Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Le biotecnologie innovative

2
Esperimento di Griffith (1928)
Conclusione: Esiste un fattore «trasformante» 2 3 4

3
Esperimento di Avery (1944)
Proteasi Lipasi DNAasi Amilasi Conclusione: Il fattore «trasformante» è il DNA

4
Esperimento di Hershey - Chase (1952)
Conclusione: Il DNA è «ereditario»

5
Genetica dei microbi A partire dagli anni ‘50 del secolo scorso, grazie soprattutto allo sviluppo e messa a punto di tecniche microscopiche radioattive, fosforescenti e fluorescenti, elettroforetiche ecc. la ricerca fece passi da gigante verso la scoperta dei processi biochimici più intimi delle cellule. Si riuscì, così, a conoscere: La struttura e i meccanismi di azione dei virus La struttura dei cromosomi (molecole di DNA contenente informazioni fondamentali per la vita degli organismi). La presenza nei batteri di un solo DNA (grande molecoa a struttrura circolare e sparsa nel citoplasma L’esistenza, nei batteri, e la struttura di altre 20 /30 piccole molecole circolari di DNA, indicate con il nome di plasmidi, contenenti informazioni per caratteri «accessori» (non fondamentali, comunque utili per il batterio stesso). I meccanismi di variabilità ed «evoluzione» batterica
. La presenza nei batteri di un solo DNA (grande molecoa a struttrura circolare e sparsa nel citoplasma. L’esistenza, nei batteri, e la struttura di altre 20 /30 piccole molecole circolari di DNA, indicate con il nome di plasmidi, contenenti informazioni per caratteri «accessori» (non fondamentali, comunque utili per il batterio stesso). I meccanismi di variabilità ed «evoluzione» batterica.")
6
Meccanismo di un virus a struttura globulare
Struttura dei virus Meccanismo di un virus a struttura globulare

7
Meccanismo di un virus a DNA con struttura complessa
Ciclo litico: il loro DNA si circolarizza all’interno del batterio divenendo simile ad un plasmidio si duplica e codifica per le altre componenti virali le varie componenti virali si assemblano formando nuovi virus che, in seguito a rottura, «lisi» del batterio, fuoriescono nell’ambiente. Ciclo lisogeno: Il DNA si integra nel cromosoma batterico (viene indicato con il nome di profago) si duplicherà normalmente e verrà trasmesso ai batteri «figli» Rimarrà inerte per diverso tempo e diverse divisioni batteriche fino a che, per cause varie si attiverà innescando un ciclo litico. profago Attivazione
 si duplicherà normalmente e verrà trasmesso ai batteri «figli» Rimarrà inerte per diverso tempo e diverse divisioni batteriche fino a che, per cause varie si attiverà innescando un ciclo litico. profago. Attivazione.")
8
I virus ad RNA possono aver uno dei seguenti meccanismi d’azione:
Ciclo della RNA-polimerasi: Se possiedono questo specifico complesso enzimatico, l’RNA virale verrà utilizzato come matrice per sintetizzare, a stampo, varie molecole di RNA messaggero virale che verranno utilizzate per «costruire» le varie componenti virali, ma anche come «stampo» per nuovo RNA virale. Il tutto verrà assemblato nei virus «figli». Ciclo della Trascrittasi inversa: Grazie a questo particolare complesso enzimatico, l’RNA virale farà da matrice per «stampare» catene di DNA virale (provirus) definito anche DNA complementare (cDNA). Tale molecola si integrerà stabilmente nel cromosoma batterico e, di tanto in tanto, si attiverà producendo materiale virale.
 definito anche DNA complementare (cDNA). Tale molecola si integrerà stabilmente nel cromosoma batterico e, di tanto in tanto, si attiverà producendo materiale virale.")
9
Ciclo della RNA-polimerasi

10
Ciclo della Trascrittasi inversa

11
La variabilità genetica nei batteri
Mutazione (Errori nella duplicazione del DNA) Ricombinazione endogena (crossing-over: tra parti del cromosoma, tra cromosoma e plasmidi, tra plasmidi)
 Ricombinazione endogena (crossing-over: tra parti del cromosoma, tra cromosoma e plasmidi, tra plasmidi)")
12
Trasformazione Trasduzione Coniugazione
Trasferimento genico (trasformazione, trasduzione, coniugazione) Trasformazione Trasduzione Coniugazione
 Trasformazione. Trasduzione. Coniugazione.")
13
Trasformazione Processo in cui il DNA presente all’esterno del batterio viene internalizzato nella cellula ospite, integrato nel cromosoma nella corrispondente regione omologa e stabilmente ereditato con le divisioni cellulari

14
Il DNA (cromosoma o plasmide) viene introdotto integro nella cellula
La trasformazione artificiale La maggior parte dei batteri possono essere trasformati artificialmente Vengono utilizzati shock chimici e termici (CaCl2) o elettrici (elettroporazione) Il DNA (cromosoma o plasmide) viene introdotto integro nella cellula DNA lineare a doppia elica DNA plasmidico integrazione mediante ricombinazione omologa replicazione del plasmide
 o elettrici (elettroporazione) Il DNA (cromosoma o plasmide) viene introdotto integro nella cellula. DNA lineare a doppia elica. DNA plasmidico. integrazione mediante ricombinazione omologa. replicazione del plasmide.")
15
Trasduzione Modalità si scambio genetico fra batteri mediato da un virus DNA batterico DNA virale

16
Trasduzione generalizzata
I fagi capaci di dare trasduzione hanno un DNA costituito dall’assemblaggio di frammenti virali molto simili a frammenti di DNA batterico lisato.

17
Durante l’assemblaggio è possibile lo scambio di frammenti di DNA virale con quelli del DNA batterico Il fago trasducente (1/103 della popolazione virale) può infettare un altro batterio ma non iniziare un normale ciclo di infezione in quanto il DNA virale è assente o difettoso (particella difettiva). Risultato alternativo: trasmissione di nuovi caratteri
 può infettare un altro batterio ma non iniziare un normale ciclo di infezione in quanto il DNA virale è assente o difettoso (particella difettiva). Risultato alternativo: trasmissione di nuovi caratteri.")
18
Fago trasducente DNA batterico o ibrido Infetta un’altro batterio
Il virus, date le piccoledimensioni, può contenere non più del 3% del cromosoma batterico Nuovi alleli, o anche nuovi geni, possono essere integrati nel cromosoma ospite. Dal 70 al 90% del DNA trasdotto non riesce a integrarsi nel cromosoma ospite. I frammenti non integrati rimangono nel citoplasma ed si esprimono solo per un numero limitato di generazioni (trasduzione abortiva)
")
19
Coniugazione Pili sessuali: presenti in numero di 1-10 per cellula, sono spessi 9-10 nm

20
Plasmidi coniugativi F- F+ F+ 94 Kb
IS3 gd IS2 inc, rep oriT A L E B C F H G S D I J K 94 Kb Sono indicati come «fattori F» (o plasmidi F). Oltre a informazioni varie accessorie, contengono le informazioni per costruire i pili sessuali (essenziali per «agganciare» i batteri sprovvisti di fattore F e «veicolare» il passaggio di materiale genetico) F+ F- F+
. Oltre a informazioni varie accessorie, contengono le informazioni per costruire i pili sessuali (essenziali per «agganciare» i batteri sprovvisti di fattore F e «veicolare» il passaggio di materiale genetico) F+ F- F+")
21
Il fattore F, in alcuni casi, riesce a integrarsi nel cromosoma batterico.
In questo caso si formano le cellule Hfr (High frequency recombination)
")
22
Enzimi di restrizione Sempre intorno agli anni ‘50 del secolo scorso, alcune esperienze condotte relativamente ai processi di ricombinazione genetica batterica nell’E. Coli evidenziarono un comportamento significativo in alcuni suoi ceppi. Il materiale genetico estraneo, una volta introdotto nel batterio ospite, veniva frammentato in pezzi più piccoli impedendone la sua attività (tale fenomeno venne definito come restrizione genica). Tali ceppi batterici manifestavano resistenza all’infezioni di molti fagi (il DNA virale veniva «ristretto», frammentato). Solo qualche decennio dopo vennero identificati, nell’E. Coli, una serie di enzimi capaci di degradare (restringere) molecole di DNA. Furono, quindi, chiamati enzimi di restrizione.
. Tali ceppi batterici manifestavano resistenza all’infezioni di molti fagi (il DNA virale veniva «ristretto», frammentato). Solo qualche decennio dopo vennero identificati, nell’E. Coli, una serie di enzimi capaci di degradare (restringere) molecole di DNA. Furono, quindi, chiamati enzimi di restrizione.")
23
Un enzima di restrizione «riconosce» precise sequenze di nucleotidi del DNA (siti di restrizione). Vi si aggancia ed opera dei precisi tagli (rottura dei legami fosfodiesterici). Essendo costituito da una cortissima sequenza di nucleotidi, un sito di restrizione si ritrova ripetuto varie volte e a varie distanze sul DNA. Questo spiega la «frammentazione» di tale macromolecola.

24
Gli enzimi di restrizione (Endonucleasi di restrizione)
Sono enzimi capaci di riconoscere specifiche sequenze di nucleotidi (siti di restrizione) e di tagliare la molecola in quei punti. I tagli possono lasciare estremità piatte o sfalsate In quest’ultimo caso, data la complementarietà delle basi, i frammenti risultanti presentano estremità (o code) coesive (stick ends). Sono stati, al momento, identificati alcune migliaia di enzimi di restrizione, ma i siti sono poco più di 200.
 e di tagliare la molecola in quei punti. I tagli possono lasciare estremità piatte o sfalsate. In quest’ultimo caso, data la complementarietà delle basi, i frammenti risultanti presentano estremità (o code) coesive (stick ends). Sono stati, al momento, identificati alcune migliaia di enzimi di restrizione, ma i siti sono poco più di 200.")
25
I vari frammenti possono «riaccoppiarsi» spontaneamente, grazie proprio a queste code coesive complementari. Tuttavia solo l’azione di una DNA-ligasi potrà ricostruire un’unica catena dai due frammenti. L’accoppiamento può avvenire anche tra due frammenti con estremità «piatte», ma è meno frequente.

26
L’Eco R-I riconosce e taglia la seguente sequenza
Enzima Sito di riconoscimento EcoR I G A A T T C C T T A A G BamH I G G A T C C C C T A G G Hind III A A G C T T T T C G A A KPN I G G T A C C C C A T G G NOT I G C G G C C G C C G C C G G C G EcoR V G A T A T C C T A T A G Xho I C T C G A G G A G C T C I primi enzimi di restrizione, isolati dall’E. Coli, furono di tre tipi indicati con Eco R-I Eco R-II Eco R-III Escherichia coli ceppo R. Il numero romano indica l’ordine di scoperta.

27
Una sequenza specifica (sito di restrizione) non è unica, ma si riscontra in vari punti e a varie distanze lungo tutta la molecola del DNA. In tal modo un enzima specifico riesce a creare frammenti di varie dimensioni (e peso) tutte aventi le stesse «code». L’elettroforesi su gel è una tecnica utilizzata per separare i frammenti di DNA in funzione delle loro diverse dimensioni. Sfrutta la carica negativa dell’acido nucleico (e dei suoi frammenti).
 tutte aventi le stesse «code». L’elettroforesi su gel è una tecnica utilizzata per. separare i frammenti di DNA in funzione delle loro diverse dimensioni. Sfrutta la carica negativa dell’acido nucleico (e dei suoi frammenti).")
28
Utilizzando vari enzimi di restrizione su molecole di DNA derivanti da un unico ceppo di batteri (o tipo di cellule eucariote) è possibile costruire la mappa del DNA.
 è possibile costruire la mappa del DNA.")
29
La conseguenza più significativa della scoperta degli enzimi di restrizione fu la possibilità del loro uso per la «manipolazione genetica». Occorre precisare che gli studi sulla struttura degli acidi nucleici e delle proteine avevano già evidenziato come la loro composizione chimica fondamentale è la stessa in tutti gli organismi viventi (4 nucleoidi per gli acidi nucleici organizzati in triplette e 20 aminoacidi per le proteine riscontrabili in tutti gli esseri viventi) e che la relativa differenza è da imputare solo nel numero e nella sequenza specifica delle unità molecolari. Anche diversi processi metabolici (respirazione, sintesi proteica, fermentazione ecc.) se non uguali, fu dimostrato essere simili e in buona parte sovrapponibili nei vari organismi. Questo fa sì che i frammenti di DNA ottenuti dopo la «restrizione» (taglio), non siano più riconducibili agli organismi originali. Se mischiati, i frammenti aventi code «complementari» possono accoppiarsi in modo casuale e indifferente, indipendentemente dalla loro origine.
 e che la relativa differenza è da imputare solo nel numero e nella sequenza specifica delle unità molecolari. Anche diversi processi metabolici (respirazione, sintesi proteica, fermentazione ecc.) se non uguali, fu dimostrato essere simili e in buona parte sovrapponibili nei vari organismi. Questo fa sì che i frammenti di DNA ottenuti dopo la «restrizione» (taglio), non siano più riconducibili agli organismi originali. Se mischiati, i frammenti aventi code «complementari» possono accoppiarsi in modo casuale e indifferente, indipendentemente dalla loro origine.")
30
DNA ricombinante Le prime semplici esperienze di ingegneria genetica, basate propri sugli enzimi di restrizione, furono condotte utilizzando i plasmidi. Sottoponendo ad uno stesso enzima di restrizione i plasmidi di due ceppi batterici aventi caratteri accessori differenti (ad esempio uno resistente alla penicillina, l’altro no), miscelando i vari frammenti ottenuti e trattando il tutto con DNA – ligasi, si ottennero nuovi plasmidi che introdotti (per trasformazione) in altri batteri furono in grado di trasmettere a quest’ultimi caratteri nuovi. Questo procedimento fu nominato tecnica del DNA ricombinante e i batteri ottenuti furono i primi OGM. Con tale tecnica è possibile costruire plasmidi completamente «personalizzati», contenenti, cioè, una serie di informazioni scelte dall’ingegnere genetico. Di conseguenza è possibile produrre batteri con i caratteri desiderati per poi produrne in quantità industriale (data la facilità con cui questi microrganismi si moltiplicano clonazione dei caratteri) . Il plasmidio, perciò, è definito vettore di clonaggio della tecnica del DNA ricombinante. Questa tecnica è utilizzata per «produrre» farmaci a basso costo (clonaggio molecolare antibiotici, ormoni, vaccini ecc.) Es. l’insulina plasmide chimerico in E. Coli (in commercio dal 1982).
, miscelando i vari frammenti ottenuti e trattando il tutto con DNA – ligasi, si ottennero nuovi plasmidi che introdotti (per trasformazione) in altri batteri furono in grado di trasmettere a quest’ultimi caratteri nuovi. Questo procedimento fu nominato tecnica del DNA ricombinante e i batteri ottenuti furono i primi OGM. Con tale tecnica è possibile costruire plasmidi completamente «personalizzati», contenenti, cioè, una serie di informazioni scelte dall’ingegnere genetico. Di conseguenza è possibile produrre batteri con i caratteri desiderati per poi produrne in quantità industriale (data la facilità con cui questi microrganismi si moltiplicano clonazione dei caratteri) . Il plasmidio, perciò, è definito vettore di clonaggio della tecnica del DNA ricombinante. Questa tecnica è utilizzata per «produrre» farmaci a basso costo (clonaggio molecolare antibiotici, ormoni, vaccini ecc.) Es. l’insulina plasmide chimerico in E. Coli (in commercio dal 1982).")
31
Rappresentazione schematica del clonaggio
DNA SORGENTE DNA BERSAGLIO VETTORE DI CLONAZIONE 1 Restrizione Restrizione 2 frammentazione linearizzazione congiungimento DNA bersaglio- vettore di clonazione 3 Ligasi Plasmide ibrido o chimerico (DNA ricombinato)
")
32
4 5 cellula ospite Riproduzione batterica (clonazione -amplificazione)
Introduzione del DNA nella cellula ospite (trasformazione) e isolamento delle cellule con il gene clonato 4 cellula ospite Riproduzione batterica (clonazione -amplificazione) 5 Produzione della proteina codificata dal gene clonato (Clonazione molecolare)
 e isolamento delle cellule con il gene clonato. 4. cellula ospite. Riproduzione batterica (clonazione -amplificazione) 5. Produzione della proteina codificata dal gene clonato. (Clonazione molecolare)")
33
L’accoppiamento dei frammenti è casuale per cui si verificano varie situazioni:
Alcuni frammenti non riescono ad «assemblarsi» in modo efficace non si forma alcun plasmide. Si riforma il plasmide originario Si forma il plasmide ibrido desiderato. Aggiungendo al gene da clonare il gene per la resistenza ad un antiobiotico (es. all’ampicillina), i batteri che inglobano il plasmide desiderato acquisteranno tale resistenza per cui, alla fine dell’esperienza, saranno gli unici a sopravvivere al trattamento con l’antibiotico specifico. Questo permette di «purificare» il bioreattore prodotto.
, i batteri che inglobano il plasmide desiderato acquisteranno tale resistenza per cui, alla fine dell’esperienza, saranno gli unici a sopravvivere al trattamento con l’antibiotico specifico. Questo permette di «purificare» il bioreattore prodotto.")
34
Utilizzi farmaceutici
Insulina. Costituita da due catene proteiche (alfa e beta), i rispettivi geni vengono prima legati al gene per Lac-Z (rende più stabile il plasmide chimerico) e poi impiantati in plasmidi differenti trasmessi, a loro volta, in batteri di E. coli separati. I prodotti dei due bio-reattori verranno poi assemblate chimicamente. Vaccino contro virus epatite B Prodotto negli anni 1986, questo vaccino è costituito dall’antigene di superficie del virus (HBsAg) prodotto da un lievito (Saccharomyces cerevisiae). Precedentemente venivano usati HBsAg estratti da plasma ematico di persone siero-positive al virus dell’epatite B. Tale vaccino produceva vari effetti collaterali dovute alla presenza di altre molecole chimiche plasmatiche. Il vaccino prodotto con la tecnica del DNA ricombinante è praticamente «puro» Vaccini, ormone della crescita (GH o STH), interferone, fattori di coagulazione, anticorpi, citochine, LH, FSH, enzimi antiinfiammatori, antitumorali, ecc. ecc.
, i rispettivi geni vengono prima legati al gene per Lac-Z (rende più stabile il plasmide chimerico) e poi impiantati in plasmidi differenti trasmessi, a loro volta, in batteri di E. coli separati. I prodotti dei due bio-reattori verranno poi assemblate chimicamente. Vaccino contro virus epatite B. Prodotto negli anni 1986, questo vaccino è costituito dall’antigene di superficie del virus (HBsAg) prodotto da un lievito (Saccharomyces cerevisiae). Precedentemente venivano usati HBsAg estratti da plasma ematico di persone siero-positive al virus dell’epatite B. Tale vaccino produceva vari effetti collaterali dovute alla presenza di altre molecole chimiche plasmatiche. Il vaccino prodotto con la tecnica del DNA ricombinante è praticamente «puro» Vaccini, ormone della crescita (GH o STH), interferone, fattori di coagulazione, anticorpi, citochine, LH, FSH, enzimi antiinfiammatori, antitumorali, ecc. ecc.")
35
Libreria genica e genomica
Collezione di una serie di ceppi batterici, ognuno dei quali contenenti un plasmide con un frammento specifico del DNA di un cromosoma (genica) o di tutti i cromosomi di un organismo (genomica). La collezione completa contiene tutti i geni di un cromosoma (genica) o di tutti i cromosomi (genomica). Estrazione del DNA cromosomico 1 Taglio del DNA con endonucleasi di restrizione (digestione parziale o totale) 2 Inserimento di ogni frammento di DNA in un vettore 3 4 Trasformazione batterica
 o di tutti i cromosomi di un organismo (genomica). La collezione completa contiene tutti i geni di un cromosoma (genica) o di tutti i cromosomi (genomica). Estrazione del DNA. cromosomico. 1. Taglio del DNA con. endonucleasi di restrizione. (digestione parziale o totale) 2. Inserimento di ogni frammento. di DNA in un vettore Trasformazione batterica.")
36
Il cDNA I primi farmaci prodotti, tuttavia, risultarono poco o per nulla efficaci. Le molecole prodotto non erano perfettamente uguali a quelle naturali. La spiegazione fu trovata scoprendo che l’informazione sul DNA (gene) è grezza, fatta da introni ed esoni. Solo con il processo dello splicing a carico dell’RNA messaggero si ottiene la sintesi di una molecola sgrezzata e perfettamente funzionante. Ora, tale processo richiede complessi enzimatici specifici, non comuni ai vari tipi organismi, per cui la stessa informazione del DNA viene processata in modo differente nei diversi tipi di cellule. Per ovviare a ciò si utilizza la tecnica del cDNA (trascrittasi inversa) In questa tecnica, si isola l’RNA messaggero presente nel citoplasma (che, quindi, ha già subito lo splicing) contente l’informazione sgrezzata della proteina desiderata. Si tratta tale RNA messaggero con l’enzima virale trascrittasi inversa per ottenere il cDNA. Sarà quest’ultimo a essere utilizzato nella tecnica del DNA ricombinante.
 è grezza, fatta da introni ed esoni. Solo con il processo dello splicing a carico dell’RNA messaggero si ottiene la sintesi di una molecola sgrezzata e perfettamente funzionante. Ora, tale processo richiede complessi enzimatici specifici, non comuni ai vari tipi organismi, per cui la stessa informazione del DNA viene processata in modo differente nei diversi tipi di cellule. Per ovviare a ciò si utilizza la tecnica del cDNA (trascrittasi inversa) In questa tecnica, si isola l’RNA messaggero presente nel citoplasma (che, quindi, ha già subito lo splicing) contente l’informazione sgrezzata della proteina desiderata. Si tratta tale RNA messaggero con l’enzima virale trascrittasi inversa per ottenere il cDNA. Sarà quest’ultimo a essere utilizzato nella tecnica del DNA ricombinante.")
37
Polymerase Chain Reaction - PCR
La PCR rappresenta la seconda rivoluzione nelle tecniche di manipolazione del DNA E’ essenzialmente una tecnica di amplificazione del DNA E’ usata per ottenere grandi quantità di DNA da campioni limitati (anche da materiale vecchio e degradato mummie, fossili ecc.) E’ una tecnica che «copia» il processo della duplicazione del DNA. Essa consiste, sinteticamente nel: Digerire gli istoni e le varie proteine che tengono il DNA raggomitolato. Scaldare la molecola a 95° per ottenere la separazione delle catene complementari Aggiungere specifici primer (si tratta di oligonucleotidi, cioè di brevi sequenze di nucleotidi) complementari ai primi tratti del gene da amplificare (ognuno ha il suo primer) Raffreddare a circa 50° per permettere l’accoppiamento spontaneo dei primer Aggiungere i nucleotidi liberi Aggiungere DNA polimerasi per la formazione di nuove catene. Il ciclo si può ripetere all’infinito con le molecole man mano prodotte
 E’ una tecnica che «copia» il processo della duplicazione del DNA. Essa consiste, sinteticamente nel: Digerire gli istoni e le varie proteine che tengono il DNA raggomitolato. Scaldare la molecola a 95° per ottenere la separazione delle catene complementari. Aggiungere specifici primer (si tratta di oligonucleotidi, cioè di brevi sequenze di nucleotidi) complementari ai primi tratti del gene da amplificare (ognuno ha il suo primer) Raffreddare a circa 50° per permettere l’accoppiamento spontaneo dei primer. Aggiungere i nucleotidi liberi. Aggiungere DNA polimerasi per la formazione di nuove catene. Il ciclo si può ripetere all’infinito con le molecole man mano prodotte.")
38
Identificazione di un gene
Attualmente, per produrre una proteina specifica in quantità industriale: Si studia la struttura primaria della proteina per conoscere la sua sequenza di amminoacidi Attraverso il codice genetico si risale alla sequenza di nucleotidi corrispondenti nell’RNA messaggero. Si isola tale mRNA dal citoplasma della cellula Tramite trascrittasi inversa di sintetizza una catena di cDNA utilizzando nucleotidi marcati radioattivamente. Sui miscela tale catena di cDNA marcato con l’intero genoma della cellula. Il cDNA si «legherà», per complementarietà, in un tratto specifico di un cromosoma. In tal modo si può identificare ed isolare il gene per la proteina. Una volta isolato lo si può amplificare e impiantare in un altro organismo.

39
Vettore virale Grazie alla scoperta degli enzimi di restrizione, e alla tecnologia del DNA ricombinante, è stato possibile dotare alcuni batteri di sistemi in grado di «restringere» il DNA (o RNA) virale in modo tale da fargli perdere la sua tossicità, ma lasciandogli intatta la capacità di infettare e di integrare il proprio materiale genetico con quello dell’ospite (ciclo lisogeno) La tecnologia riuscì anche a ricombinare il DNA virale in modo da «agganciargli» frammenti estranei. In tal modo il virus ricombinato diventa vettore genetico. Infetta l’ospite senza comportrne la morte, bensì una sua ricombinazione. I vantaggi di un vettore virale sono: l’efficienza della ricombinazione (si sfrutta la capacità naturale di integrazione del DNA ospite) Gli svantaggi: - Possibilità di produrre virus patogeni (anche tumori) - Reazione immunitaria - Frammenti di DNA piccoli
 virale in modo tale da fargli perdere la sua tossicità, ma lasciandogli intatta la capacità di infettare e di integrare il proprio materiale genetico con quello dell’ospite (ciclo lisogeno) La tecnologia riuscì anche a ricombinare il DNA virale in modo da «agganciargli» frammenti estranei. In tal modo il virus ricombinato diventa vettore genetico. Infetta l’ospite senza comportrne la morte, bensì una sua ricombinazione. I vantaggi di un vettore virale sono: l’efficienza della ricombinazione (si sfrutta la capacità naturale di integrazione del DNA ospite) Gli svantaggi: - Possibilità di produrre virus patogeni (anche tumori) - Reazione immunitaria. - Frammenti di DNA piccoli.")
40
Vettore Cosmidico In seguito allo studio del batteriofago lambda (il cui DNA non si integra nel cromosoma dell’ospite, ma si ciclizza nel suo citoplasma divenendo plasmide, con possibilità di autoreplicarsi ed esprimersi) furono identificate e isolate due sequenze nuclotidiche estreme a catena singola complementari (dette cosR e cosL) responsabili, appunto, della ciclizzazione. Grazie a tali frammenti è possibile costruire un plasmide artificiale costituito dalle cos in aggiunta ad altri geni desiderati: si ottiene il cosmide. Introdotto trasforma l’ospite in modo efficiente senza grossi svantaggi.
 furono identificate e isolate due sequenze nuclotidiche estreme a catena singola complementari (dette cosR e cosL) responsabili, appunto, della ciclizzazione. Grazie a tali frammenti è possibile costruire un plasmide artificiale costituito dalle cos in aggiunta ad altri geni desiderati: si ottiene il cosmide. Introdotto trasforma l’ospite in modo efficiente senza grossi svantaggi.")
41
Agrobacter tumefaciens
Verso la fine degli anni ‘80 del secolo scorso si scoprì che l’agrobacter tumefaciens, responsabile del tumore al colletto di molte piante, agisce attuando un meccanismo di coniugazione con le cellule vegetali. In queste veniva trasferito un plasmide, Il Ti (tumor inducens). Il Ti è fatto da un DNA circolare contenente diversi frammenti genici, tra cui: Il T-DNA (8 geni responsabili del tumore) Due frammenti Vir (responsabili dell’integrazione del T-DNA nel DNA Frammenti F per la coniugazione Altri geni accessori
. Il Ti è fatto da un DNA circolare contenente diversi frammenti genici, tra cui: Il T-DNA (8 geni responsabili del tumore) Due frammenti Vir (responsabili dell’integrazione del T-DNA nel DNA. Frammenti F per la coniugazione. Altri geni accessori.")
42
Osservare il DNA A metà circa degli anni ’70 Frederick Sanger introdusse la prima tecnica per il sequenziamento del DNA, ovvero per la determinazione della successione di basi in una sequenza di acidi nucleici (In parole povere la «lettura» del codice genetico) Tale tecnica è basata sul naturale meccanismo di duplicazione del DNA (DNA polimerasi) e sull’utilizzo di di-desossiribonucleotidi (ddNtP). dNtP ddNtP
 Tale tecnica è basata sul naturale meccanismo di duplicazione del DNA (DNA polimerasi) e sull’utilizzo di di-desossiribonucleotidi (ddNtP). dNtP. ddNtP.")
43
Tecnica di Sanger 1 Si parte da molecole di DNA replicate tramite tecnica PCR o tramite clonaggio in plasmidi di E. Coli 2 Ottenuta una certa quantità di molecole di DNA, si purificano e si trattano al calore (95°C) in modo da ottenere catene singole.
 in modo da ottenere catene singole.")
44
Il materiale ottenuto si distribuisce equamente in 4 provette
3 Il materiale ottenuto si distribuisce equamente in 4 provette 4 In ognuna si aggiunge una pari quantità dei 4 nucleotidi (dNtP) liberi
 liberi.")
45
DNA-polimerasi + primer
5 In ciascuna provetta si aggiunge uno specifico tipo di di-desossiribonucleotide (ddNtP), cioè un desossiribonuclotide mancante di un ossigeno in posizione 3’ (oltre che in posizione 2’) ddNtP ddATP ddGTP ddTTP ddCTP DNA-polimerasi + primer 5 In ciascuna provetta si aggiunge il complesso enzimatico DNA-polimerasi corredato dal suo primer (sequenza di nucleotidi che agganciandosi in modo complementare all’inizio della catena di DNA permette l’attacco dell’enzima)
, cioè un desossiribonuclotide mancante di un ossigeno in posizione 3’ (oltre che in posizione 2’) ddNtP. ddATP. ddGTP. ddTTP. ddCTP. DNA-polimerasi + primer. 5. In ciascuna provetta si aggiunge il complesso enzimatico DNA-polimerasi corredato dal suo primer (sequenza di nucleotidi che agganciandosi in modo complementare all’inizio della catena di DNA permette l’attacco dell’enzima)")
46
5 La DNA polimerasi inizierà la «costruzione» di nuove catene di DNA, a stampo su quelle originali presenti, utilizzando i nucleotidi liberi a disposizione.

47
La RNA polimerasi non distingue i dNTP dai ddNTP, per cui li utilizza indifferentemente.
Ora, quando «incorpora» nella sequenza un ddNTP, la polimerizzazione si interrompe in quanto non sarà possibile formare il legame tra il fosfato e lo zucchero in posizione 3’ (manca il gruppo -OH per la «condensazione». Dato che, in una provetta, l’ultilizzo di uno specifico ddNTP (ad es, ddCTP) al posto del corrispondente dNTP (dCTP) è casuale, l’interruzione si avrà a varie distanza dall’inizio delle varie molecole di DNA, il che darà, alla fine, frammenti di varia lunghezza tutti terminanti, però, con lo stesso tipo di ddNTP (nel nostro esempio: ddCTP)
 al posto del corrispondente dNTP (dCTP) è casuale, l’interruzione si avrà a varie distanza dall’inizio delle varie molecole di DNA, il che darà, alla fine, frammenti di varia lunghezza tutti terminanti, però, con lo stesso tipo di ddNTP (nel nostro esempio: ddCTP)")
48
Dato che in ogni provetta si è inserito un diverso tipo di ddNTP, i frammenti di ognuna saranno caratterizzati da nucleotidi terminali differenti. Si allestisce un’analisi elettroforetica presentante 4 pozzetti in ognuno dei quali si appoggia una goccia di ciascuna provetta. Alla fine dell’esame (trascinamento elettroforetico), si avrà un grafico con bande colorate in diverse posizioni (a seconda della grandezza dei frammenti in gioco) da cui si ricava la sequenza finale.
, si avrà un grafico con bande colorate in diverse posizioni (a seconda della grandezza dei frammenti in gioco) da cui si ricava la sequenza finale.")
50
Piante transgeniche cibi OGM
Ricavare il Ti (plasmide dell’agrobacter tumefaciens) Trattarlo con enzimi di restrizione appositi per «tagliare» il frammento T-DNA. Eliminare il frammento T-DNA Miscelare il resto del Ti con geni contenenti informazioni volute Trattare con DNA-ligasi Trasferire il plasmide ibrido in batteri comuni Infettare embrioni di piante. I batteri trattati saranno in grado di trasferire nelle cellule vegetali infettate nuovi caratteri, senza determinare tumore. Piante transgeniche cibi OGM
 Trattarlo con enzimi di restrizione appositi per «tagliare» il frammento T-DNA. Eliminare il frammento T-DNA. Miscelare il resto del Ti con geni contenenti informazioni volute. Trattare con DNA-ligasi. Trasferire il plasmide ibrido in batteri comuni. Infettare embrioni di piante. I batteri trattati saranno in grado di trasferire nelle cellule vegetali infettate nuovi caratteri, senza determinare tumore. Piante transgeniche cibi OGM.")
51
Terapia genica Insieme di procedimenti atti a curare o ad alleviare una malattia modificando geneticamente le cellule dei pazienti.

52
La terapia genica è particolarmente
adatta a certi tipi di patologia: le malattie ereditarie monogeniche (dovute a deficienze genetiche di un singolo prodotto genico o dall'errata espressione di un gene) le malattie infettive (causate dall'infezione di un singolo agente patogeno sia virale sia batterico) le malattie del sistema immunitario (che comprendono soprattutto le allergie, le infiammazioni e le malattie autoimmuni) i tumori (causati dall'errata prosecuzione della divisione e della proliferazione cellulare a seguito dell'attivazione di un oncogene o dell'inattivazione di un oncosoppressore)
 le malattie infettive (causate dall infezione di un singolo agente patogeno sia virale sia batterico) le malattie del sistema immunitario (che comprendono soprattutto le allergie, le infiammazioni e le malattie autoimmuni) i tumori (causati dall errata prosecuzione della divisione e della proliferazione cellulare a seguito dell attivazione di un oncogene o dell inattivazione di un oncosoppressore)")
53
Vantaggi teorici della terapia genica
Possibilità di agire su meccanismi molecolari per i quali risulta estremamente difficile sviluppare farmaci specifici Correzione radicale dei difetti Vantaggi economici (se fosse permanente eviterebbe la necessità di trattamenti ripetuti)
")
54
L'idea della terapia genica
è nata verso la fine degli anni '70 quando per la prima volta è stato clonato un gene. Il primo gene clonato era quello codificante per la b-globina, una proteina che da sola costituisce circa il 97% del globulo rosso maturo e che se viene espressa in modo errato o in quantità ridotta causa la talassemia. La possibilità di intervenire su questo gene in modo da ottenere globuli rossi sani e quindi non più talassemici ha dato il via ai primi numerosi tentativi di terapia genica. Per curare la talassemia occorre trasfettare il gene della b-globina nelle cellule progenitrici del globulo rosso che sono però numericamente pochi e difficili da isolare. Inoltre l'emogobina è una proteina espressa in modo strutturale dal globulo rosso e per correggere la talassemia occorre raggiungere altissimi livelli di espressione del gene inserito.

55
La terapia genica si può basare su:
inserimento di geni umani clonati inserimento di segmenti genici che possono derivare da altri genomi inserimento di segmenti genici sintetizzati in laboratorio.

56
Il materiale genetico può venire trasferito:
nelle cellule precedentemente prelevate dal paziente e solo in seguito reintrodotte nel paziente stesso terapia genica ex-vivo direttamente nelle cellule nel corpo di un paziente terapia genica in-vivo

57
Strategie terapeutiche
In caso di mutazioni che hanno reso difettoso o non funzionante un gene. Individuare la proteina difettosa o non funzionante Risalire al gene specifico Isolare tale gene da cellule sane (o sintetizzzarlo) Impiantarlo nelle cellule malate
 Impiantarlo nelle cellule malate.")
58
In caso di mutazioni ad effetto dominante negativo.
Strategie terapeutiche In caso di mutazioni ad effetto dominante negativo. 1) Si studia la sequenza del gene mutante pericoloso 2) Si individua (o si sintetizza) un gene anti-senso (è una sequenza di nucleotidi complementare al gene in questione) 3) Si introduce il gene anti-senso nelle cellule malate 4) Il gene anti-senso si legherà al gene mutante bloccandolo.
 Si studia la sequenza del gene mutante pericoloso. 2) Si individua (o si sintetizza) un gene anti-senso (è una sequenza di nucleotidi complementare al gene in questione) 3) Si introduce il gene anti-senso nelle cellule malate. 4) Il gene anti-senso si legherà al gene mutante bloccandolo.")
59
Strategie terapeutiche
Uccisione diretta di cellule patologiche (in genere tumorali) Gene che codifica un oncosoppressore Virus oncolitico
 Gene che codifica un oncosoppressore. Virus oncolitico.")
60
Gene che codifica un oncosoppressore
Strategie terapeutiche Inserimento di geni Gene che codifica un oncosoppressore Virus oncolitico

61
Vettori non virali Vettori virali Integrazione Casuale o
Come faccio entrare il DNA nelle cellule, e cosa gli succede quando è entrato? Vettori non virali Vettori virali Integrazione Casuale o sito-specifica

62
Lentivirus (Es. HIV) Adenovirus Herpes simplex
 Adenovirus Herpes simplex")
63
Herpes simplex virus Gli herpes simplex virus, grazie alla loro naturale capacità di stabilire infezioni latenti nei neuroni, vengono utilizzati per il trasporto di geni nel CNS. Il genoma degli herpes virus è costituito da un doppio filamento di DNA di 152kb, contiene più di 80 geni, la metà dei quali non risulta essenziale per la crescita in cellule di coltura; una volta eliminati tali geni è possibile inserire un transgene di grosse dimensioni (circa 40-50kb).
.")
64
DNA nudo Consiste nell’iniettare il gene terapeutico, legato ad un plasmide o a cristalli di fosfato di calcio direttamente nella cellula tramite l’utilizzo d’una micropipetta. Lo svantaggio di questa metodica consiste nel fatto che bisogna iniettare il DNA in ogni cellula, una per una. Il rendimento, inoltre, è decisamente basso. L’inserzione di DNA nudo è la procedura più lineare e più semplice ed inoltre permette di trasferire costrutti genici di grandi dimensioni.

65
DNA in Liposomi Usando liposomi cationici è possibile far complessare ad essi il DNA, che a pH neutro presenta carica negativa. Il complesso DNA-liposoma può fondersi con la membrana cellulare ma nella maggior parte dei casi viene internalizzato tramite endocitosi. Successivamente il DNA viene liberato nel citoplasma, entra nel nucleo e viene espresso. Sfortunatamente questo processo è a bassa efficienza in quanto si è visto che solo lo 0,1% del DNA introdotto viene espresso. Per ovviare a ciò nei liposomi sono state anche inserite proteine ed anticorpi che possano aumentare l’efficacia della procedura minimizzando la degradazione del DNA e facilitando il corretto direzionamento della vescicola.

66
Polimeri cationici Molto simile alla tecnica dei liposomi è la procedura di trasfezione che utilizza polimeri cationici, infatti macromolecole dotate di molteplici cariche positive possono interagire con il DNA, il quale a pH fisiologico è un polianione, provocandone la condensazione e proteggendolo da aggressivi sia chimici che enzimatici, oltre che da radiazioni ionizzanti. Anche i complessi DNA-policatione vengono internalizzati dalla cellula per endocitosi, e possono essere attivamente indirizzati verso specifiche linee cellulari o tessuti utilizzando anticorpi o altre molecole direzionanti.

67
Bombardamento tramite particelle (gene gun)
Il bombardamento tramite particelle consiste nell’utilizzo di particolari strumenti elettrici o ad alta pressione, dette pistole geniche (gene gun), che permettono di inviare nella cellula particelle microscopiche d’oro o di tungsteno ricoperte di DNA. Al momento non esistono studi sull’uomo di questa metodica ma solo su animali.
, che permettono di inviare nella cellula particelle microscopiche d’oro o di tungsteno ricoperte di DNA. Al momento non esistono studi sull’uomo di questa metodica ma solo su animali.")
68
Principali malattie oggetto di studio per la terapia genica:
Deficit Incidenza Cellule bersaglio GENETICHE SCID Adenosina deaminasi IL-2R gamma Rara Midollo osseo Linfociti T Emofilia A e B Fattori VIII o IX 1:10,000 maschi Fegato, musc. Ipercolesterolemia familiare Recettore per LDL 1:1,000,000 Fegato Fibrosi cistica Gene CFTR 1:3,000 caucasici Vie resp. Emoglobinopatie (tal., SCA) Difetti in - o -globina Fino a 1:600 Enfisema ereditario 1-antitripsina 1:3,500 Polmoni,fegato ACQUISITE Cancro Vari > 1 milione/anno Vari tipi Malattie neurologiche Parkinson, Alzheimer,SLA 1 milione/anno Neuroni, glia Malattie infettive AIDS, epatite C CellT, MF,feg. Malattie dell’occhio Retinopatia diabetica Degenerazione maculare Epitelio retinico
 Difetti in - o -globina. Fino a 1:600. Enfisema ereditario. 1-antitripsina. 1:3,500. Polmoni,fegato. ACQUISITE. Cancro. Vari. > 1 milione/anno. Vari tipi. Malattie neurologiche. Parkinson, Alzheimer,SLA. 1 milione/anno. Neuroni, glia. Malattie infettive. AIDS, epatite C. CellT, MF,feg. Malattie dell’occhio. Retinopatia diabetica. Degenerazione maculare. Epitelio retinico.")
69
Fibrosi cistica: malattia autosomica recessiva
terapia genica di patologie ereditarie Fibrosi cistica: malattia autosomica recessiva determinata da un deficit di trasporto degli ioni cloro attraverso le cellule epiteliali dovuta a mutazioni nel gene CFTR Quadro patologico: distruzione del parenchima polmonare insufficienza pancreatica Terapia genica per trasferire un minigene CFTR utilizza : Adenovirus ricombinante: stabilire prima la massima dose tollerabile Liposomi: più sicuri, ma con efficienza inferiore.

70
terapia genica contro la cecità Moorfields Eye Hospital
«degenerazione ereditaria della retina» patologia progressiva causata da un’alterazione di un singolo gene RPE65 L’intervento: Inserire una versione sana del gene nei cromatociti Il gene è stato «trasportato» all’interno dell’occhio da un vettore virale, geneticamente modificato, che lo ha inserito nel DNA delle cellule. per mandare il gene a più cellule possibili, deve essere usata una quantità tale di soluzione da consentire di alzare la retina causandone il temporaneo distacco University College di Londra Moorfields Eye Hospital (The Indipendent, 2007)
")
71
Limitazioni della terapia genica
Rischio troppo elevato in presenza di terapie alternative Mancanza di efficacia dovuta a problemi tecnici (inefficiente trasferimento genico, inattivazione del gene terapeutico) Costi elevati Alto rischio di effetti indesiderati (espressione non regolata, mutagenesi inserzionale, passaggio delle modifiche alla linea germinale)
 Costi elevati. Alto rischio di effetti indesiderati (espressione non regolata, mutagenesi inserzionale, passaggio delle modifiche alla linea germinale)")
72
Attuali problemi tecnici della terapia genica
Efficienza di trasferimento genico Selettività del sistema di trasferimento Espressione instabile nel tempo Espressione non regolata Reazioni del sistema immunitario Possibili interferenze con il genoma (mutagenesi inserzionale) Problemi etici
 Problemi etici.")
73
Terapia cellulare Allotrapianto: cellule provenienti da un donatore della stessa specie Xenotrapianto: cellule provenienti da animali di specie diverse Autotrapianto: cellule provenienti dallo stesso paziente

74
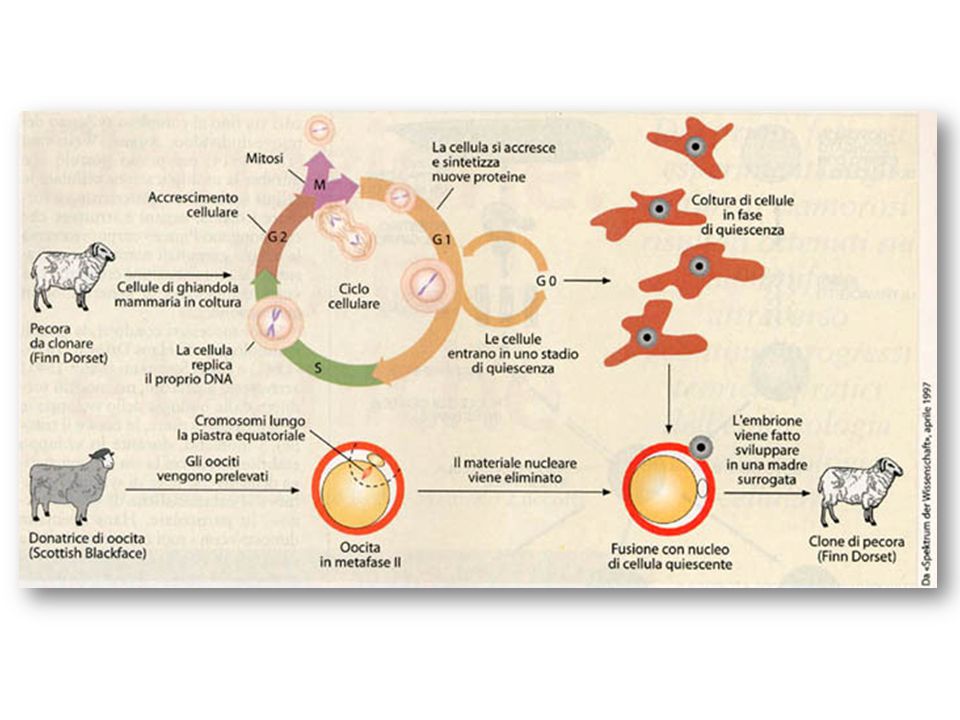
La clonazione animale Dolly

75
Perché qualcuno sta pensando seriamente alla clonazione umana?
(clonazione terapeutica) Il problema principale della terapia cellulare è l’istocompatibilità rigetto La clonazione permetterebbe di ottenere cellule staminali totipotenti dotate delle stesse caratteristiche antigeniche del paziente, da usare per il trapianto direttamente o previa correzione del difetto tramite terapia genica.
 Il problema principale della terapia cellulare è l’istocompatibilità rigetto. La clonazione permetterebbe di ottenere cellule staminali totipotenti dotate delle stesse caratteristiche antigeniche del paziente, da usare per il trapianto direttamente o previa correzione del difetto tramite terapia genica.")
76
Le cellule staminali

77
Pluripotenti (blastociste)
Cellule staminali Totipotenti (zigote) E’ la capacità di una singola cellula di dividersi e di produrre tutte le cellule differenziate di un organismo. Sono le primissime 6-8 (15-20) cellule derivate dalla cellula-uovo fecondata e chiamate blastomeri. Pluripotenti (blastociste) E’ la capacità di una singola cellula di dividersi e di differenziarsi in uno dei tre strati germinali: - Endoderma (rivestimento interno dello stomaco, del tratto gastro-intestinale, i polmoni); - Mesoderma (muscoli, ossa, sangue, urogenitale); Ectoderma (tessuti epidermici e del sistema nervoso); Hanno perso parte della propria potenzialità.
 E’ la capacità di una singola cellula di dividersi e di produrre tutte le cellule differenziate di un organismo. Sono le primissime 6-8 (15-20) cellule derivate dalla cellula-uovo fecondata e chiamate blastomeri. Pluripotenti (blastociste) E’ la capacità di una singola cellula di dividersi e di differenziarsi in uno dei tre strati germinali: - Endoderma (rivestimento interno dello stomaco, del tratto gastro-intestinale, i polmoni); - Mesoderma (muscoli, ossa, sangue, urogenitale); Ectoderma (tessuti epidermici e del sistema nervoso); Hanno perso parte della propria potenzialità.")
78
Utilizzazione: Biologia applicata
le cellule staminali possano essere isolate, caratterizzate e utilizzate comprendendone i meccanismi di proliferazione e differenziazione, per essere coltivate con successo in laboratorio; le cellule staminali cresciute in laboratorio potranno essere condizionate per trasformarsi in uno specifico tipo cellulare; le cellule staminali che hanno formato particolari tipi cellulari potranno essere usate nella cura di pazienti i cui tessuti sono malati o danneggiati; i tessuti e gli organi ottenuti in questo modo possono svilupparsi normalmente senza che ci possano essere rischi per i pazienti

79
Limiti cellule staminali adulte
Non è facile individuarle (l’unica sistema è studiarne il funzionamento) Sono rarissime (nel midollo osseo del topo le staminali sono 1 su ) Crescono molto lentamente e necessitano di particolari cure in coltura.
 Sono rarissime (nel midollo osseo del topo le staminali sono 1 su ) Crescono molto lentamente e necessitano di particolari cure in coltura.")
80
Impieghi terapeutici Ipotesi terapeutiche
Leucemia: trapianto di midollo osseo Ricostruzione dell’epidermide Ipotesi terapeutiche Cura del diabete Trattamento dell’infarto e di alcuni tumori Malattie neurovegetative (in particolare Alzheimer e Parkinson)
")
81
Come ottenerle? dagli embrioni nei primi stadi di sviluppo (blastocisti) creati attraverso fecondazione in vitro (quelli che non sono più necessari per il trattamento dell’infertilità qualche volta detti (embrioni di scorta) o creati specificatamente per la ricerca. dagli embrioni nei primi stadi di sviluppo, creati inserendo il nucleo di una cellula adulta in una cellula uovo a cui era stato rimosso il nucleo (sostituzione del nucleo cellulare più comunemente detta clonazione terapeutica); Da embrioni non impiantabili in utero (embrioni soprannumerari): dalle cellule del sangue del cordone ombelicale al momento della nascita; da alcuni tessuti negli organismi adulti (derma, strati proliferativi delle mucose, parte più interna del cervello …) dalle cellule germinali o dagli organi di un feto abortito (no problemi etici);
 creati attraverso fecondazione in vitro (quelli che non sono più necessari per il trattamento dell’infertilità qualche volta detti (embrioni di scorta) o creati specificatamente per la ricerca. dagli embrioni nei primi stadi di sviluppo, creati inserendo il nucleo di una cellula adulta in una cellula uovo a cui era stato rimosso il nucleo (sostituzione del nucleo cellulare più comunemente detta clonazione terapeutica); Da embrioni non impiantabili in utero (embrioni soprannumerari): dalle cellule del sangue del cordone ombelicale al momento della nascita; da alcuni tessuti negli organismi adulti (derma, strati proliferativi delle mucose, parte più interna del cervello …) dalle cellule germinali o dagli organi di un feto abortito (no problemi etici);")
82
Clonazione Ipotizzata negli anni ’30 del secolo scorso, il primo vero tentativo di questo genere coronato da successo avvenne per opera di John Gordon, il quale nel 1962 riuscì a clonare una rana a partire dal nucleo di una cellula intestinale di un’altra rana adulta. Tuttavia da questo e dai successivi esperimenti era emerso evidente un limite alla clonazione di individui adulti: la perdita della totipotenza dei nuclei di cellule differenziate. La totipotenza è la capacità che cellule non ancora differenziate (generalmente cellule embrionarie) hanno di ricostruire l’intero organismo.
 hanno di ricostruire l’intero organismo.")
83
Dolly Questo limite è stato però superato dagli studi del gruppo di ricercatori del Roslin Institute di Edimburgo che nel 1996 riuscirono ad ottenere pecore vive a partire da una linea cellulare stabilizzata in vitro. Queste cellule, in tutto assimilabili a cellule embrionali totipotenti, furono però indotte in quiescenza mediante deprivazione di siero. Il risultato dell’esperimento era estremamente importante perché la quiescenza è lo stato del ciclo cellulare in cui si trovano normalmente le cellule differenziate che costituiscono l’organismo adulto. L’anno successivo lo stesso gruppo guidato da Ian Whilmut annunciò di essere riuscito a clonare una pecora – nota in tutto il mondo con il nome di Dolly – a partire da un nucleo proveniente da una cellula differenziata.

86
Può avere diversa finalizzazione:
Perché clonare Può avere diversa finalizzazione: Riproduttiva Terapeutica

87
Dopo Dolly 1996 clonazione della pecora Dolly da parte di Wilmut
1998 topi 2000 maiale 2002 Gatto 2002 annuncio della prima clonazione di un embrione umano (attivazione di una cellula uovo) 2003 muore Dolly 2005 Snuppy il primo cane e Little Nick (prima clonazione a pagamento $ dollari) 2007 Wilmut annuncia di aver abbandonato la tecnica della clonazione
 2003 muore Dolly Snuppy il primo cane e Little Nick (prima clonazione a pagamento $ dollari) 2007 Wilmut annuncia di aver abbandonato la tecnica della clonazione.")
88
Costituzione europea Art. II-61 Dignità umana
La dignità umana è inviolabile. Essa deve essere rispettata e tutelata art. II-63 Diritto all’integrità della persona Ogni individuo ha diritto alla propria integrità fisica e psichica. Nell’ambito della medicina e della biologia devono essere in particolare rispettati: a) il consenso libero e informato della persona interessata secondo le modalità definite dalla legge b) il divieto delle pratiche eugenetiche, in particolare di quelle aventi come scopo la selezione delle persone c) il divieto di fare del corpo umano e delle sue parti in quanto tali una fonte di lucro d) il divieto della clonazione riproduttiva degli esseri umani
 il consenso libero e informato della persona interessata secondo le modalità definite dalla legge. b) il divieto delle pratiche eugenetiche, in particolare di quelle aventi come scopo la selezione delle persone. c) il divieto di fare del corpo umano e delle sue parti in quanto tali una fonte di lucro. d) il divieto della clonazione riproduttiva degli esseri umani.")
89
Protocollo addizionale alla Convenzione per la salvaguardia dei diritti dell'uomo e della dignità di essere umano con riguardo alle applicazioni della biologia e della medicina Art. 1 E’ vietato qualsiasi intervento volto a creare un essere umano geneticamente identico a un altro essere umano vivo o morto. Ai fini della presente sezione, l'espressione essere umano "geneticamente identico" ad un altro essere umano significa un essere umano che condivide con un altro l'insieme dei geni nucleari.

Presentazioni simili
















. E’ stato sperimentato in Scozia (1997.>")




