Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il compartimento naturale dell’immunità innata
Dr. Luigi Lembo Fazio Sapienza Università di Roma Dipartimento di Biologia e Biotecnologie “Charles Darwin”

2
Il sistema immunitario: la funzione
Riconosce gli organismi estranei “invasori” Previene/limita la loro diffusione Eradica la loro presenza nell’organismo Ritorna ad uno stato quiescente instaurando una condizione di protezione duratura (memoria immunologica) Ma non solo…. È un sistema integrato di cellule professionali che collaborano e comunicano al fine di mantenere l’omeostasi
 Ma non solo…. È un sistema integrato di cellule professionali che collaborano e comunicano al fine di mantenere l’omeostasi.")
3
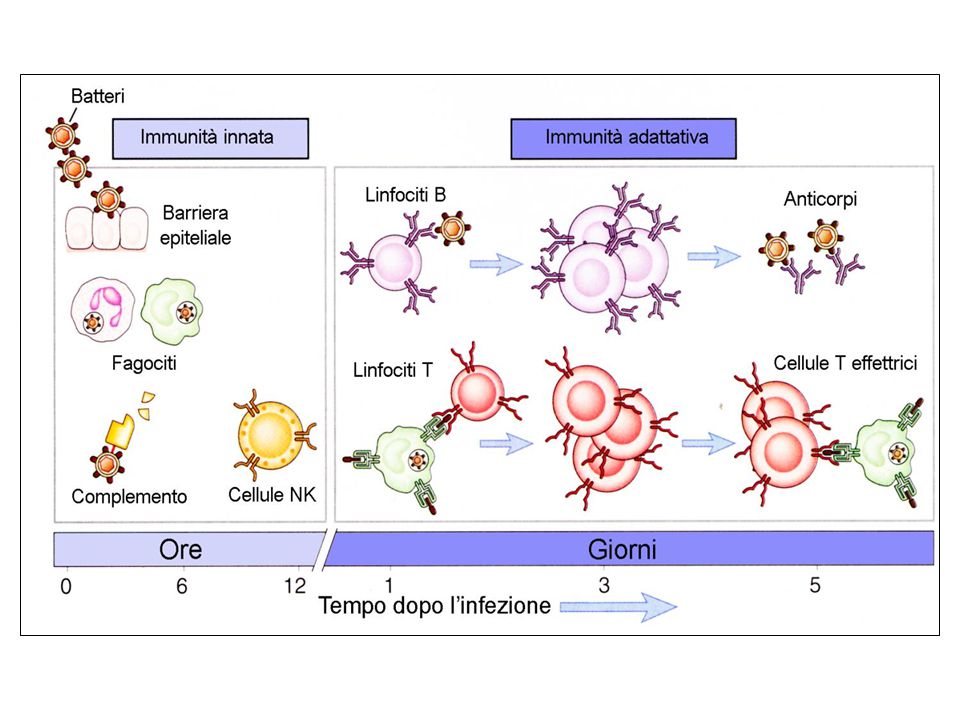
Esistono due fondamentali sistemi di riconoscimento e di difesa, correlati tra loro: quello dell’immunità innata, detta anche naturale o nativa, in quanto pre-esistente all’esposizione agli agenti microbici o alle macromolecole estranee, e quello dell’immunità adattativa. La combinazione di questi due sistemi fornisce un’efficiente difesa contro i microrganismi patogeni o più in generale contro qualsiasi agente che venga riconosciuto come “non-self”

4
I due tipi di immunità…un po’ di storia
Immunita’ innata o naturale controllo/ pronta risposta (fase precoce) limitazione dell’infezione Immunita’ adattativa o acquisita eradicazione più lenta (fase tardiva) dell’infezione Immunità di lunga durata Innata Adattativa APC
 limitazione dell’infezione. Immunita’ adattativa o acquisita eradicazione più lenta. (fase tardiva) dell’infezione Immunità di lunga durata. Innata. Adattativa. APC.")
5
Immunità innata È un meccanismo di difesa dell’ospite contro le infezioni, altamente conservato Si ritrova in tutti gli organismi pluricellulari (al contrario dell’immunità adattativa che è presente solo nei vertebrati) Prima linea di difesa sempre attiva che può indurre l’attivazione del compartimento dell’immunità adattativa Fornisce protezione verso una grande varietà di patogeni in quanto manca di specificità È in grado di distinguere tra self e non-self
 Prima linea di difesa sempre attiva che può indurre l’attivazione del compartimento dell’immunità adattativa. Fornisce protezione verso una grande varietà di patogeni in quanto manca di specificità. È in grado di distinguere tra self e non-self.")
7
Immunità innata e adattativa
Proprietà Innata Adattativa Recettori Fissati nel genoma Non è necessario il riarrangiamento Codificati in segmenti genici Il riarrangiamento è necessario Distribuzione Non clonale Tutte le cellule di una classe identica Clonale Tutte le cellule di una distinta classe Ligandi Molecole conservate Singoli elementi di molecole Discriminazione self-non self Perfetta: selezionata durante l’evoluzione Imperfetta: selezionata nelle cellule somatiche dell’individuo Tempo di azione Immediata attivazione degli effettori Ritardata attivazione degli effettori Risposta Molecole co-stimolatorie Citochine (IL-1b, IL-6) Chemochine (CXCL-8) Espansione clonale Citochine effettrici (IL-4, IFN-g)
 Chemochine (CXCL-8) Espansione clonale. Citochine effettrici (IL-4, IFN-g)")
8
L’immunità innata e l’immunità adattativa: similitudini e differenze
Acquisita Barriere chimico-fisiche Epiteli Muco pH Temperatura Complemento Anticorpi Recettori circolanti Recettori di membrana Recettori intracellulari Molecole circolanti Fagociti: Macrofagi Linfociti T e B cellule dendritiche (DC) PMN’s, cellule NK Componente cellulare Citochine Citochine (IL-1, IL-6, TNF-α) (IL-2, IFN-γ) Mediatori solubili Tessuti linfoidi associate alle mucose (MALT) Anticorpi nelle secrezioni
 PMN’s, cellule NK. Componente cellulare. Citochine Citochine. (IL-1, IL-6, TNF-α) (IL-2, IFN-γ) Mediatori solubili. Tessuti linfoidi associate alle mucose (MALT) Anticorpi nelle secrezioni.")
9
I meccanismi dell’immunità innata
e strutturali Chimico-fisiche Barriere Cellule ciliate pH Muco Epiteli Cellule Fagociti Monociti Macrofagi Cellule NK PMN’s solubili Mediatori Proteine della fase Citochine e acuta IFN-α IL-1 IL-6 TNF-α Recettori Circolanti e Molecole Complemento e PRRs PRRs di membrana PRRs citosolici PRRs circolanti

10
Le strutture batteriche

11
Peptidoglicano / Lipoproteine
Rappresentazione schematica delle componenti batteriche: i PAMPs e i fattori di virulenza PAMPs Fattori di virulenza LPS Peptidoglicano / Lipoproteine CpG DNA Flagelli A LPS CpG DNA Flagelli Tossine Plasmide Sistema di secrezione B

12
Quali le strutture di interesse?

13
Il batterio non scoppia!!
La parete batterica Protezione dai danni di tipo osmotico (più importante) Determina e mantiene la forma del batterio Protegge dalle offese meccaniche Funzione immunitaria Il batterio non scoppia!! Resistenza in terreni con pressione osmotica inferiore a quella del loro citoplasma
 Determina e mantiene la forma del batterio. Protegge dalle offese meccaniche. Funzione immunitaria. Il batterio non scoppia!! Resistenza in terreni con pressione osmotica inferiore a quella del loro citoplasma.")
14
Organizzazione della parete dei batteri Gram+ e Gram-

15
Organizzazione della parete dei batteri Gram-

17
Organizzazione della parete dei batteri Gram+ e Gram- [1]
Spesso peptidoglicano; anche 40 strati concentrici 40-80% del peso secco Altri componenti oltre al peptidoglicano: acidi teicoici, acidi teicuronici, proteine, carboidrati intercalati nel peptidoglicano GRAM – Sottile peptidoglicano; pochi strati concentrici (2-3) 5% del peso secco pochi legami crociati Altri componenti oltre al peptidoglicano: organizzati a formare una struttura complessa all’esterno della mureina: la membrana esterna
![Organizzazione della parete dei batteri Gram+ e Gram- [1]](http://slideplayer.it/slide/2327244/8/images/17/Organizzazione+della+parete+dei+batteri+Gram%2B+e+Gram-+%5B1%5D.jpg "Spesso peptidoglicano; anche 40 strati concentrici 40-80% del peso secco. Altri componenti oltre al peptidoglicano: acidi teicoici, acidi teicuronici, proteine, carboidrati intercalati nel peptidoglicano. GRAM – Sottile peptidoglicano; pochi strati concentrici (2-3) 5% del peso secco pochi legami crociati. Altri componenti oltre al peptidoglicano: organizzati a formare una struttura complessa all’esterno della mureina: la membrana esterna.")
18
Il lipopolisaccaride (LPS)
")
19
L’LPS: un po’ di storia [1]
Klebs Postula l’esistenza di una sostanza pirogena in microrganismi da lui indicati come “Microsporon septicum” responsabili della maggior parte dei decessi dei militari. Panum Ipotizza l’esistenza di una tossina pirogena, nel materiale in putrefazione, non volatile, resistente al calore e solubile in acqua.
![L’LPS: un po’ di storia [1]](http://slideplayer.it/slide/2327244/8/images/19/L%E2%80%99LPS%3A+un+po%E2%80%99+di+storia+%5B1%5D.jpg "Klebs. Postula l’esistenza di una sostanza pirogena in microrganismi da lui indicati come Microsporon septicum responsabili della maggior parte dei decessi dei militari. Panum. Ipotizza l’esistenza di una tossina pirogena, nel materiale in putrefazione, non volatile, resistente al calore e solubile in acqua.")
20
L’LPS: un po’ di storia [2]
Richard Pfeiffer (1858–1910) Lisati di batteri precedentemente uccisi al calore (V. cholerae) erano in grado di indurre una reazione di shock tossico quando somministrati nel modello del guinea pig. Postula l’esistenza di un principio tossico, resistente al calore, che definisce Endotossina William B. Coley (1862–1936) Trattamento del sarcoma mediante somministrazione Streptococci Gram positivi e batteri Gram negativi (S. marcescens) vitali o inattivati con il calore. Tossina di Coley: questo primo tentativo di terapia vaccinale portava a una completa remissione dei tumori.
![L’LPS: un po’ di storia [2]](http://slideplayer.it/slide/2327244/8/images/20/L%E2%80%99LPS%3A+un+po%E2%80%99+di+storia+%5B2%5D.jpg "Richard Pfeiffer (1858–1910) Lisati di batteri precedentemente uccisi al calore (V. cholerae) erano in grado di indurre una reazione di shock tossico quando somministrati nel modello del guinea pig. Postula l’esistenza di un principio tossico, resistente al calore, che definisce Endotossina. William B. Coley (1862–1936) Trattamento del sarcoma mediante somministrazione Streptococci Gram positivi e batteri Gram negativi (S. marcescens) vitali o inattivati con il calore. Tossina di Coley: questo primo tentativo di terapia vaccinale portava a una completa remissione dei tumori.")
21
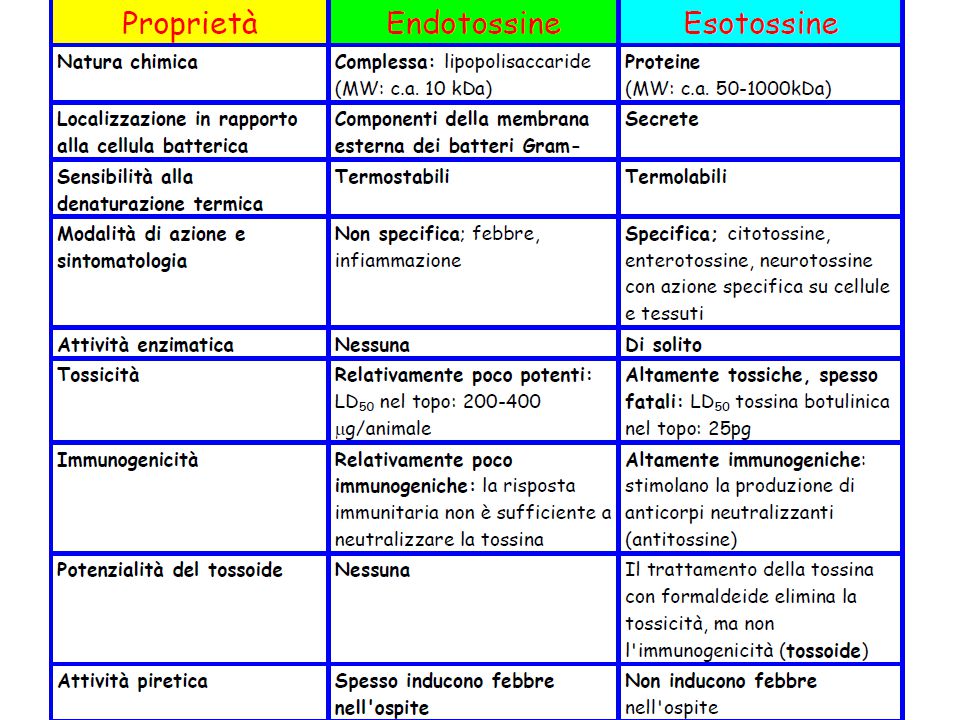
L’LPS: generalità Endotossine (per distinguerle dalle esotossine)
Molecole anfifiliche stabili al calore (massa di 10 kDa) Componente essenziale della membrana esterna di batteri Gram-negativi, commensali e patogeni: E. coli, S. typhimurium, N. meningitidis, H. influenzae, B. pertussis, P. aeruginosa, H. pylori, K. pneumoniae, L. pneumophila, C. trachomatis Diverse specie del genere Sphingomonas non possiedono LPS ma glicosfingolipidi (GSL)
 Componente essenziale della membrana esterna di batteri Gram-negativi, commensali e patogeni: E. coli, S. typhimurium, N. meningitidis, H. influenzae, B. pertussis, P. aeruginosa, H. pylori, K. pneumoniae, L. pneumophila, C. trachomatis. Diverse specie del genere Sphingomonas non possiedono LPS ma glicosfingolipidi (GSL)")
24
Porzione oligo/polisaccaridica
L’LPS: la struttura [1] Regione lipofilica, il Lipide A Regione oligo/polisaccaridica idrofilica legata covalentemente Porzione oligo/polisaccaridica Lipide A Stabilizzazione delle molecole di LPS mediante il legame con cationi divalenti Funzione di barriera impermeabile (Polimixine, poliamine, peptidi e proteine cationiche, agenti chelanti [EDTA])
")
26
L’LPS: la struttura [2] In E. coli sono presenti circa 3 x 106 molecole di LPS (4,9 mm2 vs 6,7 mm2) Principale componente della membrana (45 %); il 75 % della superficie della membrana esterna è occupato dall’LPS I lipopolisaccaridi tendono a formare aggregati L’architettura strutturale dell’LPS è molto conservata e comparabile nell’ambito delle Enterobacteriaceae (Salmonella spp., E. coli, Shigella) In questo modulo comune si ritrovano varianti strutturali: 1. Diversità nella composizione chimica della regione polisaccaridica 2. Diversità nella struttura del Lipide A
![L’LPS: la struttura [2] In E. coli sono presenti circa 3 x 106 molecole di LPS (4,9 mm2 vs 6,7 mm2)](http://slideplayer.it/slide/2327244/8/images/26/L%E2%80%99LPS%3A+la+struttura+%5B2%5D+In+E.+coli+sono+presenti+circa+3+x+106+molecole+di+LPS+%284%2C9+mm2+vs+6%2C7+mm2%29.jpg "Principale componente della membrana (45 %); il 75 % della superficie della membrana esterna è occupato dall’LPS. I lipopolisaccaridi tendono a formare aggregati. L’architettura strutturale dell’LPS è molto conservata e comparabile nell’ambito delle Enterobacteriaceae (Salmonella spp., E. coli, Shigella) In questo modulo comune si ritrovano varianti strutturali: 1. Diversità nella composizione chimica della regione polisaccaridica. 2. Diversità nella struttura del Lipide A.")
27
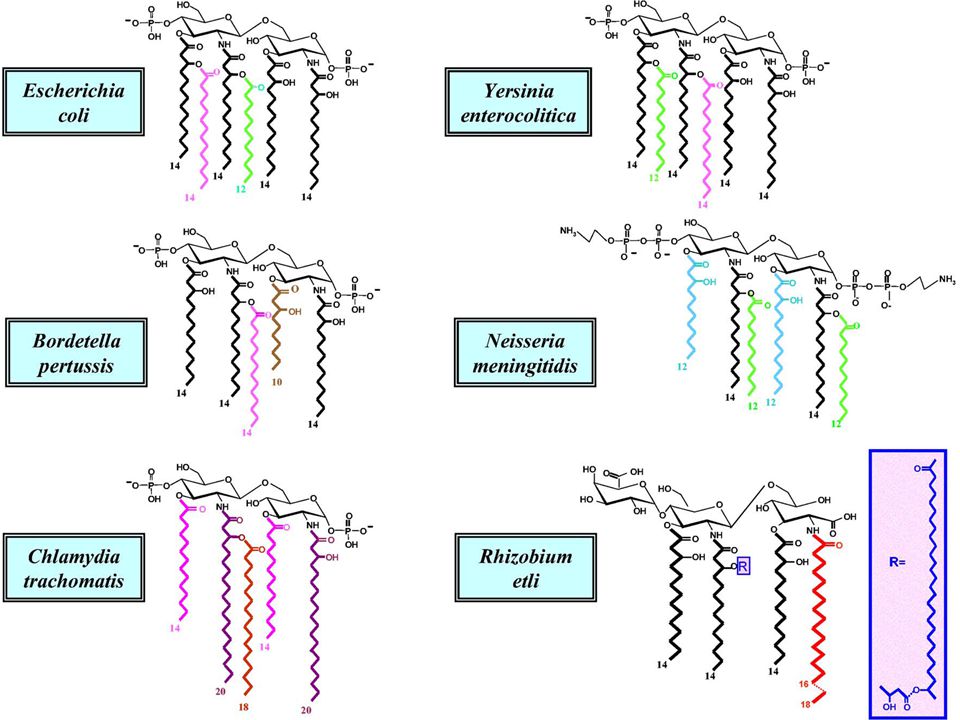
Lipide A: struttura delle Enterobacteriaceae
Legame a-glicosidico estere Scheletro di D-glucosamine lo scheletro è costituito da un disaccaride con legame b,1-6 sono presenti due gruppi fosforici (solitamente il Lipide A è monofosforilato) sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi Posizione di attacco della regione polisaccaridica
 sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi. Posizione di attacco della regione polisaccaridica.")
28
Lipide A: il caso di E. coli
Il lipide A di E. coli viene generalmente riportato come esa-acilato; in quantità più o meno variabili anche le specie penta e tetracilate. Gli acidi grassi principali hanno un numero di atomi di carbonio compresi tra C10 e C16 C18 e C21 in H. pylori e C. trachomatis Il numero di catene aciliche varia considerevolmente e ha un effetto diretto sulla tossicità dell’LPS Gli acidi grassi idrossilati sono legati allo scheletro glucidico mediante legami esterici o amidici in posizione 2 e 2’ e 3 e 3’

29
K12

30
Lipide A: variazioni nei gruppi fosforici
N. meningitidis LipideA Fosfatidil-etanolamina Le principali sostituzioni presenti nel Lipide A dei Gram-negativi sono: Gruppi pirofosforici (PP), etilamina, etanolamina, difosfoetanolamina Residui saccaridici: L-arabinosio; L,D-glicero-D-manno-eptosio; acido D-galatturonico
, etilamina, etanolamina, difosfoetanolamina. Residui saccaridici: L-arabinosio; L,D-glicero-D-manno-eptosio; acido D-galatturonico.")
31
Lipide A: variazioni del pattern di acilazione
Le principali variazioni riguardano: Il numero delle catene di acidi grassi La posizione relativa delle catene di acidi grassi La natura dei gruppi acilici - acido miristico - acido palmitico - acido stearico - acido laurico Legame a-glicosidico estere Scheletro di D-glucosamine

32
Lipide A: variazioni nei gruppi fosforici e nel pattern di acilazione
Acido galatturonico b-idrossibutirrato al C28

34
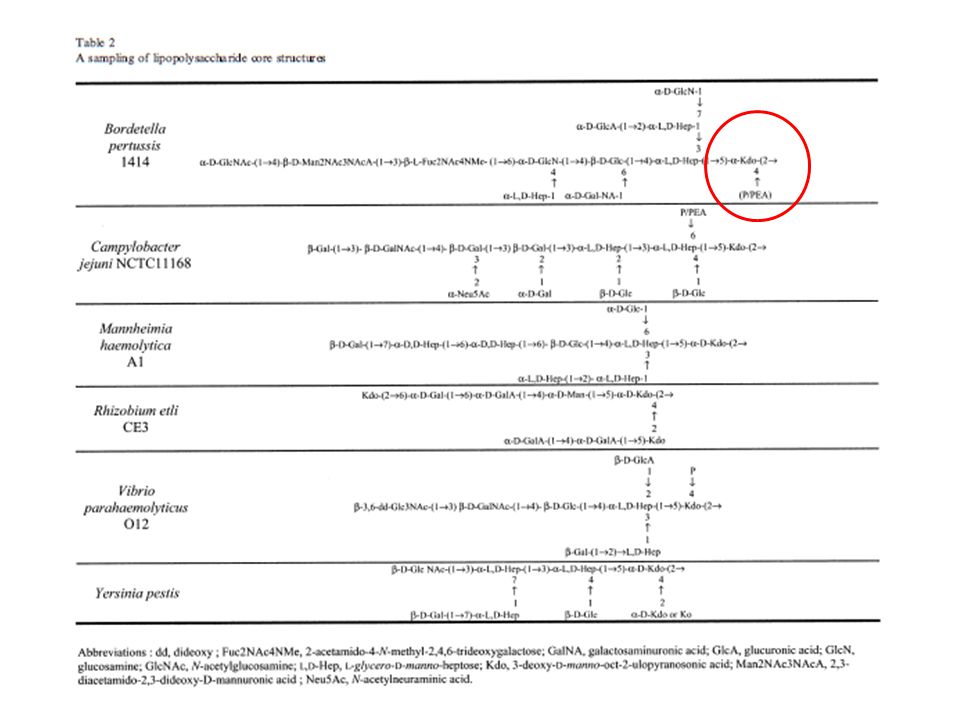
La regione del “Core”[1]
Sulla base della composizione in zuccheri (monosaccaridi) nelle Enterobacteriaceae si distinguono: CORE ESTERNO CORE INTERNO 2. Core esterno: consiste prevalentemente di ESOSI (D-glucosio, D-galattosio, D-glucosamina, N-acetilglucosamina, N-acetilgalattosamina) Il numero delle unità può variare da 8 a 12 3. Core interno: presenza di un caratteristico monosaccaride, il Kdo (acido 3-deossi-D-manno-octolusonico) Gli eptosi del core interno possono essere sostituiti con gruppi fosforici, pirofosforici, e fosforiletanolamina (Salmonella spp., Shigella, E. coli)
![La regione del Core [1]](http://slideplayer.it/slide/2327244/8/images/34/La+regione+del+Core+%5B1%5D.jpg "Sulla base della composizione in zuccheri (monosaccaridi) nelle Enterobacteriaceae si distinguono: CORE ESTERNO. CORE INTERNO. 2. Core esterno: consiste prevalentemente di ESOSI (D-glucosio, D-galattosio, D-glucosamina, N-acetilglucosamina, N-acetilgalattosamina) Il numero delle unità può variare da 8 a Core interno: presenza di un caratteristico monosaccaride, il Kdo (acido 3-deossi-D-manno-octolusonico) Gli eptosi del core interno possono essere sostituiti con gruppi fosforici, pirofosforici, e fosforiletanolamina (Salmonella spp., Shigella, E. coli)")
36
La regione del “Core” [2]
Alcune eccezioni - Francisella tularensis Manca completamente degli eptosi del core esterno; il solo residuo di Kdo presente è legato a un disaccaride di mannosio [b-mannosio (1–4) –a mannosio] - Legionella pneumophila Come per F. tularensis non vi sono eptosi ma residui di mannosio ….e ancora Acinetobacter, Chlamydia, Moraxella, Rhizobium La regione minima dell’LPS richiesta in E. coli è costituita dal lipide A e da due unità di Kdo
![La regione del Core [2]](http://slideplayer.it/slide/2327244/8/images/36/La+regione+del+Core+%5B2%5D.jpg "Alcune eccezioni. - Francisella tularensis. Manca completamente degli eptosi del core esterno; il solo residuo di Kdo presente è legato a un disaccaride di mannosio [b-mannosio (1–4) –a mannosio] - Legionella pneumophila. Come per F. tularensis non vi sono eptosi ma residui di mannosio. ….e ancora Acinetobacter, Chlamydia, Moraxella, Rhizobium. La regione minima dell’LPS richiesta in E. coli è costituita dal lipide A e da due unità di Kdo.")
37
La regione del “Core” [3]
Alcune eccezioni Alcune specie batteriche, Acinetobacter haemolyticus, B. cepacia, Y. Pestis, incorporano analoghi del Kdo, l’acido D-glicero-D-talo-octulosonico (Ko). La presenza del Kdo e dei residui di eptosio è stata (e tuttora è in uso) considerata un marker di identificazione dell’LPS. Nella clinica rappresenta un importante marker diagnostico per il rilevamento delle infezioni da parte di batteri Gram negativi. Patogeni non enterici, quali N. meningiditis, mancano dei residui di eptosio e incorporano il Ko in sostituzione del Kdo.
![La regione del Core [3]](http://slideplayer.it/slide/2327244/8/images/37/La+regione+del+Core+%5B3%5D.jpg "Alcune eccezioni. Alcune specie batteriche, Acinetobacter haemolyticus, B. cepacia, Y. Pestis, incorporano analoghi del Kdo, l’acido D-glicero-D-talo-octulosonico (Ko). La presenza del Kdo e dei residui di eptosio è stata (e tuttora è in uso) considerata un marker di identificazione dell’LPS. Nella clinica rappresenta un importante marker diagnostico per il rilevamento delle infezioni da parte di batteri Gram negativi. Patogeni non enterici, quali N. meningiditis, mancano dei residui di eptosio e incorporano il Ko in sostituzione del Kdo.")
38
La regione del “Core” [4]
Eterogeneità della composizione del core Il grado di diversità osservato nella regione del core dell’LPS dipende in buona parte da due fattori: 1. Presenza di sostituenti in rapporto non stechiometrico 2. Meccanismi genetici, primo fra tutti la variazione di fase Variazione di fase: scivolamento della polimerasi, durante il processo della replicazione in piccole sequenze nucleotidiche ripetute, all’interno di geni codificanti gli enzimi del macchinario biosintetico dell’LPS. H. influenzae, Neisseria spp., Campylobacter In N. meningiditis aggiunta di gruppi di acido sialico che correla con una più spiccata invasività e persistenza nei tessuti dell’ospite.
![La regione del Core [4]](http://slideplayer.it/slide/2327244/8/images/38/La+regione+del+Core+%5B4%5D.jpg "Eterogeneità della composizione del core. Il grado di diversità osservato nella regione del core dell’LPS dipende in buona parte da due fattori: 1. Presenza di sostituenti in rapporto non stechiometrico. 2. Meccanismi genetici, primo fra tutti la variazione di fase. Variazione di fase: scivolamento della polimerasi, durante il processo della replicazione in piccole sequenze nucleotidiche ripetute, all’interno di geni codificanti gli enzimi del macchinario biosintetico dell’LPS. H. influenzae, Neisseria spp., Campylobacter. In N. meningiditis aggiunta di gruppi di acido sialico che correla con una più spiccata invasività e persistenza nei tessuti dell’ospite.")
39
L’antigene somatico o Antigene-O [1]
1. L’antigene somatico è la regione dell’LPS che entra a diretto contatto con le membrane dell’ospite 2. Regione fortemente antigenica; la caratterizzazione della composizione in zuccheri rappresenta la base per una classificazione sierologica dei diversi sierotipi 3. Rappresenta una sorta di fingerprint del batterio. 4. Queste catene saccaridiche aiutano il batterio a sfuggire all’azione litica da parte delle molecole del complemento, in un processo che viene definito “shielding”. 5. Proteggono la cellula batterica dall’azione di numerosi antibiotici 6. Possono funzionare anche come importanti sistemi di adesione (Acinetobacillus pleuropneumoniae).
![L’antigene somatico o Antigene-O [1]](http://slideplayer.it/slide/2327244/8/images/39/L%E2%80%99antigene+somatico+o+Antigene-O+%5B1%5D.jpg "1. L’antigene somatico è la regione dell’LPS che entra a diretto contatto con le membrane dell’ospite. 2. Regione fortemente antigenica; la caratterizzazione della composizione in zuccheri rappresenta la base per una classificazione sierologica dei diversi sierotipi. 3. Rappresenta una sorta di fingerprint del batterio. 4. Queste catene saccaridiche aiutano il batterio a sfuggire all’azione litica da parte delle molecole del complemento, in un processo che viene definito shielding . 5. Proteggono la cellula batterica dall’azione di numerosi antibiotici. 6. Possono funzionare anche come importanti sistemi di adesione (Acinetobacillus pleuropneumoniae).")
40
L’antigene somatico o Antigene-O [2]
50 unità oligosaccaridiche ripetute costituite da 2-8 monosaccaridi variabili Si conoscono almeno 20 residui saccaridici che possono entrare nella composizione dell’O-chain, molti dei quali presentano caratteristiche uniche (dideossi-esosi): colitosio, paratosio… alcuni sierotipi di E. coli (O8 e O9), di K. pneumoniae (O3 e O5) e L. pneumophila contengono omopolimeri (mannosio) Estrema variabilità nella composizione anche all’interno della stessa specie La sintesi dell’antigene-O è determinata dal cluster genico wb (rfb) Mutanti difettivi nel cluster wb o mutanti di delezione Dwb presentano una morfologia diversa: R-mutants (R: rought) batteri wild-type: S (S: smooth)
![L’antigene somatico o Antigene-O [2]](http://slideplayer.it/slide/2327244/8/images/40/L%E2%80%99antigene+somatico+o+Antigene-O+%5B2%5D.jpg "50 unità oligosaccaridiche ripetute costituite da 2-8 monosaccaridi variabili. Si conoscono almeno 20 residui saccaridici che possono entrare nella composizione dell’O-chain, molti dei quali presentano caratteristiche uniche (dideossi-esosi): colitosio, paratosio… alcuni sierotipi di E. coli (O8 e O9), di K. pneumoniae (O3 e O5) e L. pneumophila contengono omopolimeri (mannosio) Estrema variabilità nella composizione anche all’interno della stessa specie. La sintesi dell’antigene-O è determinata dal cluster genico wb (rfb) Mutanti difettivi nel cluster wb o mutanti di delezione Dwb presentano una morfologia diversa: R-mutants (R: rought) batteri wild-type: S (S: smooth)")
41
L’antigene somatico o Antigene-O [3]
I mutanti R sono in grado di crescere in brodocoltura La porzione dell’antigene-O è dispensabile per la crescita batterica Nel caso delle specie patogene (Salmonella, Shigella, E. coli EPEC) la mancanza della regione somatica rende i batteri più suscettibile alla fagocitosi e all’attacco delle proteine del complemento. Patogeni batterici come N. meningitidis, H. influenzae, B. pertussis, H. influenzae, e C. trachomatis mancano dell’antigene-O (LPS a basso peso molecolare o lipooligosaccaridi [LOS])
![L’antigene somatico o Antigene-O [3]](http://slideplayer.it/slide/2327244/8/images/41/L%E2%80%99antigene+somatico+o+Antigene-O+%5B3%5D.jpg "I mutanti R sono in grado di crescere in brodocoltura. La porzione dell’antigene-O è dispensabile per la crescita batterica. Nel caso delle specie patogene (Salmonella, Shigella, E. coli EPEC) la mancanza della regione somatica rende i batteri più suscettibile alla fagocitosi e all’attacco delle proteine del complemento. Patogeni batterici come N. meningitidis, H. influenzae, B. pertussis, H. influenzae, e C. trachomatis mancano dell’antigene-O (LPS a basso peso molecolare o lipooligosaccaridi [LOS])")
42
L’antigene somatico o Antigene-O [4]
Lo scheletro dell’antigene somatico può presentarsi in forma lineare o ramificata, dipendente dalla presenza di gruppi sostituenti. I più comuni sostituenti sono: gruppi fosfato e fosforil-etanolammina; gruppi acetili. Meno frequenti: gruppi formilici, e acido glicerico. Durante il pathway biosisntetico le singole unità saccaridiche vengono polimerizzate in blocchi e aggiunte alla regione del core. Le molecole di LPS sono dunque formate da catene saccaridiche di diverse lunghezza (elettrofresi SDS).
![L’antigene somatico o Antigene-O [4]](http://slideplayer.it/slide/2327244/8/images/42/L%E2%80%99antigene+somatico+o+Antigene-O+%5B4%5D.jpg "Lo scheletro dell’antigene somatico può presentarsi in forma lineare o ramificata, dipendente dalla presenza di gruppi sostituenti. I più comuni sostituenti sono: gruppi fosfato e fosforil-etanolammina; gruppi acetili. Meno frequenti: gruppi formilici, e acido glicerico. Durante il pathway biosisntetico le singole unità saccaridiche vengono polimerizzate in blocchi e aggiunte alla regione del core. Le molecole di LPS sono dunque formate da catene saccaridiche di diverse lunghezza (elettrofresi SDS).")
43
Funzioni dell’LPS Nel caso delle specie patogene (Salmonella, Shigella, E. coli EPEC) l’LPS è fondamentale per l’interazione ospite-patogeno Difesa dall’attacco del complemento: a seconda della lunghezza delle catene polisaccaridiche l’LPS può inibire la formazione del MAC e quindi la lisi batterica Consente una maggiore protezione dal processo di fagocitosi Mimetismo molecolare: H. pylori (composizione dell’LPS che ricalca gli acidi di Lewis) Campilobacter jejuni (simile ai gangliosidi delle cellule eucariotiche); disordini autoimmunitari neurodegenerativi, sindrome di Guillan Barré Neisseria meningiditis aggiunta di residui di acido sialico ai LOS con conseguente inattivazione del complemento
 Campilobacter jejuni (simile ai gangliosidi delle cellule eucariotiche); disordini autoimmunitari neurodegenerativi, sindrome di Guillan Barré. Neisseria meningiditis aggiunta di residui di acido sialico ai LOS con conseguente inattivazione del complemento.")
44
La diversa struttura tridimensionale influenza il potenziale immunogenico
Struttura conica struttura cilindrica Agonista TLR4 Antagonista TLR4

45
Lipide A: struttura delle Enterobacteriaceae
Legame a-glicosidico estere Scheletro di D-glucosamine lo scheletro è costituito da un disaccaride con legame b,1-6 sono presenti due gruppi fosforici (solitamente il Lipide A è monofosforilato) sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi Posizione di attacco della regione polisaccaridica
 sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi. Posizione di attacco della regione polisaccaridica.")
46
Francisella tularensis
F. tularensis è un cocco bacillo Gram negativo, non mobile, aerobio Agente eziologico della tularemia: infezione/infiammazione della vie aeree che può evolvere verso broncopolmoniti. Patogeno umano e animale. La trasmissione avviene attraverso il morso degli artropodi, il contatto con tessuti animali infetti, via aerosol, acqua e cibo contaminati. Diffusione dei batteri a livello dei linfonodi e disseminazione per via ematica ai polmoni, pleura, milza, fegato e reni.

47
Modificazioni del Lipide A: il caso di Francisella tularensis [1]
F. tularensis sintetizza quantità relativamente scarse di LPS l’LPS di F. tularensis contiene quantità notevoli di lipide A libero Mancanza del Kdo Mancanza di molte specie di zuccheri nella regione del core interno Quale sia il significato biologico delle modificazioni del lipide A di F. tularensis non è noto. Nel genoma sono infatti presenti tutti gli enzimi per la biosintesi del Kdo e per l’aggiunta degli zuccheri del core. Possibile esistenza di un’idrolasi di membrana ?
![Modificazioni del Lipide A: il caso di Francisella tularensis [1]](http://slideplayer.it/slide/2327244/8/images/47/Modificazioni+del+Lipide+A%3A+il+caso+di+Francisella+tularensis+%5B1%5D.jpg "F. tularensis sintetizza quantità relativamente scarse di LPS. l’LPS di F. tularensis contiene quantità notevoli di lipide A libero. Mancanza del Kdo. Mancanza di molte specie di zuccheri nella regione del core interno. Quale sia il significato biologico delle modificazioni del lipide A di F. tularensis non è noto. Nel genoma sono infatti presenti tutti gli enzimi per la biosintesi del Kdo e per l’aggiunta degli zuccheri del core. Possibile esistenza di un’idrolasi di membrana")
48
Modificazioni del Lipide A: il caso di Francisella tularensis [2]
![Modificazioni del Lipide A: il caso di Francisella tularensis [2]](http://slideplayer.it/slide/2327244/8/images/48/Modificazioni+del+Lipide+A%3A+il+caso+di+Francisella+tularensis+%5B2%5D.jpg "Modificazioni del Lipide A: il caso di Francisella tularensis [2]")
49
Modificazioni del Lipide A: il caso di Francisella tularensis [3]
Quali sono le reazioni enzimatiche responsabili di queste modifiche del lipide A? Presenza di due specifiche fosfatasi di membrana: LpxE LpxF Presenza di una trasferasi: ArnT (PmrK) In F. tularensis catalizza l’aggiunta del residuo di galattosamina al gruppo fosfato C1. Lo stesso enzima funziona anche nel pathway biosintetico dell’LPS delle Enterobacteriaceae. Ceppi di F. novocida che mancano della fosfatasi LpxF mostrano un fenotipo attenuato nei modelli murini di infezione. Maggiore suscettibilità a peptidi cationici antimicrobici Ceppi di Salmonella che esprimono la fosfatasi LpxE presentano un Lipide A monofosforilato (monofosforil Lipide A). Importanti nell’ambito clinico come adiuvanti vaccinali e per lo sviluppo di vaccini orali.
![Modificazioni del Lipide A: il caso di Francisella tularensis [3]](http://slideplayer.it/slide/2327244/8/images/49/Modificazioni+del+Lipide+A%3A+il+caso+di+Francisella+tularensis+%5B3%5D.jpg "Quali sono le reazioni enzimatiche responsabili di queste modifiche del lipide A Presenza di due specifiche fosfatasi di membrana: LpxE. LpxF. Presenza di una trasferasi: ArnT (PmrK) In F. tularensis catalizza l’aggiunta del residuo di galattosamina al gruppo fosfato C1. Lo stesso enzima funziona anche nel pathway biosintetico dell’LPS delle Enterobacteriaceae. Ceppi di F. novocida che mancano della fosfatasi LpxF mostrano un fenotipo attenuato nei modelli murini di infezione. Maggiore suscettibilità a peptidi cationici antimicrobici. Ceppi di Salmonella che esprimono la fosfatasi LpxE presentano un Lipide A monofosforilato (monofosforil Lipide A). Importanti nell’ambito clinico come adiuvanti vaccinali e per lo sviluppo di vaccini orali.")
50
Modificazioni del Lipide A: il caso delle Enterobacteriaceae [1]
Legame a-glicosidico estere Scheletro di D-glucosamine
![Modificazioni del Lipide A: il caso delle Enterobacteriaceae [1]](http://slideplayer.it/slide/2327244/8/images/50/Modificazioni+del+Lipide+A%3A+il+caso+delle+Enterobacteriaceae+%5B1%5D.jpg "Legame. a-glicosidico. estere. Scheletro di D-glucosamine.")
51
Modificazioni del Lipide A: il caso delle Enterobacteriaceae [2]
E coli K12 e S. typhimurium codificano numerosi enzimi responsabili delle modificazioni del Lipide A Ma quali sono queste modifiche? Sostituzione dei gruppi fosfato al C1 e al C4’ con gruppi fosfoetanolamminici Sostituzione e aggiunta al C4’ di un residuo di 4 ammino-arabinosio (L-Ara4N) Modifiche nelle catene aciliche primarie, incorporazione di palmitato Idrossilazione delle catene aciliche primarie Modifiche delle catene aciliche secondarie Molte delle modifiche osservate nella composizione chimica del lipide A rispondono a cambiamenti delle condizioni di crescita
![Modificazioni del Lipide A: il caso delle Enterobacteriaceae [2]](http://slideplayer.it/slide/2327244/8/images/51/Modificazioni+del+Lipide+A%3A+il+caso+delle+Enterobacteriaceae+%5B2%5D.jpg "E coli K12 e S. typhimurium codificano numerosi enzimi responsabili delle modificazioni del Lipide A. Ma quali sono queste modifiche Sostituzione dei gruppi fosfato al C1 e al C4’ con gruppi fosfoetanolamminici. Sostituzione e aggiunta al C4’ di un residuo di 4 ammino-arabinosio (L-Ara4N) Modifiche nelle catene aciliche primarie, incorporazione di palmitato. Idrossilazione delle catene aciliche primarie. Modifiche delle catene aciliche secondarie. Molte delle modifiche osservate nella composizione chimica del lipide A rispondono a cambiamenti delle condizioni di crescita.")
52
Modificazioni del Lipide A: il caso delle Enterobacteriaceae [3]
C14 acido miristico C16 acido palmitico
![Modificazioni del Lipide A: il caso delle Enterobacteriaceae [3]](http://slideplayer.it/slide/2327244/8/images/52/Modificazioni+del+Lipide+A%3A+il+caso+delle+Enterobacteriaceae+%5B3%5D.jpg "C14 acido miristico. C16 acido palmitico.")
53
Modificazioni del Lipide A: il caso di Salmonella [1]
Il sistema meglio conosciuto è quello PagP/PagL PagP: Lipide A palmitoleil transferasi PagL: Lipide A deacilasi (lipasi della membrana esterna) Il sistema PagP/PagL permette la rimozione della catena acilica di idrossimiristato (PagL) dal C3e conseguentemente l’aggiunta di palmitato (PagP) Le modificazioni del pattern di acilazione rispondono al sistema a due componenti PhoP/PhoQ PagL 3-O-deacilasi Mutanti di S. typhimurium incapaci di aggiungere palmitato al Lipide A diventano più suscettibili all’azione di alcuni peptidi cationici con attività antibatterica ma non alle polimixine.
![Modificazioni del Lipide A: il caso di Salmonella [1]](http://slideplayer.it/slide/2327244/8/images/53/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B1%5D.jpg "Il sistema meglio conosciuto è quello PagP/PagL. PagP: Lipide A palmitoleil transferasi. PagL: Lipide A deacilasi (lipasi della membrana esterna) Il sistema PagP/PagL permette la rimozione della catena acilica di idrossimiristato (PagL) dal C3e conseguentemente l’aggiunta di palmitato (PagP) Le modificazioni del pattern di acilazione rispondono al sistema a due componenti PhoP/PhoQ. PagL. 3-O-deacilasi. Mutanti di S. typhimurium incapaci di aggiungere palmitato al Lipide A diventano più suscettibili all’azione di alcuni peptidi cationici con attività antibatterica ma non alle polimixine.")
54
Modificazioni del Lipide A: il caso di Salmonella [2] sistemi di percezione sensoriale a due componenti Caratterizazione in seguito all’individuazione di mutazioni pleiotropiche in E. coli, Salmonella. Due componenti: Sensore (proteina di membrana) Regolatore di risposta (citosolico) Sensore: dominio sensoriale e un dominio del trasmettitore Regolatore: Dominio di ricezione e un dominio di legame al DNA PERIPLASMA NH-2 COOH CITOPLASMA NH-2 COOH
![Modificazioni del Lipide A: il caso di Salmonella [2] sistemi di percezione sensoriale a due componenti](http://slideplayer.it/slide/2327244/8/images/54/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B2%5D+sistemi+di+percezione+sensoriale+a+due+componenti.jpg "Caratterizazione in seguito all’individuazione di mutazioni pleiotropiche in E. coli, Salmonella. Due componenti: Sensore (proteina di membrana) Regolatore di risposta (citosolico) Sensore: dominio sensoriale e un dominio del trasmettitore. Regolatore: Dominio di ricezione e un dominio di legame al DNA. PERIPLASMA. NH-2. COOH. CITOPLASMA. NH-2. COOH.")
55
Modificazioni del Lipide A: il caso di Salmonella [3] Il sistema PhoP/PhoQ
PhoP: attivatore/repressore trascrizionale PhoQ: protein chinasi Il complesso PhoP/PhoQ può regolare (attivare o reprimere) simultaneamente più di 40 geni differenti Fattore ambientale Percezione del segnale Informazione di tipo trascrizionale al genoma In particolare: Idrossilazione in posizione 2 dell’acido miristico Basse concentrazioni di Mg++ Attivazione del sistema PhoP/PhoQ Modificazioni strutturali del Lipide A Attivazione del sistema PagP/PagL
![Modificazioni del Lipide A: il caso di Salmonella [3] Il sistema PhoP/PhoQ](http://slideplayer.it/slide/2327244/8/images/55/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B3%5D+Il+sistema+PhoP%2FPhoQ.jpg "PhoP: attivatore/repressore trascrizionale. PhoQ: protein chinasi. Il complesso PhoP/PhoQ può regolare (attivare o reprimere) simultaneamente più di 40 geni differenti. Fattore ambientale. Percezione del segnale. Informazione di tipo trascrizionale al genoma. In particolare: Idrossilazione in posizione 2 dell’acido miristico. Basse concentrazioni di Mg++ Attivazione del sistema PhoP/PhoQ. Modificazioni strutturali del Lipide A. Attivazione del sistema PagP/PagL.")
56
Modificazioni del Lipide A: il caso di Salmonella [4]
Il sistema PhoP/PhoQ Il sistema PmrA/PmrB
![Modificazioni del Lipide A: il caso di Salmonella [4]](http://slideplayer.it/slide/2327244/8/images/56/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B4%5D.jpg "Il sistema PhoP/PhoQ. Il sistema PmrA/PmrB.")
57
Modificazioni del Lipide A: il caso di Salmonella [4]
Il sistema PhoP/PhoQ Il sistema PmrA/PmrB
![Modificazioni del Lipide A: il caso di Salmonella [4]](http://slideplayer.it/slide/2327244/8/images/57/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B4%5D.jpg "Il sistema PhoP/PhoQ. Il sistema PmrA/PmrB.")
58
Modificazioni del Lipide A: il caso delle Enterobacteriaceae [3]
C14 acido miristico C16 acido palmitico
![Modificazioni del Lipide A: il caso delle Enterobacteriaceae [3]](http://slideplayer.it/slide/2327244/8/images/58/Modificazioni+del+Lipide+A%3A+il+caso+delle+Enterobacteriaceae+%5B3%5D.jpg "C14 acido miristico. C16 acido palmitico.")
59
Modificazioni del Lipide A: il caso delle Enterobacteriaceae [4]
Aggiunta dell’L-Ara4N Il residuo di L-Ara4N è caricato positivamente e neutralizza la carica negativa dei gruppi fosforici presenti al C1. Questa modifica del Lipide A riduce la sensibilità dei batteri a diverse classi di peptidi cationici ad attività antibatterica (defensine e catelecidine) e alle polimixine. Aggiunta di due gruppi fosfoetanolammina Due sono gli enzimi coivolti in questo tipo di modifica del Lipide A: EptA e EptB. EptA catalizza l’aggiunta della fosfoetanolammina prevalentemente al C1. In particolari condizioni di crescita o in condizioni limitanti di arabinosio la modifica del gruppo fosforico può interessare anche il C4’. Ancora non si conosce il motivo di questi cambiamenti.
![Modificazioni del Lipide A: il caso delle Enterobacteriaceae [4]](http://slideplayer.it/slide/2327244/8/images/59/Modificazioni+del+Lipide+A%3A+il+caso+delle+Enterobacteriaceae+%5B4%5D.jpg "Aggiunta dell’L-Ara4N. Il residuo di L-Ara4N è caricato positivamente e neutralizza la carica negativa dei gruppi. fosforici presenti al C1. Questa modifica del Lipide A riduce la sensibilità dei batteri a. diverse classi di peptidi cationici ad attività antibatterica (defensine e catelecidine) e. alle polimixine. Aggiunta di due gruppi fosfoetanolammina. Due sono gli enzimi coivolti in questo tipo di modifica del Lipide A: EptA e EptB. EptA. catalizza l’aggiunta della fosfoetanolammina prevalentemente al C1. In particolari. condizioni di crescita o in condizioni limitanti di arabinosio la modifica del gruppo. fosforico può interessare anche il C4’. Ancora non si conosce il motivo di questi. cambiamenti.")
60
Il sistema PagP/PagL L. pneumophila e B. bronchiseptica
PagP è essenziale per l’infezione degli animali. L’aggiunta di palmitato promuove la resistenza ai peptidi antimicrobici e alla lisi del complemento mediata da anticorpi. Y. pseudotubercolosis L’incorporazione di palmitato avviene in misura maggiore in risposta a variazioni della temperatura (all’interno dell’ospite) P. aeruginosa Modificazioni del lipide A oltre che per aggiunta di palmitato (PagP/PagL) anche per aggiunta di idrossi-laurato e amino-arabinosio
 P. aeruginosa. Modificazioni del lipide A oltre che per aggiunta di palmitato (PagP/PagL) anche per aggiunta di idrossi-laurato e amino-arabinosio.")
61
Modificazioni del Lipide A: il caso di Salmonella [4]
LpxR: lipasi della membrana esterna (deacilasi); è sotto il controllo del sistema PhoP/PhoQ LpxO: enzima della membrana interna che idrossila le catene aciliche secondarie Gli enzimi LpxR, LpxO e PagL non sono presenti in E. coli K12. Quando sovrespressi in E. coli K12 determinano le tipiche modifiche del Lipide A osservate in Salmonella.
![Modificazioni del Lipide A: il caso di Salmonella [4]](http://slideplayer.it/slide/2327244/8/images/61/Modificazioni+del+Lipide+A%3A+il+caso+di+Salmonella+%5B4%5D.jpg "LpxR: lipasi della membrana esterna (deacilasi); è sotto il controllo del sistema PhoP/PhoQ. LpxO: enzima della membrana interna che idrossila le catene aciliche secondarie. Gli enzimi LpxR, LpxO e PagL non sono presenti in E. coli K12. Quando sovrespressi in E. coli K12 determinano le tipiche modifiche del Lipide A osservate in Salmonella.")
62
Modificazioni dell’antigene-O [1]
50 unità oligosaccaridiche ripetute costituite da 2-8 monosaccaridi variabili Si conoscono almeno 20 residui saccaridici che possono entrare nella composizione dell’O-chain, molti dei quali presentano caratteristiche uniche (dideossi-esosi): colitosio, paratosio. Molecole eteropolimeriche (o omopolimeriche) La regione dell’antigene-O dell’LPS è quella sottoposta a maggiori modificazioni Salmonella: 1000 varianti chimicamente e strutturalmente diverse E coli: esistenza di 173 diverse molecole di antigene-O Le molecole di LPS estratte da una singola popolazione batterica sono spesso eterogenee nella struttura: esistenza di un ampio spettro di glicoforme. Le alterazioni nella struttura dell’LPS avvengono in risposta a variazioni delle condizioni ambientali e nel caso dei patogeni in risposta non solo all’ambiente dell’ospite ma anche ai diversi step della patogenesi.
![Modificazioni dell’antigene-O [1]](http://slideplayer.it/slide/2327244/8/images/62/Modificazioni+dell%E2%80%99antigene-O+%5B1%5D.jpg "50 unità oligosaccaridiche ripetute costituite da 2-8 monosaccaridi variabili. Si conoscono almeno 20 residui saccaridici che possono entrare nella composizione dell’O-chain, molti dei quali presentano caratteristiche uniche (dideossi-esosi): colitosio, paratosio. Molecole eteropolimeriche (o omopolimeriche) La regione dell’antigene-O dell’LPS è quella sottoposta a maggiori modificazioni. Salmonella: 1000 varianti chimicamente e strutturalmente diverse. E coli: esistenza di 173 diverse molecole di antigene-O. Le molecole di LPS estratte da una singola popolazione batterica sono spesso eterogenee nella struttura: esistenza di un ampio spettro di glicoforme. Le alterazioni nella struttura dell’LPS avvengono in risposta a variazioni delle condizioni ambientali e nel caso dei patogeni in risposta non solo all’ambiente dell’ospite ma anche ai diversi step della patogenesi.")
63
Modificazioni dell’antigene-O [2]
Che cosa intendiamo per variazioni dell’antigene-O e a che livello si realizzano? La composizione chimica dell’antigene-O La composizione in zuccheri: il tipo e il numero di monosaccaridi che entrano nella composizione La struttura dell’antigene-O Diversità nel tipo di legame glicosidico (cambiamento posizionale e anomerico) Configurazione degli zuccheri Presenza di gruppi sostituenti Lunghezza delle catene dell’antigene-O
![Modificazioni dell’antigene-O [2]](http://slideplayer.it/slide/2327244/8/images/63/Modificazioni+dell%E2%80%99antigene-O+%5B2%5D.jpg "Che cosa intendiamo per variazioni dell’antigene-O e a che livello si realizzano La composizione chimica dell’antigene-O. La composizione in zuccheri: il tipo e il numero di monosaccaridi che entrano nella. composizione. La struttura dell’antigene-O. Diversità nel tipo di legame glicosidico (cambiamento posizionale e anomerico) Configurazione degli zuccheri. Presenza di gruppi sostituenti. Lunghezza delle catene dell’antigene-O.")
64
Modificazioni dell’antigene-O [3]
Quali sono le principali variazioni in termini di gruppi sostituenti? Gruppi saccaridici: fucosilazione e glicosilazione Gruppi non saccaridici: metilici e acetilici Generalmente le variazioni non avvengono in maniera stechiometrica! Quali sono i meccanismi attraverso i quali i batteri modificano l’antigene-O? Riprogrammazione dell’espressione genica Conversione lisogenica (mediata da fagi) Trasferimento genico orizzontale (HGT) anche noto come trasferimento genico laterale (LGT) seguito da eventi di ricombinazione
![Modificazioni dell’antigene-O [3]](http://slideplayer.it/slide/2327244/8/images/64/Modificazioni+dell%E2%80%99antigene-O+%5B3%5D.jpg "Quali sono le principali variazioni in termini di gruppi sostituenti Gruppi saccaridici: fucosilazione e glicosilazione. Gruppi non saccaridici: metilici e acetilici. Generalmente le variazioni non avvengono in maniera stechiometrica! Quali sono i meccanismi attraverso i quali i batteri modificano l’antigene-O Riprogrammazione dell’espressione genica. Conversione lisogenica (mediata da fagi) Trasferimento genico orizzontale (HGT) anche noto come trasferimento genico laterale (LGT) seguito da eventi di ricombinazione.")
65
Modificazioni dell’antigene-O [4] I meccanismi del trasferimento genico orizzontale (HGT)
Comparazione delle sequenze geniche che codificano gli enzimi del pathway biosintetico dell’antigene-O Microrganismi modello: S. enterica e E. coli Gene A Gene B Gene C Gene D Gene E Gene F Cromosoma Regione variabile Regione costante Cromosoma Organizzazione in cluster Il numero dei geni varia da un minimo di 6 unità fino anche a 9 per ciascun cluster (complessità della catena dell’antigene-O) Contenuto in G+C diverso rispetto agli altri geni cromosomiali (generalmente inferiore al 40%) All’interno del cluster il contenuto in G+C dei geni è diverso Assemblaggio dei geni in un cluster (come i tasselli di un puzzle) Presenza di regioni più o meno conservate e di regioni variabili che possono favorire eventi di excisione e di ricombinazione Acquisizione del materiale genetico attraverso plasmidi Ci sono batteri, H. pylori, nei quali i geni del pathway biosisntetico dell’antigene-O non sono organizzati in cluster ma interspersi nel genoma
![Modificazioni dell’antigene-O [4] I meccanismi del trasferimento genico orizzontale (HGT)](http://slideplayer.it/slide/2327244/8/images/65/Modificazioni+dell%E2%80%99antigene-O+%5B4%5D+I+meccanismi+del+trasferimento+genico+orizzontale+%28HGT%29.jpg "Comparazione delle sequenze geniche che codificano gli enzimi del pathway biosintetico dell’antigene-O. Microrganismi modello: S. enterica e E. coli. Gene A. Gene B. Gene C. Gene D. Gene E. Gene F. Cromosoma. Regione variabile. Regione costante. Cromosoma. Organizzazione in cluster. Il numero dei geni varia da un minimo di 6 unità fino anche a 9 per ciascun cluster (complessità della catena dell’antigene-O) Contenuto in G+C diverso rispetto agli altri geni cromosomiali (generalmente inferiore al 40%) All’interno del cluster il contenuto in G+C dei geni è diverso. Assemblaggio dei geni in un cluster (come i tasselli di un puzzle) Presenza di regioni più o meno conservate e di regioni variabili che possono favorire eventi di excisione e di ricombinazione. Acquisizione del materiale genetico attraverso plasmidi. Ci sono batteri, H. pylori, nei quali i geni del pathway biosisntetico dell’antigene-O non sono organizzati in cluster ma interspersi nel genoma.")
66
Modificazioni dell’antigene-O [5] La conversione lisogenica
Un altro modo che i batteri sfruttano per acquisire nuovo materiale genetico è il processo della trasduzione, mediata dai fagi. Alcune proteine che vengono espresse dal fago, durante il suo ciclo lisogenico, entrano a far parte della parete cellulare batterica. In alcuni casi queste nuove proteine possono conferire fenotipi di virulenza Geni lom del fago l che codificano per fattori di adesione di alcuni ceppi di E. coli alle cellule dell’epitelio orale) Geni bor conferiscono una più spiccata resistenza degli E. coli nel siero L’integrazione di un fago nel genoma batterico può portare alla conversione sierotipica Conversione sierotipica Aggiunta di gruppi glucosidici o O-acetilici sugli zuccheri dell’antigene-O grazie ad enzimi espressi specificatamente dal pro-fago con conseguente shift di un batterio da un sierotipo a un altro. Quello della conversione sierotipica è un meccanismo di protezione del pro-fago.
![Modificazioni dell’antigene-O [5] La conversione lisogenica](http://slideplayer.it/slide/2327244/8/images/66/Modificazioni+dell%E2%80%99antigene-O+%5B5%5D+La+conversione+lisogenica.jpg "Un altro modo che i batteri sfruttano per acquisire nuovo materiale genetico è il processo della trasduzione, mediata dai fagi. Alcune proteine che vengono espresse dal fago, durante il suo ciclo lisogenico, entrano a far parte della parete cellulare batterica. In alcuni casi queste nuove proteine possono conferire fenotipi di virulenza. Geni lom del fago l che codificano per fattori di adesione di alcuni ceppi di E. coli alle cellule dell’epitelio orale) Geni bor conferiscono una più spiccata resistenza degli E. coli nel siero. L’integrazione di un fago nel genoma batterico può portare alla conversione sierotipica. Conversione sierotipica. Aggiunta di gruppi glucosidici o O-acetilici sugli zuccheri dell’antigene-O grazie ad enzimi. espressi specificatamente dal pro-fago con conseguente shift di un batterio da un sierotipo a. un altro. Quello della conversione sierotipica è un meccanismo di protezione del pro-fago.")
67
La conversione lisogenica: il caso di Shigella [1]
Agente eziologico Shigella spp. S. flexneri, S. boydii, S. sonnei, S. dysenteriae 50 seriotipi Sintomi Febbre, crampi addominali Sangue e muco nelle feci Sito d’infezione Epitelio del colon e del retto Trasmissione/Dose infettiva Acqua e cibo contaminati 10 – 100 batteri Dati epidemiologici 170 milioni di casi l’anno 1 milione di morti prevalentemente nei Paesi in via di sviluppo
![La conversione lisogenica: il caso di Shigella [1]](http://slideplayer.it/slide/2327244/8/images/67/La+conversione+lisogenica%3A+il+caso+di+Shigella+%5B1%5D.jpg "Agente eziologico. Shigella spp. S. flexneri, S. boydii, S. sonnei, S. dysenteriae. 50 seriotipi. Sintomi. Febbre, crampi addominali. Sangue e muco nelle feci. Sito d’infezione. Epitelio del colon e del retto. Trasmissione/Dose infettiva. Acqua e cibo contaminati. 10 – 100 batteri. Dati epidemiologici. 170 milioni di casi l’anno. 1 milione di morti prevalentemente. nei Paesi in via di sviluppo.")
68
La conversione lisogenica: il caso di Shigella [2] La glicosilazione
In S. flexneri l’antigene-O è una catena tetrasaccaridica Il batteriofago X di S. flexneri (SfX) permette la conversione del sierotipo Y al sierotipo X Aggiunta di un residuo di glucosio al primo residuo di ramnosio Serotype X
![La conversione lisogenica: il caso di Shigella [2] La glicosilazione](http://slideplayer.it/slide/2327244/8/images/68/La+conversione+lisogenica%3A+il+caso+di+Shigella+%5B2%5D+La+glicosilazione.jpg "In S. flexneri l’antigene-O è una catena tetrasaccaridica. Il batteriofago X di S. flexneri (SfX) permette la conversione del sierotipo Y al sierotipo X. Aggiunta di un residuo di glucosio al primo residuo di ramnosio. Serotype X.")
69
La conversione lisogenica: il caso di Shigella [3]
Il batteriofago X di S. flexneri (SfX) permette la conversione del sierotipo Y al sierotipo X Aggiunta di un residuo di glucosio al primo residuo di ramnosio Sono note le proteine del batteriofago che intervengono nella modifica dell’antigene-O GtrX: glicosil trasferasi GtrA: piccola proteina altamente idrofobica responsabile del trasferimento del residuo di glucosio legato ad un lipide carrier attraverso la membrana GtrB: enzima che catalizza il trasferimento del residuo di glucosio dall’UDP-glucosio al lipide carrier Le modificazioni dell’antigene–O in seguito all’aggiunta di residui di glucosio possono interessare anche gli altri residui di ramnosio e anche l’GlcNac. Questo dipende dalla natura del batteriofago. Il risultato è l’insorgenza di nuovi sierotipi.
![La conversione lisogenica: il caso di Shigella [3]](http://slideplayer.it/slide/2327244/8/images/69/La+conversione+lisogenica%3A+il+caso+di+Shigella+%5B3%5D.jpg "Il batteriofago X di S. flexneri (SfX) permette la conversione del sierotipo Y al sierotipo X. Aggiunta di un residuo di glucosio al primo residuo di ramnosio. Sono note le proteine del batteriofago che intervengono nella modifica dell’antigene-O. GtrX: glicosil trasferasi. GtrA: piccola proteina altamente idrofobica responsabile del trasferimento del residuo di glucosio legato ad un lipide carrier attraverso la membrana. GtrB: enzima che catalizza il trasferimento del residuo di glucosio dall’UDP-glucosio al lipide carrier. Le modificazioni dell’antigene–O in seguito all’aggiunta di residui di glucosio possono interessare. anche gli altri residui di ramnosio e anche l’GlcNac. Questo dipende dalla natura del batteriofago. Il. risultato è l’insorgenza di nuovi sierotipi.")
70
Modificazioni dell’antigene-O [6] Riprogrammazione dell’espressione genica
L’espressione dei geni che regolano le modificazioni dell’antigene-O può essere regolata in risposta a variazioni dell’ambiente (temperatura, disponibilità di nutrienti, pH) Nel caso dei patogeni questi meccanismi possono operare in risposta anche ai diversi step della patogenesi. Possiamo riconoscere due diversi meccanismi: 1. Attivazione/Repressione finemente regolata in risposta agli stimoli esterni ( es. sistema sensoriale a due componenti) 2. Un meccanismo random di accensione e spegnimento di specifici geni: variazione di fase, anche nota come variazione antigenica La variazione di fase è un meccanismo che avviene spontaneamente, con un’elevata frequenza (0,5%). È un meccanismo switching on/of reversibile Il flagello in Salmonella Le fimbrie di tipi I e i pili Pap di E. coli i LOS di H. influenzae e N. meningiditis le proteine Opa di N. gonorrheae Quattro sono i principali cambiamenti nella struttura dell’antigene-O dovuti a eventi di variazione di fase: fucosilazione, glicosilazione, acetilazione e cambiamenti della lunghezza della catena saccaridica
![Modificazioni dell’antigene-O [6] Riprogrammazione dell’espressione genica](http://slideplayer.it/slide/2327244/8/images/70/Modificazioni+dell%E2%80%99antigene-O+%5B6%5D+Riprogrammazione+dell%E2%80%99espressione+genica.jpg "L’espressione dei geni che regolano le modificazioni dell’antigene-O può essere regolata in risposta a variazioni dell’ambiente (temperatura, disponibilità di nutrienti, pH) Nel caso dei patogeni questi meccanismi possono operare in risposta anche ai diversi step della patogenesi. Possiamo riconoscere due diversi meccanismi: 1. Attivazione/Repressione finemente regolata in risposta agli stimoli esterni ( es. sistema sensoriale a due componenti) 2. Un meccanismo random di accensione e spegnimento di specifici geni: variazione di fase, anche nota come variazione antigenica. La variazione di fase è un meccanismo che avviene spontaneamente, con un’elevata frequenza (0,5%). È un meccanismo switching on/of reversibile. Il flagello in Salmonella. Le fimbrie di tipi I e i pili Pap di E. coli. i LOS di H. influenzae e N. meningiditis. le proteine Opa di N. gonorrheae. Quattro sono i principali cambiamenti nella struttura dell’antigene-O dovuti a eventi di variazione di. fase: fucosilazione, glicosilazione, acetilazione e cambiamenti della lunghezza della catena. saccaridica.")
Presentazioni simili





















 Pontieri - Elementi di Patologia Generale (ed. Piccin) Majno-Joris - Cellule,>")
