Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Una visione sistematica per l’ingegneria metabolica
Creazione di vie metaboliche

2
Ingegneria metabolica
probabilmente efficace probab. inefficace (1) (2) (3) (4) (5) (6) (7) (8) (9) S S S S S S S S S + TF A A A A A + A A A A + Y B B B B E B + B B B B Z W C C Q C F C C C C C X P P P Q P P P P P P ATP
 (2) (3) (4) (5) (6) (7) (8) (9) S. S. S. S. S. S. S. S. S. + TF. A. A. A. A. A. + A. A. A. A. + Y. B. B. B. B. E. B. + B. B. B. B. Z. W. C. C. Q. C. F. C. C. C. C. C. X. P. P. P. Q. P. P. P. P. P. P. ATP.")
3
Espressione di nuovi enzimi
Q (3) P C A B S F E (4) P C A B Q S Ingegneria metabolica dei lipidi Olii ad alto contenuto di ac. Laurico (C12) Ac. grassi poli-insaturi (Ac. Erucico) Olii ad alto contenuto di acidi monoinsaturi (ac. oleico) Olii con acido ricinoleico Manipolazioni efficaci che riguardano uno o pochi enzimi
 P. C. A. B. S. F. E. (4) P. C. A. B. Q. S. Ingegneria metabolica dei lipidi. Olii ad alto contenuto di ac. Laurico (C12) Ac. grassi poli-insaturi (Ac. Erucico) Olii ad alto contenuto di acidi monoinsaturi (ac. oleico) Olii con acido ricinoleico. Manipolazioni efficaci che riguardano uno o pochi enzimi.")
4
Knutzon et al., (1999) LPAAT from coconut endosperm mediates the insertionof laurate at the sn-2 position of triacylglycerols in Lauric rapeseed oil and can increase total laurate levels. Plant Physiology 120: Katavic et al., (2000) Utility of the Arabidopsis FAE1 and yeast SLC1-1 genes for improvements in erucic acid and oil content in rapeseed. Biochem Soc Trans. 28:935-7. Mietkiewska E et al., (2004) Seed-specific heterologous expression of a nasturtium FAE gene in Arabidopsis results in a dramatic increase in the proportion of erucic acid. Plant Physiol. 136: Stoutjesdijk PA et al., (2002) hpRNA-mediated targeting of the Arabidopsis FAD2 gene gives highly efficient and stable silencing. Plant Physiol. 129: Liu Q, Singh SP, Green AG. (2002) High-stearic and High-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiol. 129: Jain et al., (2000) Enhancement of seed oil content by expression of glycerol-3-phosphate acyltransferase genes. Biochem Soc Trans. 28: Klaus D, Ohlrogge JB, Neuhaus HE, Dormann P. (2004) Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta. 219: J. Burgal, J. Shockey, C. Lu, J. Dyer, T. Larson, I. Graham, J. Browse, Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil, Plant Biotechnol J. 6 (2008) Andrianov W et al. (2010) Tobacco as a production platform for biofuel: overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotech. J 8:277–287 Broun P, Gettner S, Somerville C. (1999) Genetic engineering of plant lipids. Annu Rev Nutr. 19: Review. Maisonneuve et al. (2010) Expression of rapeseed microsomal lysophosphatidic acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol. 152:
 Utility of the Arabidopsis FAE1 and yeast SLC1-1 genes for improvements in erucic acid and oil content in rapeseed. Biochem Soc Trans. 28: Mietkiewska E et al., (2004) Seed-specific heterologous expression of a nasturtium FAE gene in Arabidopsis results in a dramatic increase in the proportion of erucic acid. Plant Physiol. 136: Stoutjesdijk PA et al., (2002) hpRNA-mediated targeting of the Arabidopsis FAD2 gene gives highly efficient and stable silencing. Plant Physiol. 129: Liu Q, Singh SP, Green AG. (2002) High-stearic and High-oleic cottonseed oils produced by hairpin RNA-mediated post-transcriptional gene silencing. Plant Physiol. 129: Jain et al., (2000) Enhancement of seed oil content by expression of glycerol-3-phosphate acyltransferase genes. Biochem Soc Trans. 28: Klaus D, Ohlrogge JB, Neuhaus HE, Dormann P. (2004) Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta. 219: J. Burgal, J. Shockey, C. Lu, J. Dyer, T. Larson, I. Graham, J. Browse, Metabolic engineering of hydroxy fatty acid production in plants: RcDGAT2 drives dramatic increases in ricinoleate levels in seed oil, Plant Biotechnol J. 6 (2008) Andrianov W et al. (2010) Tobacco as a production platform for biofuel: overexpression of Arabidopsis DGAT and LEC2 genes increases accumulation and shifts the composition of lipids in green biomass. Plant Biotech. J 8:277–287. Broun P, Gettner S, Somerville C. (1999) Genetic engineering of plant lipids. Annu Rev Nutr. 19: Review. Maisonneuve et al. (2010) Expression of rapeseed microsomal lysophosphatidic acid acyltransferase isozymes enhances seed oil content in Arabidopsis. Plant Physiol. 152:")
5
Importanza degli olii Produzione di olio vegetale mondiale: ca. 120 Mt/anno (50 Billions $) 20 Kg/persona/anno; usati in gran parte per l’alimentazione Usati anche per la produzione di saponi, detergenti, lubrificanti, combustibili (biodiesel), cosmetici, fibre e vernici… Aumenta il consumo, ma aumenta anche la produzione, per cui il prezzo rimane abbastanza costante (0.6 $/Kg) In ordine di importanza: soia, palma, colza, girasole (>70% della prod.) Globally, over 126 million tonnes of oils were consumed in 2006/2007. Palm and soybean oils are consumed the most at 38.9 and 37.5 million tonnes respectively, followed by rapeseed oil (including canola oil) at 18.9 million tonnes and sunflower oil at 10.5 million tonnes.
, cosmetici, fibre e vernici… Aumenta il consumo, ma aumenta anche la produzione, per cui il prezzo rimane abbastanza costante (0.6 $/Kg) In ordine di importanza: soia, palma, colza, girasole (>70% della prod.) Globally, over 126 million tonnes of oils were consumed in 2006/2007. Palm and soybean oils are consumed the most at 38.9 and 37.5 million tonnes respectively, followed by rapeseed oil (including canola oil) at 18.9 million tonnes and sunflower oil at 10.5 million tonnes.")
6
Palm oil production Fresh Fruit Bunches (FFB) (circa 1000 seeds/bunch)
A palm oil mill produces crude palm oil and kernels, as primary products and biomass as secondary product. The capacity of mills varies between tons FFB/h.

7
Usi non alimentari: saponi
The word 'soap' comes from the Celtic word Saipo. Wood ash was mixed with water, then animal fat was added. When the mixture boiled, more and more ash was added as the water evaporated. Soap cleaned clothes because it soaked into the fabric and lifted up dirt, which was then carried away in the rinse water.

8
Detergent making Soap making Fuel making... methanolysis (FAME)
Soap making La materia prima per produrre saponi e detergenti sono gli ACIDI GRASSI (i detergenti sono essenz. come i saponi ma non precipitano con Ca++ o Mg++) Detergent making In the making of soap, a vegetable oil such as palm oil is boiled with a concentrated alkali solution in a reaction known as saponification. The alkali reacts with the oil and breaks it down into glycerol and fatty acids. These fatty acids react with the alkali to form sodium or potassium salt of the fatty acids. Read more: Fuel making... methanolysis (FAME)
 Detergent making. In the making of soap, a vegetable oil such as palm oil is boiled with a concentrated alkali solution in a reaction known as saponification. The alkali reacts with the oil and breaks it down into glycerol and fatty acids. These fatty acids react with the alkali to form sodium or potassium salt of the fatty acids. Read more: Fuel making... methanolysis (FAME)")
9
Acidi grassi Molecole con parte polare e parte apolare
Gli acidi grassi nelle piante sono lineari con atomi di C La maggior parte di quelli estratti da pianta contengono: linoleate, palmitate, laurate e oleate. C16: acido palmitico

10
Acidi grassi: saturi ed insaturi
Le insaturazioni influenzano l’impaccamento dei lipidi nelle membrane e quindi la temperatura a cui “cristallizzano”

11
Chiaramente per un C16. Per un C18, l’insaturazione si sposta avanti di un’unità C2
Palmitico, stearico e oleico sono i più abbondanti, L’insaturazione più comune è tra C7 e C8*

12
Sintesi glicerolipidi
sn-1 sn-2 sn-3 I carboni del glicerolo3P sono identificati con questi simboli Sintesi del Diacilglicerolofosfato (acido fosfatidico)
")
13
Di e tri-acilgliceroli
Olii e grassi (animali e vegetali)
")
14
Destino degli acidi grassi
Sintesi di lipidi nel cloroplasto AT Esporto al RE FAT

15
Due vie Entrambe le vie funzionano nelle piante

16
Overview of lipid synthesis in plants
Via eucariotica Lipidi plastidiali derivati dalla via eucariota Via procariotica Marcatore della via procariotica Somerville and Browse (1996) Trends Cell Biology 6:
 Trends Cell Biology 6:")
17
Sintesi degli acidi grassi (cloroplasto)
")
18
Sintesi di Acetil-CoA e Malonil-CoA
Oxaloacetato Citrato Malonil-CoA Acetil-CoA

19
Acetyl CoA Carboxylase as RLS
Although biochemical analysis indicates that Acetyl CoA carboxylase is a major metabolic control point in fatty acid synthesis (ref…), its overexpression had only minor impact on seed oil content (ref…), probably due… Ohlrogge J. et al, (2000) Fatty acid synthesis: from CO2 to functional genomics Biochem Soc Trans. 28: Review Balle! Il Malonil-CoA viene regolato a livello del demand. Aumentare il supply non comporta un aumento di flusso (incorporazione nei lipidi)
, its overexpression had only minor impact on seed oil content (ref…), probably due… Ohlrogge J. et al, (2000) Fatty acid synthesis: from CO2 to functional genomics Biochem Soc Trans. 28: Review. Balle! Il Malonil-CoA viene regolato a livello del demand. Aumentare il supply non comporta un aumento di flusso (incorporazione nei lipidi)")
20
Enzimi coinvolti ACC Malonyl-CoA transacilasi KAS III, II & I
FAS - Acido grasso sintasi

21
Mutanti, mutanti, mutanti...
Grandi quantità di lipidi sono scambiate tra ER e cloroplasto

22
La via cloroplastica solitamente produce C16-C18
I geni Fat terminano la crescita della catena di acido grasso (staccano la catena) e determinano quindi la sua lunghezza. Hanno attività come acyl-ACP thioesterases (FatA and FatB classes) Gli acidi C16-C18 servono: * sintesi triacilgliceroli * Sintesi galattolipidi * Sintesi fosfolipidi * Sintesi acidi grassi a catena più lunga
 e determinano quindi la sua lunghezza. Hanno attività come. acyl-ACP thioesterases (FatA and FatB classes) Gli acidi C16-C18 servono: * sintesi triacilgliceroli. * Sintesi galattolipidi. * Sintesi fosfolipidi. * Sintesi acidi grassi a catena più lunga.")
23
C16-C18 Lipidi di riserva Lipidi di membrana (via procarioti)
(via eucarioti) Lipidi a lunga catena Lipidi modificati -ossidazione
 Lipidi a lunga catena. Lipidi modificati. -ossidazione.")
24
Lipidi di riserva Una volta attivati tramite CoA, gli acidi grassi possono essere esterificati con glicerolo 3-fosfato (G-3-P) per produrre LPA, PA, DAG e TAG.
 per produrre LPA, PA, DAG e TAG.")
26
Oil bodies I triacilgliceroli (TAG) si accumulano negli Oil bodies che si originano a livello dell’ER Esistono vari tipi di olii: Saturi C16-C18 a sn-1 e 3 (es. cacao) Saturi C8-C14 in tutte le posizioni (es. cocco) Insaturi (oliva, colza…) Canola production worth B $ per year to Western Canada 1% increase in oil content worth million $
 Saturi C8-C14 in tutte le posizioni (es. cocco) Insaturi (oliva, colza…) Canola production worth B $ per year to Western Canada. 1% increase in oil content worth million $")
27
I “lipid bodies” sono circondati da oleosina

28
FAE – acido grasso elongasi
Lipidi a lunga catena FAE – acido grasso elongasi Acidi grassi a lunga catena (Very Long Chain Fatty Acids -VLCFA) Altre modificazioni possibili sono: * Epossidazione * Ossidrilazione * Ciclizzazione * … La sovraespressione di FAE porta alla formazione di una maggior proporzione di VLCFA
 Altre modificazioni possibili sono: * Epossidazione. * Ossidrilazione. * Ciclizzazione. * … La sovraespressione di FAE porta alla formazione di una maggior proporzione di VLCFA.")
29
Gli acidi grassi vengono allungati oltre C18 nel reticolo endoplasmatico

30
Lipidi modificati Acidi grassi inusuali
Diverse di queste strutture sono usate come precursori nella chimica di sintesi (nylon…) “Plant factories” per la produzione di detergenti, plastica, lubrificanti, fibre… Esempio: 1-Octene is a high-demand feedstock with a global consumption of over half a million tons per year that is primarily used as a comonomer in the expanding production of linear low density polyethylene. È ottenuto a partire da acidi grassi ω-7 come ac. palmitoleico o cis-vaccenico. Plants synthesize >200 different FA structures with attractive functional properties
 Plant factories per la produzione di detergenti, plastica, lubrificanti, fibre… Esempio: 1-Octene is a high-demand feedstock with a global consumption of over half a million tons per year that is primarily used as a comonomer in the expanding production of linear low density polyethylene. È ottenuto a partire da acidi grassi ω-7 come ac. palmitoleico o cis-vaccenico. Plants synthesize >200 different FA structures with attractive functional properties.")
31
Erucic acid U$6.50/kg (for 1,000kgs)
Erucamide, unsaturated long chain carboxylic acid amide (22:1 n-9), is used as a slip agent , anti-fogging or lubricant for plastic films (polyolefin) which can be used in food packing material. It is used as a dispersant in printing and dying. It is used in paper and textile industry for water-proof as well as corrosion inhibitor in oil wells. It is used for the synthesis of organic chemicals and surfactants used in detergent, ore floating agent, fabric softener, anti-static agent, germicide, insecticide, emulsifier, anti-caking agent, lubricant and water treatment agent. La variabilità genetica nelle varie specie è già stata abbondantemente sfruttata per creare varietà con profili diversi di acidi grassi...
, is used as a slip agent , anti-fogging or lubricant for plastic films (polyolefin) which can be used in food packing material. It is used as a dispersant in printing and dying. It is used in paper and textile industry for water-proof as well as corrosion inhibitor in oil wells. It is used for the synthesis of organic chemicals and surfactants used in detergent, ore floating agent, fabric softener, anti-static agent, germicide, insecticide, emulsifier, anti-caking agent, lubricant and water treatment agent. La variabilità genetica nelle varie specie è già stata abbondantemente sfruttata per creare varietà con profili diversi di acidi grassi...")
32
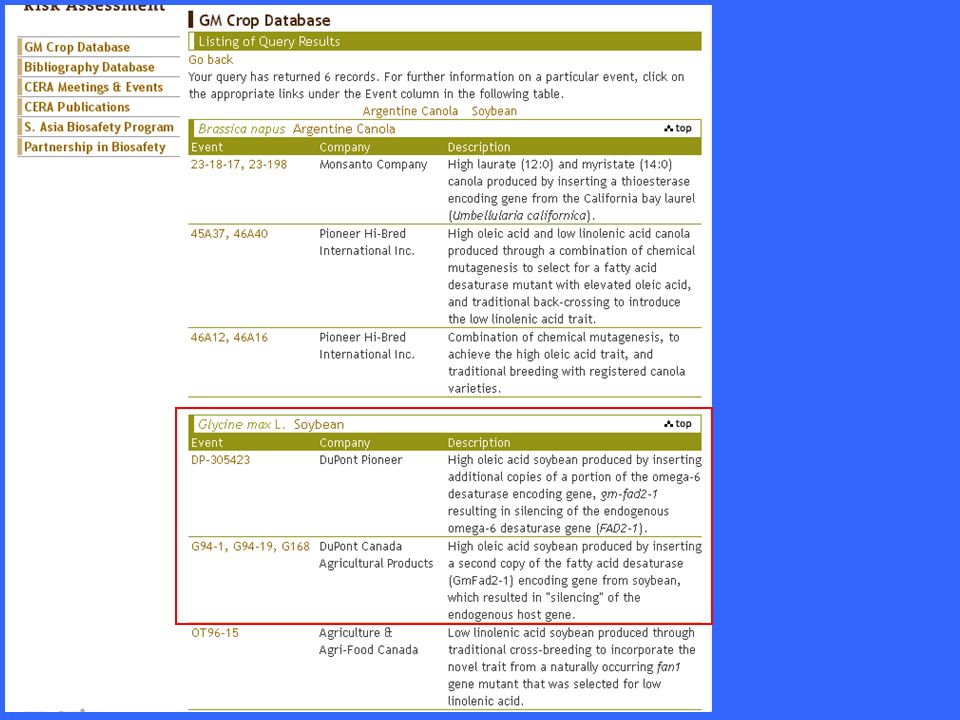
erucic acid Canola (CANadian Oil Low Acid): varietà di colza (rapeseed) creata per incrocio e selezione per ridurre il contenuto di ac. Erucico (che era ritenuto tossico) Analogamente per migliorare le qualità alimentari dell’olio di lino ftp://ftp.fao.org/es/esn/food/bio-10t.pdf
: varietà di colza (rapeseed) creata per incrocio e selezione per ridurre il contenuto di ac. Erucico (che era ritenuto tossico) Analogamente per migliorare le qualità alimentari dell’olio di lino. ftp://ftp.fao.org/es/esn/food/bio-10t.pdf.")
33
...e dell’olio di girasole
...e di soia

34
Manipolazioni del metabolismo
Large-scale new industrial uses of engineered plant oils are on the horizon but will require a better understanding of factors that limit the accumulation of unusual fatty acid structures in seeds Thelen & Ohlrogge (2002) Major goals: • Increase content of ‘‘healthy’’ fatty acids and reduce ‘‘unhealthy’’ fatty acids. • Improve oil stability to expand applications and reduce the need for hydrogenation. • Expand the repertoire of fatty acids available at low cost and high volume through exploitation of genetic diversity and enzyme engineering. • Increase oil content to reduce production costs. Più recentemente si è sfruttata anche l’ingegneria genetica in combinazione con il breeding
 Major goals: • Increase content of ‘‘healthy’’ fatty acids and reduce ‘‘unhealthy’’ fatty acids. • Improve oil stability to expand applications and reduce the need for hydrogenation. • Expand the repertoire of fatty acids available at low cost and high volume through exploitation of genetic diversity and enzyme engineering. • Increase oil content to reduce production costs. Più recentemente si è sfruttata anche l’ingegneria genetica in combinazione con il breeding.")
35
Reazioni plastidiali di sintesi e modificazione degli acidi grassi esplorate nei transgeni (il numero accanto alle frecce corrisponde al numero in tabella nella colonna evidenziata in verde) Thelen and Ohlrogge (2002)
")
36
Incorporazione degli acidi grassi nei trigliceridi
Reazioni plastidiali di sintesi e modificazione degli acidi grassi esplorate nei transgeni Thelen and Ohlrogge (2002)
")
37
Selected Examples of Fatty Acid Engineering in Transgenic Plants

38
Modificazione del profilo di acidi grassi in soia
The first trait-modified crop introduced by Monsanto, by Pioneer Hi-bred a division of DuPont and by Asoyia is low linolenic soybean. Achieved through conventional breeding methods using marker assisted selection, these seeds have reduced the linolenic acid level to under 3% (Monsanto and DuPont) and 1% (Asoyia), reducing oxidation potential. Oil from these seeds primarily is used for frying and spray coating on snack foods and crackers. Use of this oil improves the stability against oxidation and extends product shelf life and fry life compared to liquid soybean oil. Not having to hydrogenate the oil, trans fat levels in food are significantly reduced while maintaining or slightly reducing saturated fat levels. Wilkes RS (2008) Low linolenic soybeans and beyond. Lipid Technology 20:
 and 1% (Asoyia), reducing oxidation potential. Oil from these seeds primarily is used for frying and spray coating on snack foods and crackers. Use of this oil improves the stability against oxidation and extends product shelf life and fry life compared to liquid soybean oil. Not having to hydrogenate the oil, trans fat levels in food are significantly reduced while maintaining or slightly reducing saturated fat levels. Wilkes RS (2008) Low linolenic soybeans and beyond. Lipid Technology 20:")
40
TREUSTM brand High Oleic Soybean TREUS™ Low Linolenic Soybean Oil
Low linolenic soybean oil produced from Pioneer® brand soybean varieties, and previously marketed as NUTRIUM Low Linolenic Soybean Oil, will now be marketed as TREUS™ Low Linolenic Soybean Oil. Acres planted with Pioneer® brand low linolenic soybeans grew from about 35,000 in 2005, 200,000 in 2006, 500,000 in 2007, 1.8 million acres in 2008. VISTIVE is a new range of winter oilseed rape with a high oleic, low linolenic fatty acid oil profile (HOLL) (conventional)
 (conventional)")
41
Soia ad alto acido oleico

42
Because these soybeans differ from commodity soybeans in their fatty acid content, an identity preservation system has been established in the USA beginning at the farm and includes grain elevators, processors, and oil refiners to assure that customers receive identity preserved oil made from low linolenic acid soybeans.

44
Major vegetable oils currently or soon-to-be available to food processors and other manufacturers
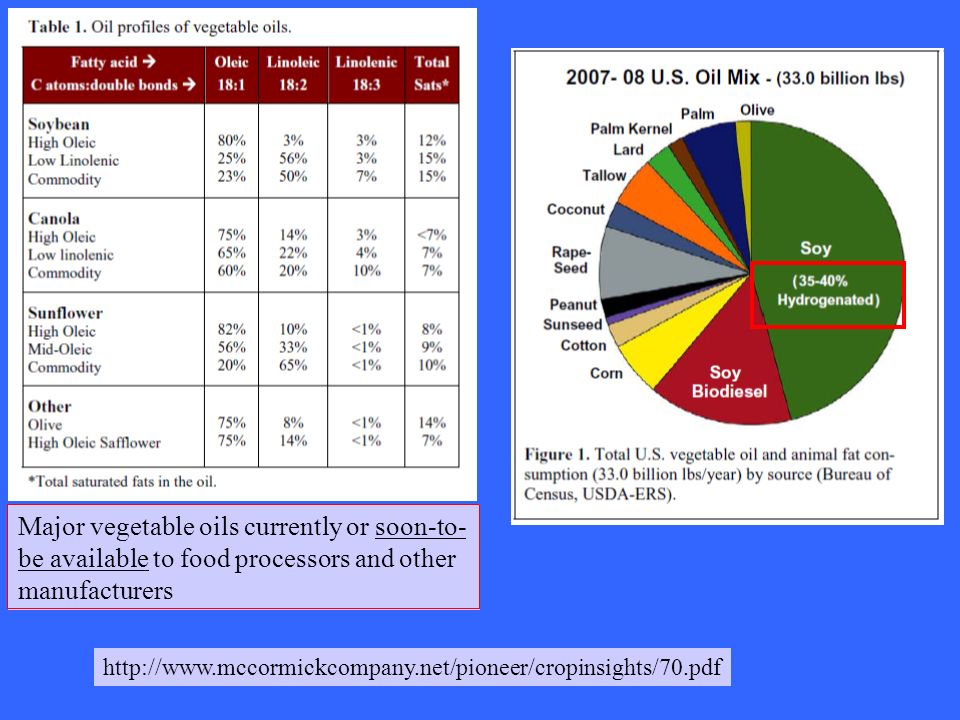
45
palmitoleic 16:1Δ9 and cis-vaccenic 18:1Δ11
Nguyen et al., (2010) Metabolic engineering of seeds can achieve levels of ω-7 fatty acids comparable with the highest levels found in natural plant sources. Plant Physiol 154: A plant oil containing high (70%) content of ω-7 FA would represent a new and sustainable feedstock for 1-octene production. Heterologous expression of the milkweed desaturase in Arabidopsis failed to produce detectable ω-7 FA, and when the Doxantha desaturase was expressed in Brassica napus, it resulted in the accumulation of only approx. 9% ω-7 FA
 Metabolic engineering of seeds can achieve levels of ω-7 fatty acids comparable with the highest levels found in natural plant sources. Plant Physiol 154: A plant oil containing high (70%) content of ω-7 FA would represent a new and sustainable feedstock for 1-octene production. Heterologous expression of the milkweed desaturase in Arabidopsis failed to produce detectable ω-7 FA, and when the Doxantha desaturase was expressed in Brassica napus, it resulted in the accumulation of only approx. 9% ω-7 FA.")
46
Biochemical evidence has confirmed that the fab1 lesion is in KASII
Biochemical evidence has confirmed that the fab1 lesion is in KASII. Expression of Com25 in fab1 increased the accumulation of 16:1Δ9 and 18:1Δ11 to approx. 23% and approx. 16% respectively, yielding a total of approximately 39% ω-7 FA. … increased ω-7 FA to as much as 71%

47
Primo es.: produzione di ac. Laurico
“Niente è più insensato di una risposta a una domanda che non si pone” (R. Niehbur) Introdotta una FAT (Fatty acid -ACP Thioesterase) BTE in colza. Thelen and Ohlrogge (2002)
 Introdotta una FAT (Fatty acid -ACP Thioesterase) BTE in colza. Thelen and Ohlrogge (2002)")
48
Rapeseed BTE (Bay Thio Esterase) plants
L’olio di colza di piante wt contiene solo il 7% di ac. Grassi saturi (C16 e C18) Piante che esprimono una BTE (Bay Thio-Esterase) riescono ad accumulare fino al 60% di ac.laurico, ma non oltre Il limite è probabilmente da attribuirsi alla LPAAT endogena di colza che non riesce ad incorporare laurato in sn-2 Trasformare colza con una LPAAT da cocco che riesce a incorporare laurato in sn-2
 Piante che esprimono una BTE (Bay Thio-Esterase) riescono ad accumulare fino al 60% di ac.laurico, ma non oltre. Il limite è probabilmente da attribuirsi alla LPAAT endogena di colza che non riesce ad incorporare laurato in sn-2. Trasformare colza con una LPAAT da cocco che riesce a incorporare laurato in sn-2.")
49
LPAAT substrate specificity of CLP-expressing canola seeds
A pool of mid-maturation seeds from a control plant (white bars) and from a transgenic plant, pCGN5511-LP004-5 (black bars), were assayed for LPAAT substrate specificity using 12:0-LPA and various 14C-labeled acyl-CoAs. PA, Phosphatitic acid. Knutzon, D. S., et al. Plant Physiol. 1999;120:
 and from a transgenic plant, pCGN5511-LP004-5 (black bars), were assayed for LPAAT substrate specificity using 12:0-LPA and various 14C-labeled acyl-CoAs. PA, Phosphatitic acid. Knutzon, D. S., et al. Plant Physiol. 1999;120:")
50
Correlation of 12:0-LPAAT activity with 12:0 accumulation at sn-2
LPAAT activity using 12:0-CoA and 12:0-LPA was determined in membrane fractions derived from developing untransformed, control canola seeds, as well as from independent CLP LP004 transformants. In addition, the proportion of 12:0 at sn-2 was measured in 20-seed pools of mature F1 seeds derived from crosses of the same set of CLP plants with a homozygous BTE-containing line, DH22. The figure correlates these two determinations. , Control; ×, individual CLP × BTE F1 seed lots. PA, Phosphatitic acid. Each primary transformant was crossed with DH22, a homozygous BTE line that contains 51 mol % laurate. Since the BTE parent was homozygous, each resulting F1 seed should have a complement of BTE alleles. Knutzon, D. S., et al. Plant Physiol. 1999;120:

51
Relationship between total laurate content and trilaurin
Nelle piante con sola BTE, trilaurina era <3% anche con il laurato al 47% ×, Canola lines transformed with BTE alone ○, seeds from F2, cross: CLP X BTE plants ─, theoretical, assuming laurate at random F1 plants from crosses of several LPAAT transformants with the 50 mol % laurate containing BTE homozygous line DH22 were grown and CLP-homozygous F2 lines selected in the next generation. Le piante con entrambi i transgeni accumulano più trilaurina dell’atteso sulla base della [12:0] Seed oil was extracted from transformed canola plants and the total laurate content of the oil and proportion of trilaurin compared with other TAGs in the oil were determined. Each symbol reflects a sample derived from a seed pool from one dihaploid plant as described. ×, Canola lines transformed with BTE alone; ○, seeds from F2 plants resulting from crosses of several different CLP transformants with the homozygous BTE plant. The line was calculated by assuming that laurate was positioned randomly at all three positions of the triglyceride. Knutzon, D. S., et al. Plant Physiol. 1999;120:

52
Laurate proportion at sn-2 is dependent on total laurate levels and coconut LPAAT
○, CLP-positive plant ×, CLP-negative plant The primary CLP transformants were crossed with the homozygous BTE line. A F1 plant harboring CLP and BTE alleles was grown. Independently segregating F2 microspores derived from this plant were made diploid and grown into (homozygous) dihaploid plants as described. The presence or absence of the coconut LPAAT gene in the individual dihaploid plants was determined via PCR of leaf tissue. All plants were selfed, and oil was extracted from the resulting seeds. The sn-2 analysis of seed oil was executed using R. arrhizus lipase. Each symbol represents a seed pool derived from one dihaploid plant. ○, CLP-positive plant; ×, CLP-negative plant. For this analysis, we selected the top 12 laurate producers of the generated dihaploids, as well as randomly chosen plants throughout the laurate range. Knutzon, D. S., et al. Plant Physiol. 1999;120:
 dihaploid plants as described. The presence or absence of the coconut LPAAT gene in the individual dihaploid plants was determined via PCR of leaf tissue. All plants were selfed, and oil was extracted from the resulting seeds. The sn-2 analysis of seed oil was executed using R. arrhizus lipase. Each symbol represents a seed pool derived from one dihaploid plant. ○, CLP-positive plant; ×, CLP-negative plant. For this analysis, we selected the top 12 laurate producers of the generated dihaploids, as well as randomly chosen plants throughout the laurate range. Knutzon, D. S., et al. Plant Physiol. 1999;120:")
53
Coconut LPAAT can boost laurate levels
Le piante CLP- non riescono ad accumulare oltre il 60% di laurato. I transgeni con la sola BTE si fermano al 60% di laurato ( non riescono ad andare oltre) Each symbol represents a seed-pool analysis of an individual dihaploid plant. All dihaploid plants resulting from crosses described in Figure 4 were sorted via PCR into a CLP-containing (CLP +) and a CLP-free (CLP ) populations. Plants of both populations were grouped into 1% laurate intervals. Knutzon, D. S., et al. Plant Physiol. 1999;120:
 Each symbol represents a seed-pool analysis of an individual dihaploid plant. All dihaploid plants resulting from crosses described in Figure 4 were sorted via PCR into a CLP-containing (CLP +) and a CLP-free (CLP ) populations. Plants of both populations were grouped into 1% laurate intervals. Knutzon, D. S., et al. Plant Physiol. 1999;120:")
54
Correlation of oil levels with BTE and CLP.
L’eccesso viene degradato Nelle piante CLP- esiste un limite (60%) nell’accumulo di laurato, limite che viene superato nelle piante CLP+ (che esprimono LPAAT) The total seed oil mass as a percentage of dry weight of the dihaploid plants described in Figure 4 were determined by NMR. The data are shown separately for CLP-free (CLP-) and CLP-containing (CLP+) populations, with the oil percentage plotted against total laurate levels
 nell’accumulo di laurato, limite che viene superato nelle piante CLP+ (che esprimono LPAAT) The total seed oil mass as a percentage of dry weight of the dihaploid plants described in Figure 4 were determined by NMR. The data are shown separately for CLP-free (CLP-) and CLP-containing (CLP+) populations, with the oil percentage plotted against total laurate levels.")
55
Successful increase in lipid content by increasing demand
(Bouvier-Nave et al., 2000; Maisonneuve et al., 2010; Oakes et al., 2011; Petrie et al., 2012; Taylor et al., 2001, 2009b; Zheng et al., 2008) especially mediated by augmenting the diacylglycerol transacylase (DGAT) activity, the major TAG biosynthetic enzyme (for more examples, see Table 1). When a change in composition without substantial increase in overall content is the desired target, this is achieved by expressing the required biosynthetic enzymes (be they thioesterases, desaturases, elongases or hydroxylases) as well as the acyltransferases or other activities that are able to incorporate the specific fatty acids into lipids/TAG (Burgal et al., 2008; Hoffmann et al., 2008; Li et al., 2010; Mietkiewska et al., 2004; Nguyen et al., 2010; Ruiz-Lopez et al., 2012a; Sayanova et al., 2012; Taylor et al., 2009a; Truksa et al., 2006; Wilkes, 2008; Wu et al., 2005; additional examples in Ruiz-Lopez et al., 2012b). traditional Kennedy pathway (green) The increased preference for hydroxyacids by a DGAT or a phospholipid:diacylglycerol acyltransferase from Ricinus stimulated the incorporation of ricinoleic acid into TAG from 17% to respectively 30% and 25% of total seed lipid (Burgal et al., 2008; Kim et al., 2011), and similar situations were reported for lauric (Knutzon et al., 1999) and vernolic acid (Li et al., 2010). When the supply of fatty acids is not matched by an increase in demand, a futile cycle of fatty acid degradation via b-oxidation and sucrose re-synthesis is triggered (Moire et al., 2004; Poirier et al., 1999; Voelker et al., 1996). Main enzymes: ACS, acyl- CoA synthase; DGAT, diacylglycerol acyltransferase; GPAT, glycerol-3-phosphate acyltransferase; LPAAT, lysophosphatidic acid acyltransferase; LPCAT, lysophosphatidylcholine acyltransferase; PAP, 3-sn-phosphatidate phosphohydrolase; PDAT, phosphatidylcholine diacylglycerol acyltransferase; PDCT, phosphatidylcholine diacylglycerol cholinephosphotransferase; PLA, phospholipase A; PLC, phospholipase C; PLD, phospholipase D.
 especially mediated by augmenting the diacylglycerol transacylase (DGAT) activity, the major TAG biosynthetic enzyme (for more examples, see Table 1). When a change in composition without substantial increase in overall content is the desired target, this is achieved by expressing the required biosynthetic enzymes (be they thioesterases, desaturases, elongases or hydroxylases) as well as the acyltransferases or other activities that are able to incorporate the specific fatty acids into lipids/TAG (Burgal et al., 2008; Hoffmann et al., 2008; Li et al., 2010; Mietkiewska et al., 2004; Nguyen et al., 2010; Ruiz-Lopez et al., 2012a; Sayanova et al., 2012; Taylor et al., 2009a; Truksa et al., 2006; Wilkes, 2008; Wu et al., 2005; additional examples in Ruiz-Lopez et al., 2012b). traditional Kennedy pathway (green) The increased preference for hydroxyacids by a DGAT or a phospholipid:diacylglycerol acyltransferase from Ricinus stimulated the incorporation of ricinoleic acid into TAG from 17% to respectively 30% and 25% of total seed lipid (Burgal et al., 2008; Kim et al., 2011), and similar situations were reported for lauric (Knutzon et al., 1999) and vernolic acid (Li et al., 2010). When the supply of fatty acids is not matched by an increase in demand, a futile cycle of fatty acid degradation via b-oxidation and sucrose re-synthesis is triggered (Moire et al., 2004; Poirier et al., 1999; Voelker et al., 1996). Main enzymes: ACS, acyl- CoA synthase; DGAT, diacylglycerol acyltransferase; GPAT, glycerol-3-phosphate acyltransferase; LPAAT, lysophosphatidic acid acyltransferase; LPCAT, lysophosphatidylcholine acyltransferase; PAP, 3-sn-phosphatidate phosphohydrolase; PDAT, phosphatidylcholine diacylglycerol acyltransferase; PDCT, phosphatidylcholine diacylglycerol cholinephosphotransferase; PLA, phospholipase A; PLC, phospholipase C; PLD, phospholipase D.")
56
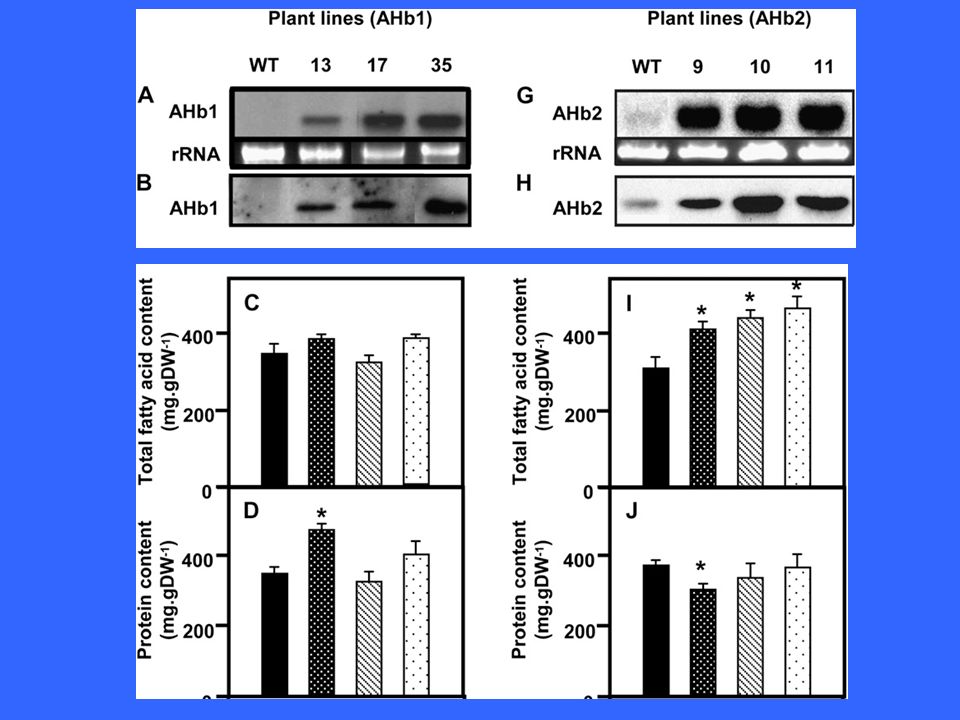
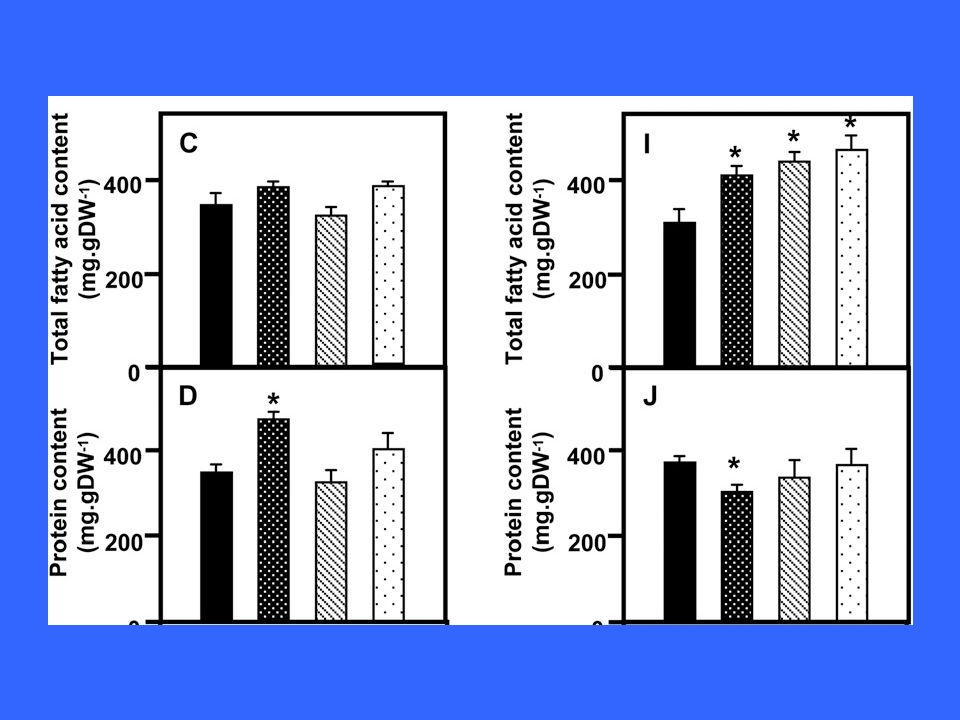
The parallel activation of many genes, especially using transcriptional regulators coordinating genes involved in TAG synthesis, achieved remarkable flux increases, even in vegetative tissues (Andrianov et al., 2010; Gao et al., 2009; Naqvi et al., 2010; Pouvreau et al., 2011; Sanjaya et al., 2011; Shen et al., 2010; Slocombe et al., 2009; reviewed in Baud and Lepiniec, 2010; Lu et al., 2011; Weselake et al., 2009). Both positive and negative regulators of oil content have been identified (LEC1 and LEC2, WRI, PKL and ASIL1) and exploited to this purpose An often overlooked factor limiting lipid accumulation in Arabidopsis seeds is oxygen, because hemoglobin overexpression boosts lipid accumulation by 40% in absolute values per seed and as percentage of seed dry weight (Vigeolas et al., 2011). The haemoglobin maintained a higher ATP/ADP ratio even under low (4%) external oxygen. However strange it may appear, seeds of various species experience an internal O2 concentration in the 2–4% range (v/v) (see references in Vigeolas et al., 2011) Vigeolas, H., Huehn, D. and Geigenberger, P. (2011) Nonsymbiotic hemoglobin-2 leads to an elevated energy state and to a combined increase in polyunsaturated fatty acids and total oil content when overexpressed in developing seeds of transgenic Arabidopsis plants. Plant Physiol. 155, 1435–1444.
. The haemoglobin maintained a higher ATP/ADP ratio even under low (4%) external oxygen. However strange it may appear, seeds of various species experience an internal O2 concentration in the 2–4% range (v/v) (see references in Vigeolas et al., 2011) Vigeolas, H., Huehn, D. and Geigenberger, P. (2011) Nonsymbiotic hemoglobin-2 leads to an elevated energy state and to a combined increase in polyunsaturated fatty acids and total oil content when overexpressed in developing seeds of transgenic Arabidopsis plants. Plant Physiol. 155, 1435–1444.")
59
Bibliografia Mittendorf et al., PNAS (1998)
Somerville and Browse (1996) Trends Cell Biology 6: Ohlrogge J. et al, (2000) Fatty acid synthesis: from CO2 to functional genomics Biochem Soc Trans. 28: Review database: A CATALOG OF GENES FOR PLANT GLYCEROLIPID BIOSYNTHESIS (Paper: TOWARDS A FUNCTIONAL CATALOG OF THE PLANT GENOME: A SURVEY OF GENES FOR LIPID BIOSYNTHESIS, Plant Physiology 122: Thelen JJ, Ohlrogge JB. (2002) Metabolic engineering of fatty acid biosynthesis in plants. Metab Eng., 4: Review. Knutzon, D. S., et al. (1999) Lysophosphatidic acid acyltransferase from coconut endosperm mediates the insertion of laurate at the sn-2 position of triacylglycerols in lauric rapeseed oil and can increase total laurate levels. Plant Physiol. 120: Klaus D, Ohlrogge JB, Neuhaus HE, Dormann P. (2004) Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta 219:
 Trends Cell Biology 6: Ohlrogge J. et al, (2000) Fatty acid synthesis: from CO2 to functional genomics Biochem Soc Trans. 28: Review. database: A CATALOG OF GENES FOR PLANT GLYCEROLIPID BIOSYNTHESIS (Paper: TOWARDS A FUNCTIONAL CATALOG OF THE PLANT GENOME: A SURVEY OF GENES FOR LIPID BIOSYNTHESIS, Plant Physiology 122: Thelen JJ, Ohlrogge JB. (2002) Metabolic engineering of fatty acid biosynthesis in plants. Metab Eng., 4: Review. Knutzon, D. S., et al. (1999) Lysophosphatidic acid acyltransferase from coconut endosperm mediates the insertion of laurate at the sn-2 position of triacylglycerols in lauric rapeseed oil and can increase total laurate levels. Plant Physiol. 120: Klaus D, Ohlrogge JB, Neuhaus HE, Dormann P. (2004) Increased fatty acid production in potato by engineering of acetyl-CoA carboxylase. Planta 219:")
Presentazioni simili











>")




>")

>")


