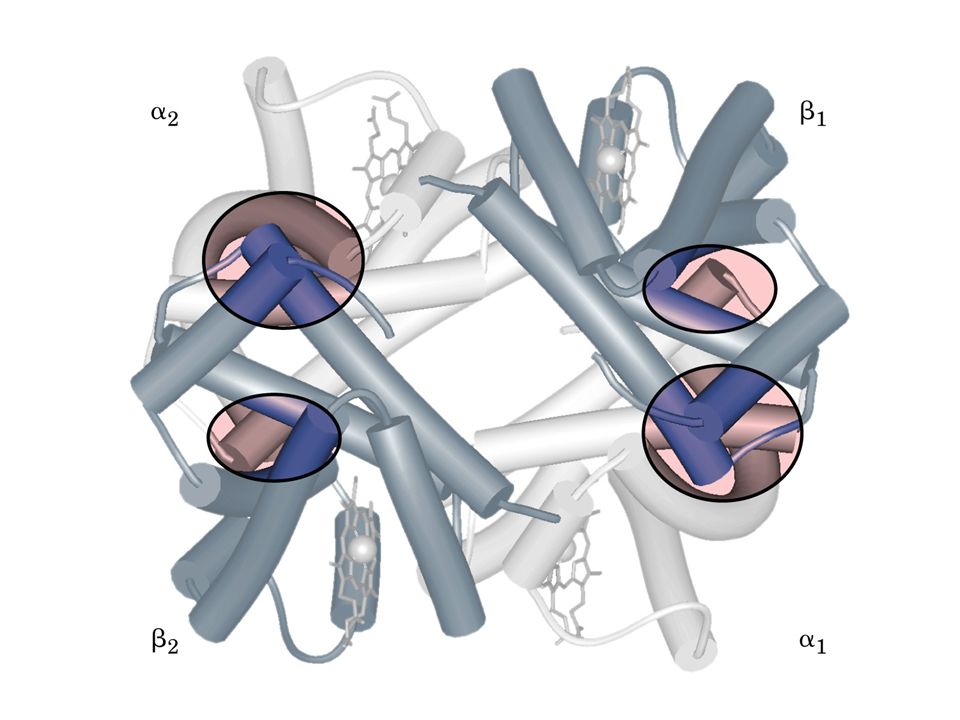
Le catene polipeptidiche sono organizzate in modo da avere molteplici interazioni tra subunità di tipo diverso (legami idrogeno, ponti salini, interazioni idrofobiche): 1. L’interfaccia a1-b1 (o a2-b2): comprende ~ 35 residui (interfaccia rigida o statica) Queste interazioni sono , in complesso, forti tanto che a basse concentrazioni di urea il tetrametro si disassembla, ma i dimeri rimangono intatti
: comprende ~ 35 residui (interfaccia rigida o statica) Queste interazioni sono , in complesso, forti tanto che a basse concentrazioni di urea il tetrametro si disassembla, ma i dimeri rimangono intatti.")
3
UREA

4
Le catene polipeptidiche son organizzate in modo da avere molteplici interazioni tra subunità di tipo diverso (legami idrogeno, ponti salini, interazioni idrofobiche): 2. L’interfaccia a1-b2 (o a2-b1): comprende ~ 19 residui (interfaccia dinamica o mobile) Questi residui fanno parte dell’elica C e l’angolo FG
: comprende ~ 19 residui (interfaccia dinamica o mobile) Questi residui fanno parte dell’elica C e l’angolo FG.")
5
I contatti a1-a2 o b1-b2 sono pochi e di carattere polare
I contatti a1-a2 o b1-b2 sono pochi e di carattere polare. Queste subunità si guardano l’un l’altra attraverso un canale pieno di solvente del diametro di ~ 20 Å coassiale con l’asse di simmetria della molecola. a1 b2 b1 a2

7
Quando l’emoglobina lega l’ossigeno i contatti a1b1 cambiano poco mentre un grande cambiamento si ha nei contatti a1b2 (parecchie coppie ioniche rotte).
.")
8
L’analisi della struttura ai raggi X ha rivelato due conformazioni per la emoglobina: lo stato R e quello T. Anche se l’ossigeno si lega ad ambedue gli stati si ha una considerevole più alta affinità per lo stato R. Quando l’ossigeno è assente sperimentalmente , lo stato T è più stabile e così lo stato T è lo stato predominante per la desossiemoglobina. T e R stanno per “teso” e “rilassato”, questo perché lo stato T è stabilizzato da un più grande numero di coppie ioniche nelle interfacce a1b2, a2b1.

9
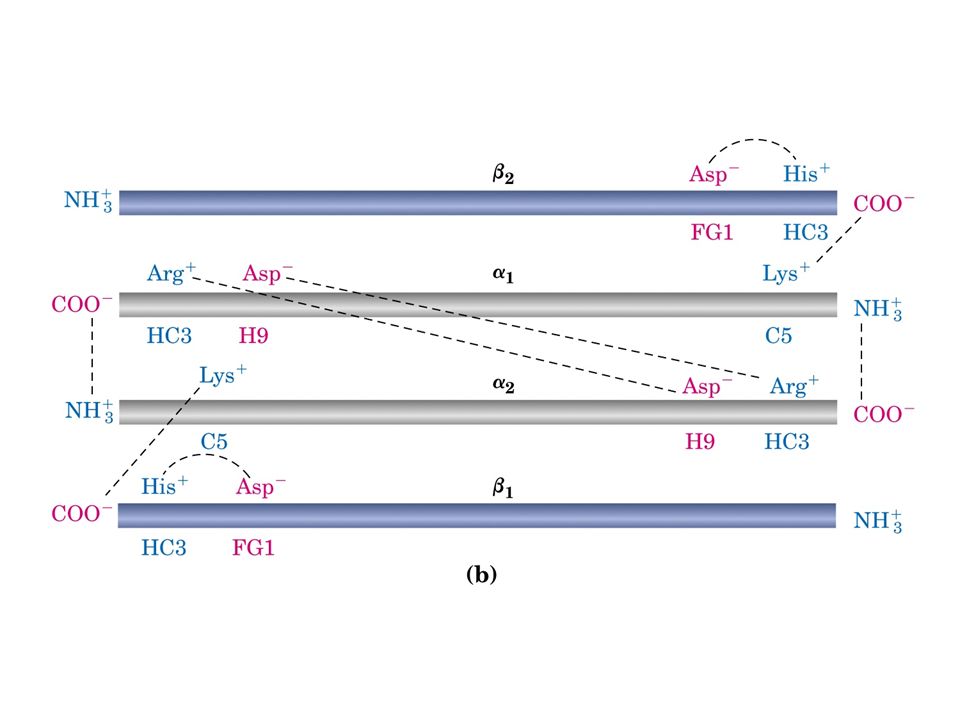
Desossiemoglobina: Coppie ioniche tra His HC3 e Asp FG1 sulla subunità beta e tra la lisina C5 della subunità alfa e il gruppo a –carbossilico dell’istidina HC3 della subunità beta

11
IL LEGAME CON L’OSSIGENO
L’ossigeno, legandosi all’emoglobina nello stato T, provoca un cambiamento conformazionale e la porta nello stato R. Quando l’intera struttura subisce questo cambiamento, la struttura delle singole subunità cambia poco ma le coppie di subunità ab scivolano e ruotano, stringendo la tasca tra le subunità b. In questo processo alcune delle coppie ioniche che stabilizzano la forma T vengono rotte e se ne formano delle nuove.

12
La transizione dallo stato T allo stato R sposta le coppie delle subunità: l’stidina HC3 ruota nello stato R verso il centro della molecola dove non c’è possibilità di formare coppie ioniche.

13
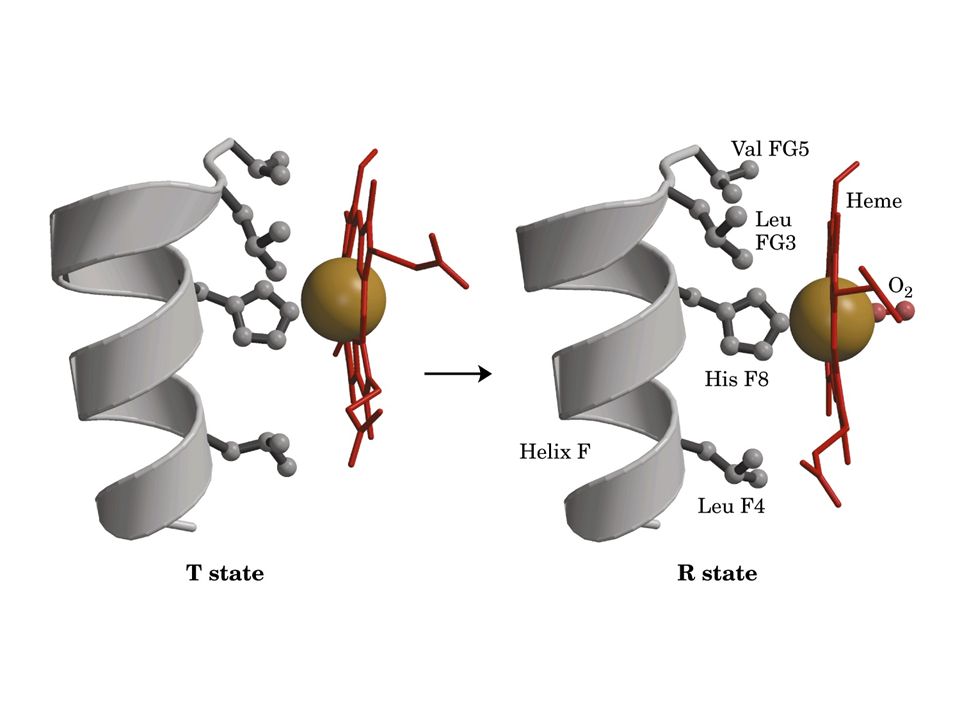
L’ossigenazione determina una grande modificazione della struttura quaternaria, ovvero nell’interazione fra residui interfacciali Lo stato desossigenato è detto T (tense): bassa affinità per l’O2; il Fe+2 è spiazzato 0.6 Å in direzione prossimale. L’eme ha una forma a cupola. Lo stato ossigenato è detto R (relaxed): alta affinità per l’O2; il Fe+2 giace nel piano dell’eme (che assume una forma planare)
: bassa affinità per l’O2; il Fe+2 è spiazzato 0.6 Å in direzione prossimale. L’eme ha una forma a cupola. Lo stato ossigenato è detto R (relaxed): alta affinità per l’O2; il Fe+2 giace nel piano dell’eme (che assume una forma planare)")
15
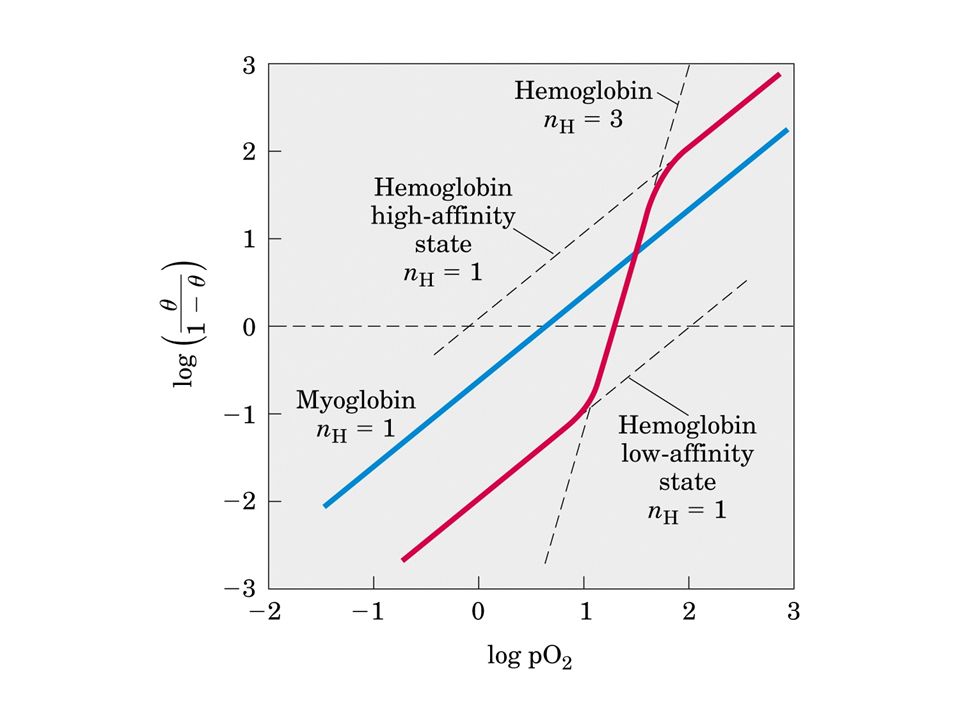
L’emoglobina lega l’ossigeno in maniera cooperativa
Una curva sigmoide può essere vista come una curva ibrida che riflette una transizione tra uno stato a bassa affinità ad uno ad alta affinità

16
L’emoglobina lega l’ossigeno in maniera cooperativa
L’emoglobina deve legare l’ossigeno efficientemente nei polmoni dove la pO2 è circa 13,3 kPa (1 mm Hg=133,322 Pa) e rilasciare l’ossigeno al livello dei tessuti dove la pressione parziale di ossigeno è 4 kPa. Per risolvere questo problema l’emoglobina subisce un cambiamento conformazionale da uno stato a bassa affinità ad uno ad alta affinità. Come risultato la curva di ossigenazione è sigmoide il che significa che ogni ligando (l’ossigeno) si lega alla proteina in maniera non indipendente: la prima molecola di ossigeno si lega con bassa affinità all’emoglobina perchè si lega allo stato T. Il suo legame, però, porta ad un cambiamento conformazionale che viene comunicato alle subunità adiacenti rendendo più facile alla seconda molecola di ossigeno di legarsi. In effetti la transizione T—R avviene più velocemente nella seconda subunità una volta che la prima molecola di ossigeno si è legata. Il quarto ossigeno si lega all’eme in una subunità che è già nello stato Re si lega, quindi, con una più alta affinità.
 e rilasciare l’ossigeno al livello dei tessuti dove la pressione parziale di ossigeno è 4 kPa. Per risolvere questo problema l’emoglobina subisce un cambiamento conformazionale da uno stato a bassa affinità ad uno ad alta affinità. Come risultato la curva di ossigenazione è sigmoide il che significa che ogni ligando (l’ossigeno) si lega alla proteina in maniera non indipendente: la prima molecola di ossigeno si lega con bassa affinità all’emoglobina perchè si lega allo stato T. Il suo legame, però, porta ad un cambiamento conformazionale che viene comunicato alle subunità adiacenti rendendo più facile alla seconda molecola di ossigeno di legarsi. In effetti la transizione T—R avviene più velocemente nella seconda subunità una volta che la prima molecola di ossigeno si è legata. Il quarto ossigeno si lega all’eme in una subunità che è già nello stato Re si lega, quindi, con una più alta affinità.")
17
PROTEINE ALLOSTERICHE
Le proteine allosteriche sono quelle nelle quali il legame di un ligando ad un sito influenza le proprietà di legame di un altro sito sulla stessa proteina. Sono proteine che posseggono “altre” conformazioni indotte dal ligandi che possono essere chiamati “modulatori” o effettori allosterici.: questi modulatori possono essere sia inibitori che attivatori. Quando il ligando e il modulatore sono la stessa molecola l’interazione è chiamata omotropica, altrimenti eterotropica. Alcune proteine hanno più di un modulatore e quindi possono dar luogo a interazioni omotropiche ed eterotropiche.

18
L’EQUAZIONE DI HILL Il legame cooperativo dell’ossigeno fu studiato da Archibald Hill nel 1910

19
Archibald Vivian Hill 1886-1977
Il primo tentativo volto ad analizzare la curva si saturazione sigmoide dell’O2 nell’Hb fu formulato da Hill (Premio Nobel per la Medicina, 1922). Egli ipotizzò che l’Hb potesse legare n molecole di O2 in una sola tappa (cooperatività infinita): Archibald Vivian Hill
. Egli ipotizzò che l’Hb potesse legare n molecole di O2 in una sola tappa (cooperatività infinita): Archibald Vivian Hill")
20
La trasformazione secondo Hill produce
La quantità n, cioè il coefficiente di Hill, è una misura della cooperatività del sistema. Quando: n = 1: il sistema è non cooperativo. Il binding del legante è semplice (comportamento iperbolico) n > 1: il binding presenta cooperatività positiva (curva sigmoide) n < 1: il binding presenta cooperatività negativa (curva a scalino)
 n > 1: il binding presenta cooperatività positiva (curva sigmoide) n < 1: il binding presenta cooperatività negativa (curva a scalino)")
21
Hb n=2.8 Mb n=1 asintoto superiore p50 della 4A O2 legata
asintoto inferiore 21

23
Cooperatività del legame dell’O2: basi strutturali
La cooperatività positiva (n>1) del legame dell’ossigeno nell’Hb deriva dalle modificazioni strutturali che il binding di una molecola di O2 ad un gruppo eme può indurre sull’affinità per l’O2 di un altro gruppo eme.
 del legame dell’ossigeno nell’Hb deriva dalle modificazioni strutturali che il binding di una molecola di O2 ad un gruppo eme può indurre sull’affinità per l’O2 di un altro gruppo eme.")
24
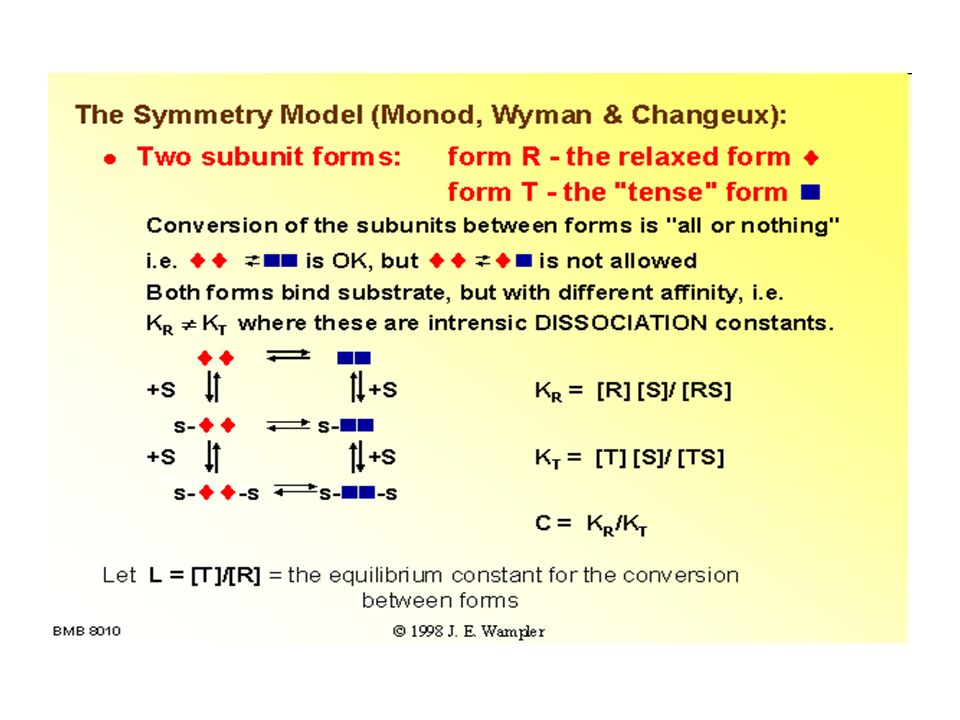
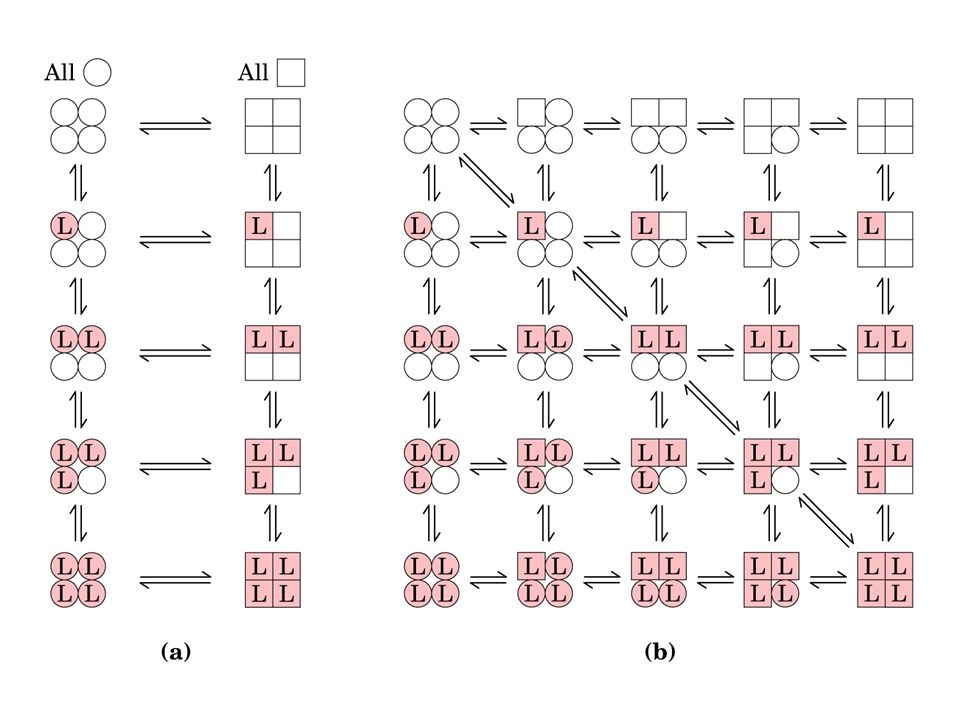
COME AVVIENE LA TRANSIZIONE T----R
Due modelli: MODELLO MWC: Monod, Wyman, Changeux 1965 Questo modello assume che le subunità di una proteina con cooperatività di legame siano funzionalmente identiche e che tutte possono esistere in due conformazioni e che ciascuna subunità va da una transizione all’altra simultaneamente e che le due conformazioni sono in equilibrio. Il ligando può legarsi ad ambedue le conformazioni, ma si lega a ciascuna con differente affinità. Il legame successivo delle molecole di ligando alla conformazione a bassa affinità fa avvenire la transizione verso la conformazione ad alta affinità

26
Secondo Modello Koshland 1966
Il legame del ligando può indurre un cambiamento di conformazione in una subunità. Questo produce cambiamenti simili nelle subunità adiacenti Esistono, quindi, un gran numero di conformazioni I due modelli non si escludono a vicenda

28
L’emoglobina trasporta anche anidride carbonica e H+
L’emoglobina trasporta via dai tessuti i prodotti del catabolismo: anidride carbonica e H+. L’anidride carbonica è trasformata velocemente in ione bicarbonato CO2 + H2O ==== H+ + HCO3- Quindi l’idratazione dell’anidride carbonica porta allo sviluppo di H+ con conseguente abbassamento di pH nei tessuti

29
L’emoglobina trasporta anche anidride carbonica e H+
Il legame dell’ossigeno all’emoglobina è fortemente influenzato dal pH e dall’anidride carbonica. L’emoglobina trasporta circa il 20% del totale di H+ e CO2 che si forma nei tessuti per portarlo ai polmoni e ai reni. Ai valori di pH relativamente bassi dei tessuti e alte concentrazioni di anidride carbonica, l’affinità dell’emoglobina per l’ossigeno diminuisce al crescere del legame con H+ e CO2. Nei capillari del polmone, invece, come la CO2 viene escreta e il pH del sangue sale, l’affinità della emoglobina per l’ossigeno cresce. Questo effetto è detto effetto Bohr

31
L’emoglobina trasporta anche anidride carbonica e H+
Quando si scrive: Hb + O2 ==== HbO2 Non è completo perché bisogna aggiungere l’ H+ HHb + O2 ==== H+ +HbO2 Forma protonata Hb Cioè l’emoglobina ossigenata è un acido più forte della emoglobina non ossigenata Quando l’Hb lega l’ossigeno si deprotona L’ossigeno e il protone non si legano allo stesso sito. Il protone si lega ad uno dei gruppi R degli aminoacidi che compongono l’Hb che, quindi, quando la proteina passa da ossigenata a desossi, cambia il suo pK, che diviene più alto, e si protona. Il maggior contributo all’effetto Bohr è dato dalla His -146 (His HC3)della subunità beta. Quando è protonato questo residuo forma una coppia ionica con l’Asp 84 (Asp FG1) che è una delle coppie ioniche che stabilizzano la forma desossi
della subunità beta. Quando è protonato questo residuo forma una coppia ionica con l’Asp 84 (Asp FG1) che è una delle coppie ioniche che stabilizzano la forma desossi.")
32
L’emoglobina trasporta anche anidride carbonica e H+

33
L’emoglobina trasporta anche anidride carbonica e H+
La coppia ionica con l’Asp stabilizza la forma protonata dell’His che infatti ha un pK anomalo. Quando la proteina passa nella sua forma R (legando l’ossigeno) il pK torna ad essere normale (6.0) perché nel movimento conformazionale la coppia ionica si rompe e di conseguenza l’His si deprotona. Quando la concentrazione di H+ sale nei tessuti la His si riprotona, l’ossigeno viene rilasciato e la forma T stabilizzata. Anche altri gruppi possono partecipare al trasporto dei protoni, per esempio i gruppi aminoterminali delle subunità alfa.
 il pK torna ad essere normale (6.0) perché nel movimento conformazionale la coppia ionica si rompe e di conseguenza l’His si deprotona. Quando la concentrazione di H+ sale nei tessuti la His si riprotona, l’ossigeno viene rilasciato e la forma T stabilizzata. Anche altri gruppi possono partecipare al trasporto dei protoni, per esempio i gruppi aminoterminali delle subunità alfa.")
35
L’emoglobina trasporta l’anidride carbonica
L’anidride carbonica viene legata dall’emoglobina in correlazione inversa all’ossigeno (come per il protone). La CO2 si lega ai gruppi amminici terminali sottoforma di carbammato. Questa reazione produce H+ e contribuisce all’effetto Bohr. O- CO NH2 ==== C—NH H+ O
. La CO2 si lega ai gruppi amminici terminali sottoforma di carbammato. Questa reazione produce H+ e contribuisce all’effetto Bohr. O- CO2 + --NH2 ==== C—NH-- + H+ O.")
Presentazioni simili















>")

>")


>")
