Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il compartimento naturale dell’immunità innata: I recettori Toll-like (TLRs) e le proteine Nods
Dr. Luigi Lembo Fazio Sapienza Università di Roma Dipartimento di Biologia e Biotecnologie “Charles Darwin”

2
Pattern-Recognition Receptors, PRRs
1. Sono molecole recettoriali che riconoscono motivi conservati nell’ambito dei microrganismi (PAMPs) 2. Recettori geneticamente determinati nella linea germinale 3. Sono espressi in diversi tipi cellulari (macrofagi, DC, mast-cells, cellule NK, cellule epiteliali, endoteliali, fibroblasti) 4. Possono essere: a) circolanti (pentraxina PTX3) b) di membrana (TLRs) c) citosolici (proteine Nods/RLRs) Notevoli omologie strutturali e funzionali con i geni R delle piante
 2. Recettori geneticamente determinati nella linea germinale. 3. Sono espressi in diversi tipi cellulari (macrofagi, DC, mast-cells, cellule NK, cellule epiteliali, endoteliali, fibroblasti) 4. Possono essere: a) circolanti (pentraxina PTX3) b) di membrana (TLRs) c) citosolici (proteine Nods/RLRs) Notevoli omologie strutturali e funzionali con i geni R delle piante.")
3
PAMPs: Pathogen-Associated Molecular Patterns
Funghi, batteri e virus producono una serie di molecole (PAMPs) che possono essere riconosciute dall’ospite Prodotti da microrganismi e NON dalle cellule eucariotiche Conservati tra i microrganismi di una data classe Essenziali per la sopravvivenza microbica Prodotti da microrganismi patogeni e NON patogeni (MAMPs: Microbe-Associated Molecular Patterns) Funzione ed espressione trascrizionale diversa dai fattori di virulenza
 che possono essere riconosciute dall’ospite. Prodotti da microrganismi e NON dalle cellule eucariotiche. Conservati tra i microrganismi di una data classe. Essenziali per la sopravvivenza microbica. Prodotti da microrganismi patogeni e NON patogeni (MAMPs: Microbe-Associated Molecular Patterns) Funzione ed espressione trascrizionale diversa dai fattori di virulenza.")
4
Fattori di virulenza PAMPs LPS Flagello CpG DNA PGN PAMPs:
si sono evoluti per fornire funzioni fisiologiche essenziali sono altamente conservati l’espressione è costitutiva Fattori di virulenza: si sono evoluti per permettere un adattamento microbico ad un determinato ambiente differiscono tra i differenti ceppi possono essere inducibili

5
I PAMPs vs i fattori di virulenza
Batteri Presenti su tutti i microrganismi, patogeni e commensali Esclusivamente espressi dai patogeni Espressione Costitutiva Inducibile Localizzazione Cromosomale Generalmente portati da elementi plasmidici o da elementi genetici mobili Funzione Strutturale Interazione con l’ospite Frequenza di mutazione Molto bassa Elevata Grado di conservazione Molto alto tra i membri di una data classe Molto basso

6
PAMPs – ligandi dei PRRs
Componenti della parete cellulare batterica: lipopolisaccaride (LPS), lipoproteine batteriche (BLPs), acido lipoteicoico (LTA), peptidoglicano (PGN), sequenze DNA ricco in guanina e citosina ipo-metilato (CpG) DNA e RNA virale Componenti della parete cellulare dei funghi: zimosani e ife Alcuni costituenti dei Protozoi Elminti e altri costituenti di parassiti? Cellule apoptotiche Ligandi endogeni (DAMPs): HSP60 Auto-antigeni ?
, lipoproteine batteriche (BLPs), acido lipoteicoico (LTA), peptidoglicano (PGN), sequenze DNA ricco in guanina e citosina ipo-metilato (CpG) DNA e RNA virale. Componenti della parete cellulare dei funghi: zimosani e ife. Alcuni costituenti dei Protozoi. Elminti e altri costituenti di parassiti Cellule apoptotiche. Ligandi endogeni (DAMPs): HSP60. Auto-antigeni")
7
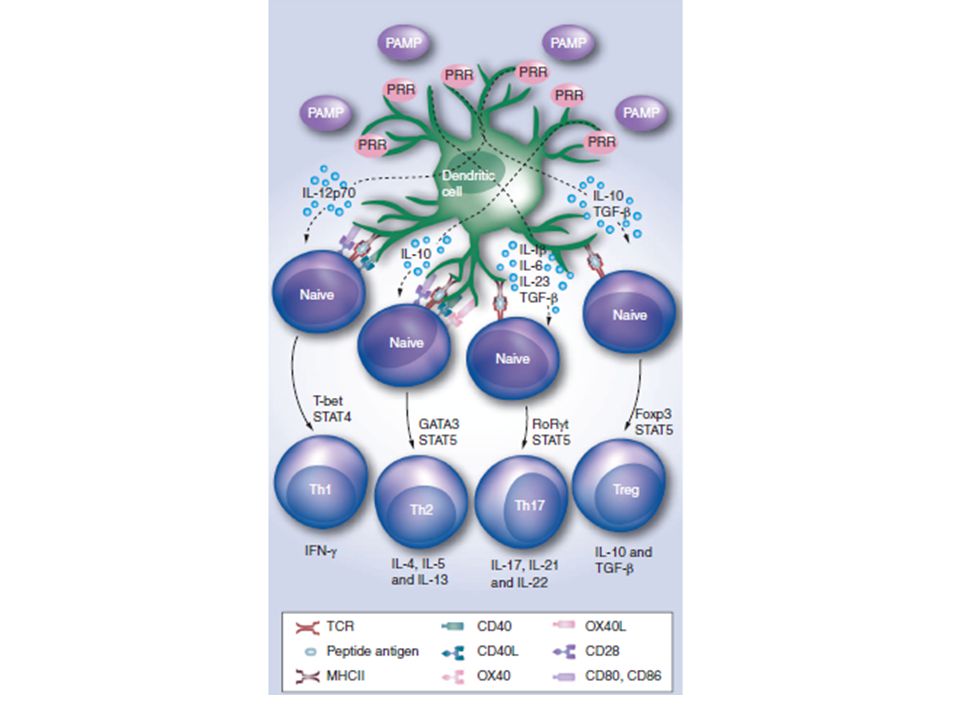
PAMPs-PRRs I PAMPs possono essere riconosciuti da molecole recettoriali dell’ospite (PRRs) In seguito al riconoscimento dei PAMPs mediato dai PRRs diversi tipi cellulari possono portare ad una potente risposta di tipo protettivo che include: Reclutamento di cellule fagocitiche Secrezione di peptidi anti-microbici Sintesi e Rilascio di citochine/chemochine pro- e anti-infiammatorie Istruzione delle cellule dendritiche a promuovere la generazione di una risposta immunitaria adattativa Azione diretta sui linfociti B naive (proliferazione e differenziazione) Azione diretta sui linfociti T effettori e memory L’attivazione dei PRRs è un meccanismo di protezione per l’ospite ma l’over-stimolazione dei PRRs può causare malattie gravi
 Azione diretta sui linfociti T effettori e memory. L’attivazione dei PRRs è un meccanismo di protezione per l’ospite ma l’over-stimolazione dei PRRs può causare malattie gravi.")
9
Le proteine NLRs

10
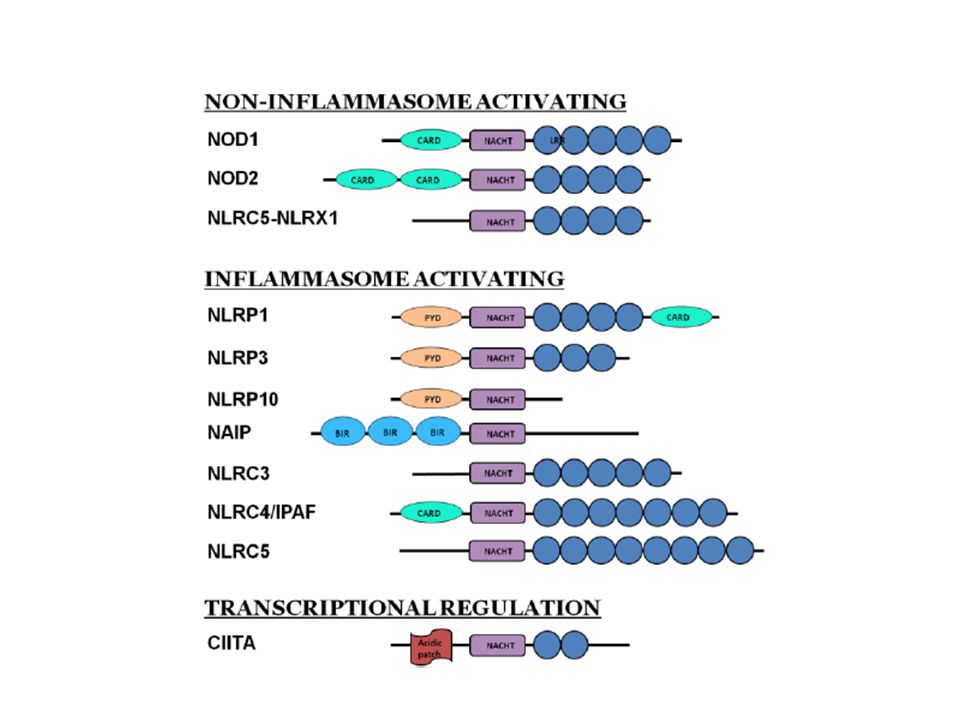
Struttura delle proteine NLRs
Le proteine NLRs presentano un’organizzazione modulare: Dominio amino-terminale EBD [Effector-Binding Domain] Dominio centrale di oligomerizzazione, NOD [Nucleotide-Oligomerization Domain] ora noto come NACHT domain Estremità carbossilica, indicata come LRD [Ligand-Recognition Domain] Le principali differenze tra le proteine NLRs si riscontrano a livello del modulo EBD, la cui funzione sembra essere quella di riconoscere specifici partners molecolari attraverso interazioni omo ed eterofiliche

11
PRRs citosolici: la famiglia delle proteine NLR
Caratteristiche comuni dei membri della famiglia NLR: Dominio C-terminale di ripetizioni ricche in leucina (LRR) coinvolto nel riconoscimento dei ligandi Dominio NACHT che media l’oligomerizzazione e che è essenziale per l’attivazione Dominio effettore N-terminale responsabile dell’interazione proteina-proteina con le molecole adattatrici Kersse et al., 2011
 coinvolto nel riconoscimento dei ligandi. Dominio NACHT che media l’oligomerizzazione e che è essenziale per l’attivazione. Dominio effettore N-terminale responsabile dell’interazione proteina-proteina con le molecole adattatrici. Kersse et al.,")
13
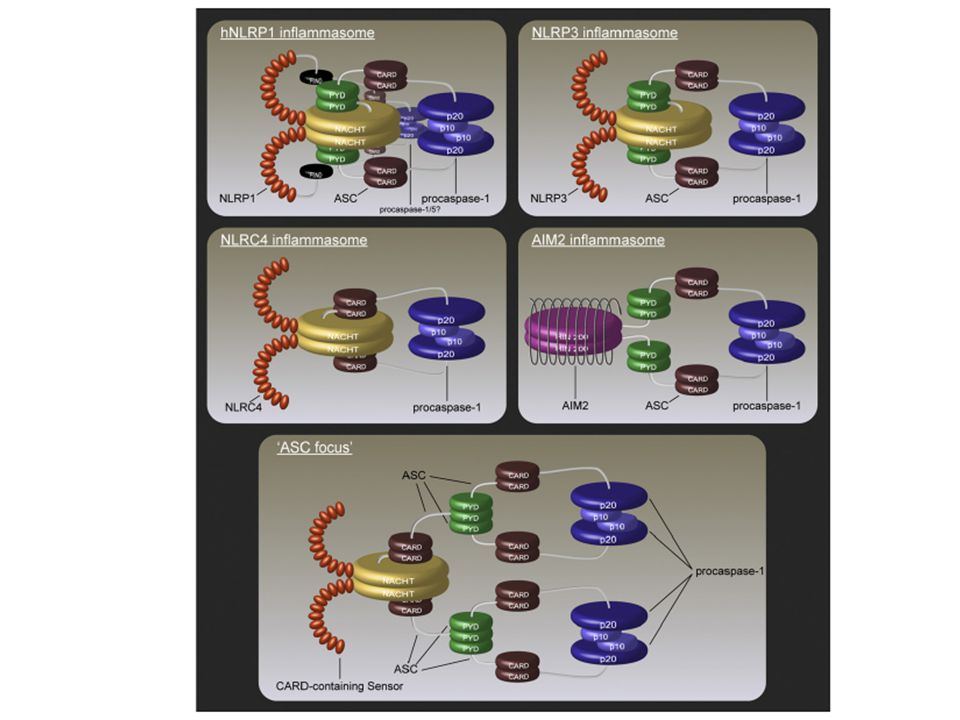
NOD-soma, inflammosoma, apoptosoma…
La diversa associazione di/tra proteine NLR e il reclutamento di molecole effettrici (caspasi, molecole adattatrici, etc ) crea delle piattaforme sensibili ai componenti microbiche e a molecole di pericolo in grado di attivare l’immunità innata
 crea delle piattaforme sensibili ai componenti microbiche e a molecole di pericolo in grado di attivare l’immunità innata.")
15
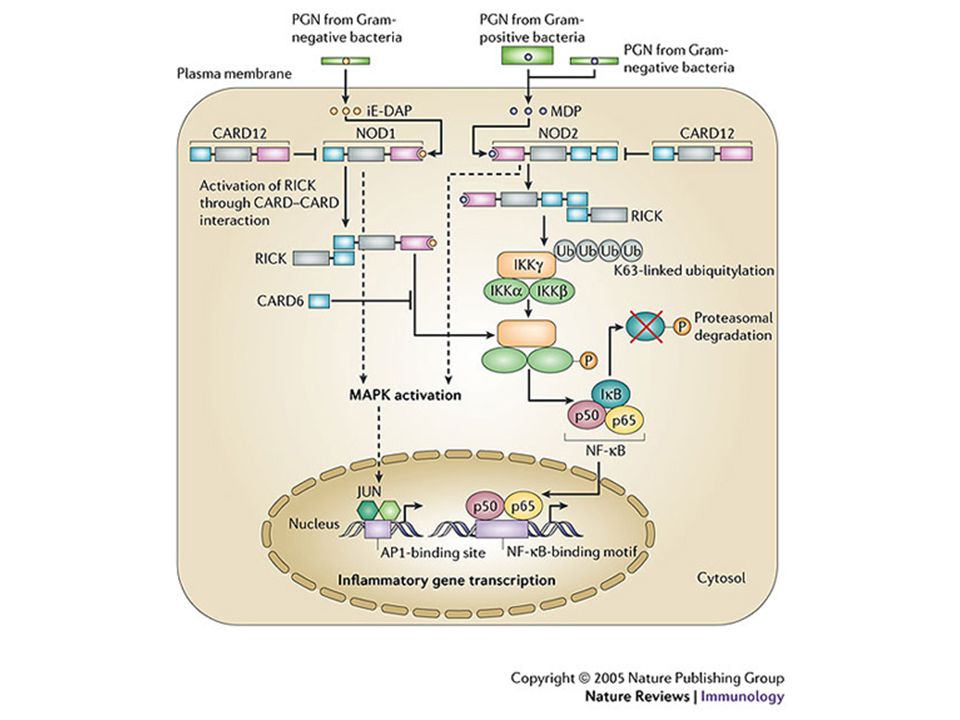
Le proteine Nod1 e Nod2 Funzionano come sensori intracellulari
CARD NOD1 NOD2 NACHT LRR Funzionano come sensori intracellulari Riconoscono frammenti di PGN mediante il dominio LRR

16
Il Peptidoglicano (PGN)
")
17
Quali sono i motivi riconosciuti da Nod1 e Nod2?

19
Quali gli effetti dell’attivazione di Nod1 e Nod2?
Nelle cellule dendritiche, nei macrofagi e nei monociti si ha la produzione di citochine pro-infiammatorie (TNFa, IL-1b, IL-6, CXCL-8, IL-12p70), ossido nitrico (NO), espressione di molecole co-stimolatorie e molecole di adesione Nelle cellule epiteliali si ha la produzione di mediatori pro-infiammatori (TNFa, CXCL-8, MIP2) peptidi antimicrobici (b-defensine)
, ossido nitrico (NO), espressione di molecole co-stimolatorie e molecole di adesione. Nelle cellule epiteliali si ha la produzione di mediatori pro-infiammatori (TNFa, CXCL-8, MIP2) peptidi antimicrobici (b-defensine)")
20
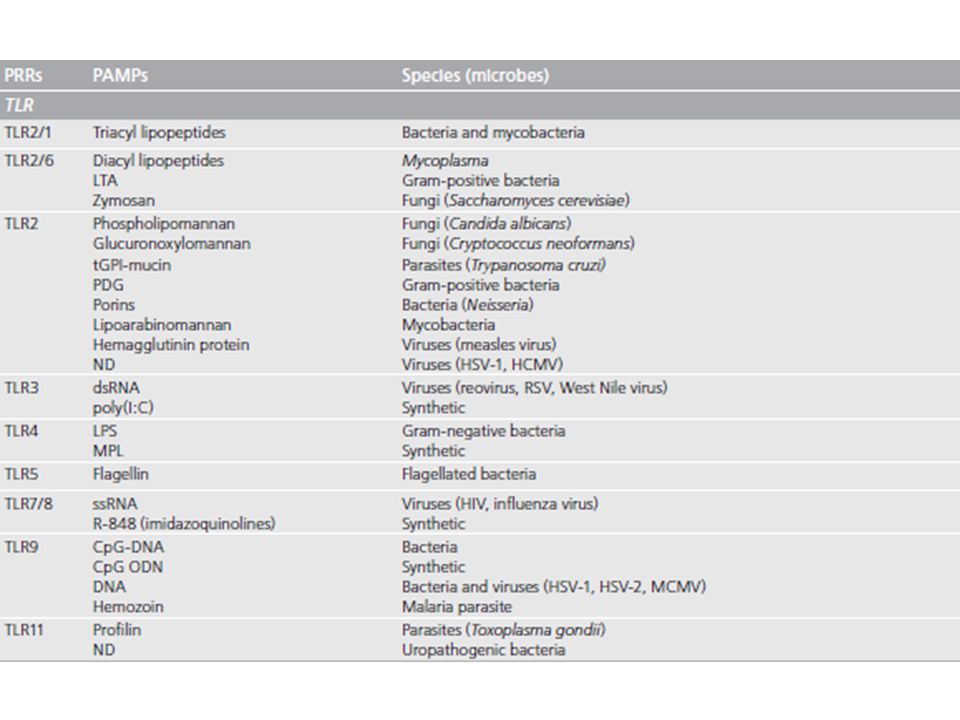
I Toll-like receptors (TLRs)
")
21
I Toll-like receptors (TLRs)
Ad oggi sono noti 13 TLRs: 10 nell’uomo (hTLRs), 13 nel topo (mTLRs) I primi nove TLRs (TLR1-TLR9) mostrano un elevato grado di conservazione strutturale e funzionale tra topo e uomo Con eccezione del TLR8 il cui ligando nel topo non è stato ancora identificato mTLR10 non è funzionale come conseguenza di un’inserzione retrovirale hTLR11 non funzionale a causa di un prematuro codon di stop nella sequenza codificante
, 13 nel topo (mTLRs) I primi nove TLRs (TLR1-TLR9) mostrano un elevato grado di conservazione strutturale e funzionale tra topo e uomo. Con eccezione del TLR8 il cui ligando nel topo non è stato ancora identificato. mTLR10 non è funzionale come conseguenza di un’inserzione retrovirale. hTLR11 non funzionale a causa di un prematuro codon di stop nella sequenza codificante.")
22
I PRRs di membrana: i recettori TLRs

23
I PRRs di membrana: i recettori TLRs

24
Struttura dei TLRs 3 domini: Extracellulare, LRR Transmembrana
Intracellulare, TIR

25
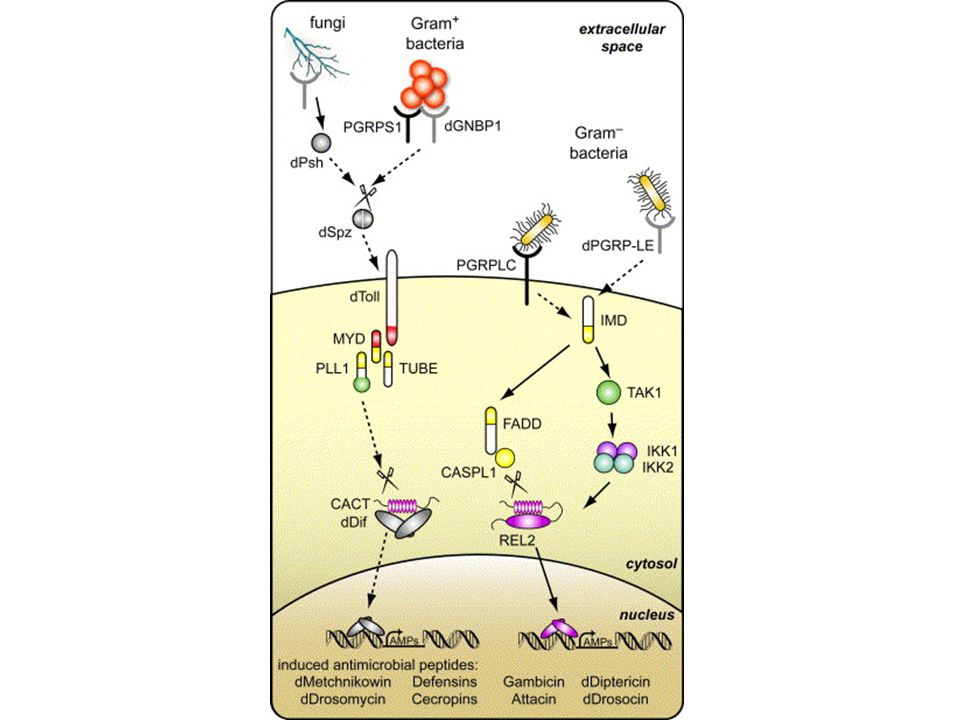
Il Recettore Toll di Drosophila

27
Le mutazioni nel gene Toll determinano la morte del moscerino in seguito a infezioni fungine
Lemaitre et al., Cell 86: 973–983

28
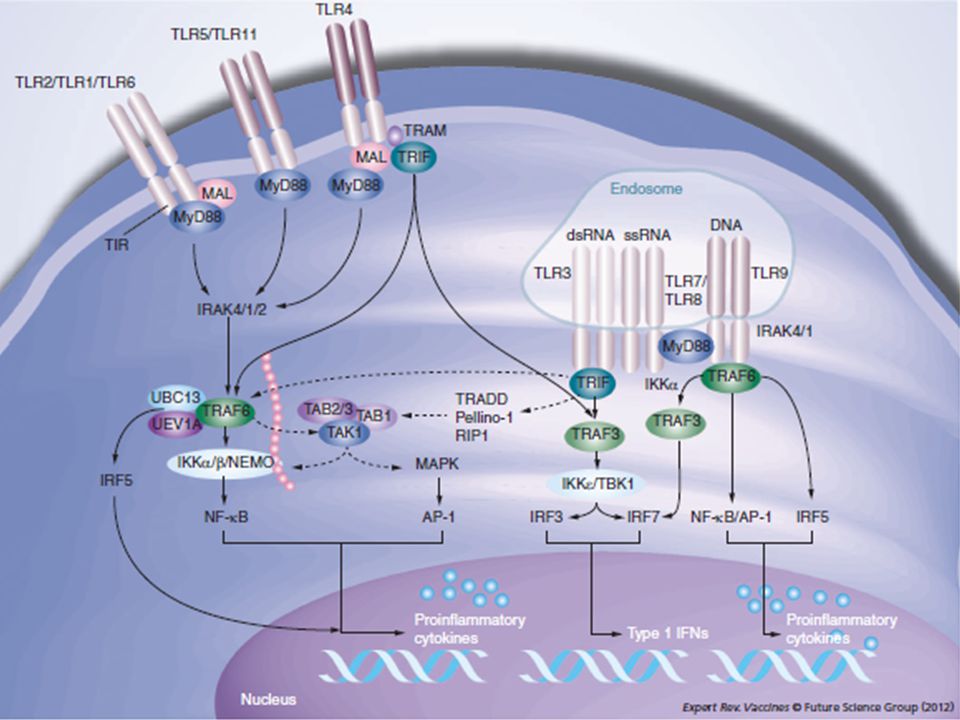
Vie di segnalazione attivate dai TLRs
In seguito a dimerizzazione si hanno DUE vie di segnalazione Via MyD88 dipendente (tutti i TLRs ad eccezione del TLR3): Attivata dalla maggior parte dei TLRs in risposta al legame con un agonista Induce l’attivazione delle MAP chinasi e di NF-kB Attivazione di geni interferonici IRF5 mediati Determina una up-regolazione delle molecole co-stimolatorie e MHC sulla superficie cellulare Via MyD88 indipendente (TLR3 e TLR4): Induce l’attivazione di IRF-3 e la produzione di interferon di tipo I Incrementa le molecole co-stimolatorie e MHC sulla superficie cellulare Induce l’espressione di citochine infiammatorie Induce la maturazione di cellule dendritiche
: Attivata dalla maggior parte dei TLRs in risposta al legame con un agonista. Induce l’attivazione delle MAP chinasi e di NF-kB. Attivazione di geni interferonici IRF5 mediati. Determina una up-regolazione delle molecole co-stimolatorie e MHC sulla superficie cellulare. Via MyD88 indipendente (TLR3 e TLR4): Induce l’attivazione di IRF-3 e la produzione di interferon di tipo I. Incrementa le molecole co-stimolatorie e MHC sulla superficie cellulare. Induce l’espressione di citochine infiammatorie. Induce la maturazione di cellule dendritiche.")
30
TLRs implicati nel riconoscimento di strutture batteriche

31
Il primo TLR ad essere stato individuato mediante omologia di sequenza
Topi C3H/HeJ che presentano una mutazione puntiforme nel dominio TIR di TLR4 sono non-responsivi agli LPS Cosa riconosce? LPS Ligandi endogeni, Heat shock protein, oligosaccaridi di acido ialuronico, fibrinogeno Pneumolisina di S. pneuomoniae

32
Porzione oligo/polisaccaridica
L’LPS: la struttura Regione lipofilica, il Lipide A Regione oligo/polisaccaridica idrofilica legata covalentemente Porzione oligo/polisaccaridica Lipide A Stabilizzazione delle molecole di LPS mediante il legame con cationi divalenti Funzione di barriera impermeabile (Polimixine, poliamine, peptidi e proteine cationiche, agenti chelanti [EDTA])
")
33
Lipide A: struttura delle Enterobacteriaceae
Legame a-glicosidico estere Scheletro di D-glucosamine lo scheletro è costituito da un disaccaride con legame b,1-6 sono presenti due gruppi fosforici (solitamente il Lipide A è monofosforilato) sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi Posizione di attacco della regione polisaccaridica
 sono presenti fino a 4 catene aciliche che a loro volta possono essere sostituite da ulteriori acidi grassi fino a 7 sostituenti acidi. Posizione di attacco della regione polisaccaridica.")
34
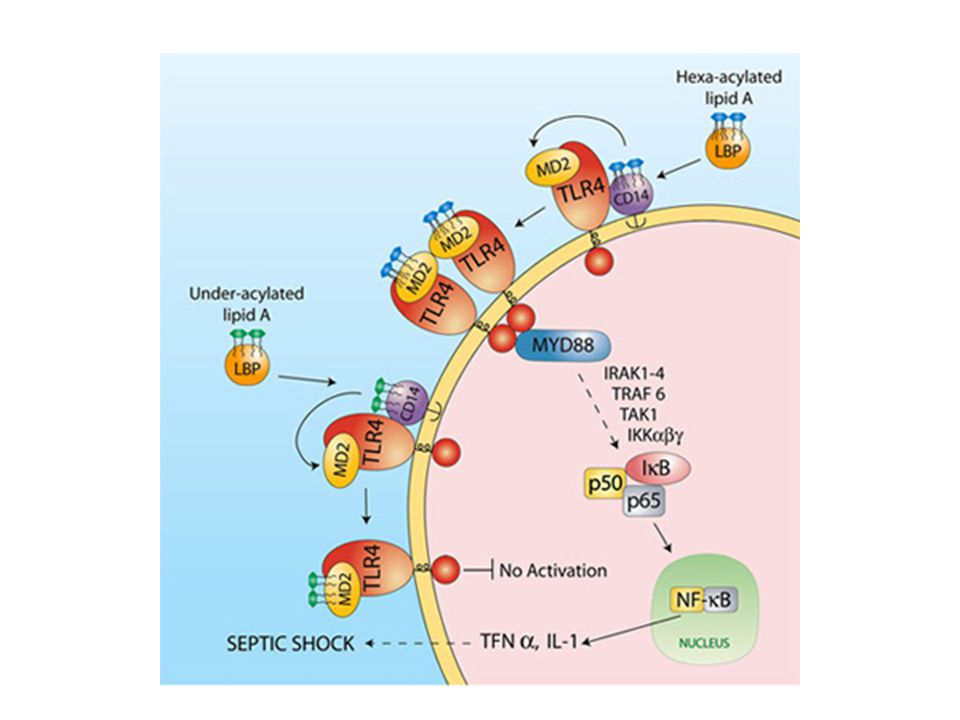
Adattatori molecolari del TLR4
Gli LPS nel siero vengono legati da LPS-binding protein (LBP) e trasportati alle cellule target Le LBP mediano il trasferimento di monomeri di LPS al CD14 che facilita il riconoscimento da parte del TLR4 Per essere attivo il TLR4 necessita di un adattatore, il fattore solubile MD2 Il complesso TLR4/MD2 si forma a livello del reticolo endoplasmatico e viene successivamente traslocato a livello della membrana citoplasmatica
 e trasportati alle cellule target. Le LBP mediano il trasferimento di monomeri di LPS al CD14 che facilita il riconoscimento da parte del TLR4. Per essere attivo il TLR4 necessita di un adattatore, il fattore solubile MD2. Il complesso TLR4/MD2 si forma a livello del reticolo endoplasmatico e viene successivamente traslocato a livello della membrana citoplasmatica.")
35
L’adattatore MD2 Il corecettore MD2 possiede una tasca, una cavità all’interno della quale vengono alloggiate 5 delle 6 catene aciliche del lipideA Negli LPS esacilati, la sesta catena di acido grasso è all’esterno della tasca, a contatto con l’ambiente esterno Le proprietà agoniste o antagoniste dell’LPS sono dettate dalla stechiometria delle catene aciliche con MD2

37
TLR2 Riconosce molteplici componenti batteriche:
Lipoproteine/lipopeptidi Acidi lipoteicoici Peptidoglicano G+ (S. aureus, S. pyogenes, S. pneumoniae) Lipoarabinomannani dei micobatteri Zimosani dei funghi (S. cerevisiae) LPS atipici (Porphyromonas gingivalis, Helicobacter pylori) HSV e VZV (varicella-zooster virus) Come può riconoscere un così vasto spettro di componenti microbici? TLR2 è in grado di formare eterodimeri (con TLR1 o TLR6) Interagisce con altri recettori, per esempio dectin-1 (zimosani) Può interagire anche con il CD14 che induce risposte potenziate verso alcuni PAMPs
 Lipoarabinomannani dei micobatteri. Zimosani dei funghi (S. cerevisiae) LPS atipici (Porphyromonas gingivalis, Helicobacter pylori) HSV e VZV (varicella-zooster virus) Come può riconoscere un così vasto spettro di componenti microbici TLR2 è in grado di formare eterodimeri (con TLR1 o TLR6) Interagisce con altri recettori, per esempio dectin-1 (zimosani) Può interagire anche con il CD14 che induce risposte potenziate verso alcuni PAMPs.")
38
Le lipoproteine

39
lipoproteine Lipoproteine triacilate CH2 CH CH2 S Cys O NH (aa)56
Lys NH2 COOH Lipoproteine diacilate Es: MALP2 (macrophage-activating lipopeptide da Mycoplasma fermentans) LIPOPROTEINA PEPTIDOGLICANO XXXXXXXXXXXXXX GlcNAc MurNAc XXXXXXXXXXXXXXXXXX D-Ala L-Ala D-Glu m-Dap NH2
 LIPOPROTEINA. PEPTIDOGLICANO. XXXXXXXXXXXXXX GlcNAc MurNAc XXXXXXXXXXXXXXXXXX. D-Ala. L-Ala. D-Glu. m-Dap NH2.")
40
Gli eterodimeri di TLR2 TLR1 TLR2 L’eterodimero TLR2/1 riconosce le lipoproteine triacilate L’eterodimero TLR2/6 riconosce le lipoproteine diacilate TLR6 TLR2

41
TLR5 Il motivo batterico riconosciuto è rappresentato da una sequenza altamente conservata tra diverse specie batteriche di 13 amminoacidi della flagellina (D1). Seppur presente nella struttura flagellare, questa sequenza è accessibile al riconoscimento solo quando in forma monomerica. Alcuni batteri patogeni flagellati hanno evoluto dei meccanismi per eludere la risposta mediata dal riconoscimento della flagellina: Helicobacter pylori: presenta una flagellina funzionale per quanto concerne la motilità ma che presenta una sequenza alterata a livello del dominio riconosciuto dal TLR5 Vibrio cholerae e H. pylori: la flagellina è mantenuta strettamente racchiusa nel flagello poiché è ricoperta da un rivestimento membranoso per evitare il rilascio di essa nell’ambiente Il TLR5 interagisce con una specifica conformazione del dominio D1 della flagellina che assume solo quando questa è in forma monomerica Ramos et al., Trends Microbiol. 12: (2004)
. Seppur presente nella struttura flagellare, questa sequenza è accessibile al riconoscimento solo quando in forma monomerica. Alcuni batteri patogeni flagellati hanno evoluto dei meccanismi per eludere la risposta mediata dal riconoscimento della flagellina: Helicobacter pylori: presenta una flagellina funzionale per quanto concerne la motilità ma che presenta una sequenza alterata a livello del dominio riconosciuto dal TLR5. Vibrio cholerae e H. pylori: la flagellina è mantenuta strettamente racchiusa nel flagello poiché è ricoperta da un rivestimento membranoso per evitare il rilascio di essa nell’ambiente. Il TLR5 interagisce con una specifica conformazione del dominio D1 della flagellina che assume solo quando questa è in forma monomerica. Ramos et al., Trends Microbiol. 12: (2004)")
42
TLR9 Riconosce oligonucleotidi CpG ipo-metilati
Questi sono motivi abbondanti nel genoma batterico, mentre nei mammiferi la frequenza è altamente ridotta e in più quando presenti sono altamente metilati Il TLR9 in condizioni fisiologiche si localizza a livello del RE ma agisce a livello endosomiale I batteri in seguito a fagocitosi si ritrovano a livello dell’endosoma dove un ambiente acido e riducente porta alla degradazione del DNA batterico in ssCpG Il TLR9 sembra essere coinvolto nella patogenesi di alcune malattie autoimmuni attraverso il riconoscimento di strutture cromatiniche CpG inducono produzione di IL-6, IL-12 e TNFa e l’attivazione di NF-kB e JNK Alcuni DNA virali (HSV, citomegalovirus) inducono attivazione di TLR9 Plasmodio della malaria induce risposta mediata da TLR9
 inducono attivazione di TLR9. Plasmodio della malaria induce risposta mediata da TLR9.")
43
TLRs implicati nel riconoscimento di strutture virali

44
TLR3 Riconosce dsRNA Il dsRNA è o il genoma virale (reovirus) o è prodotto da molti virus come intermedio del loro ciclo repicativo. I dsRNA inducono la sintesi di interferoni di tipo I (IFN-a/b) ad attività anti-virale e immunostimolatoria TLR7 e TLR8 Presentano un’elevata omologia Riconoscono: ssRNA ricchi in uridina come quelli del virus dell’immunodeficienza umana e del virus dell’influenza composti sintetici quali imidazochinoloni Streptococci di Gruppo B Sono espressi a livello della membrana endosomiale e questo potrebbe spiegare la mancata attivazione da parte degli RNA normalmente presenti nelle cellule
 o è prodotto da molti virus come intermedio del loro ciclo repicativo. I dsRNA inducono la sintesi di interferoni di tipo I (IFN-a/b) ad attività anti-virale e immunostimolatoria. TLR7 e TLR8. Presentano un’elevata omologia. Riconoscono: ssRNA ricchi in uridina come quelli del virus dell’immunodeficienza umana e del virus dell’influenza. composti sintetici quali imidazochinoloni. Streptococci di Gruppo B. Sono espressi a livello della membrana endosomiale e questo potrebbe spiegare la mancata attivazione da parte degli RNA normalmente presenti nelle cellule.")
46
Malattie associate a NOD1 e NOD2
Malattia Mutazioni Caratteristiche NOD1 Infezione di Helicobacter pylori nessuna mutazione trasporto di PGN nelle cellule epiteliali attraverso sistema di secrezione di tipo IV Infezione di Chlamidia pneumoniae nessuna mutazione Attivazione di NF-kB nelle cellule endoteliali Infiammazioni croniche dell’intestino polimorfismi Fattori di rischio per IBD nel dominio LRR Asma polimorfismi Fattori di rischio nel dominio LRR per l’asma NOD2 Morbo di Crohn Arg702Trp; Gly908Arg difetto Leu1007fsinsCys nell’attivazione di NF-kB Blau Syndrome Arg334Trp; Arg334Gln costitutiva attivazione di Leu469Phe NF-kB

47
Morbo di Crohn Processo infiammatorio transmurale e discontinuo
Presenza di granulomi Ascessi fistolosi e aree di ulcerazione Dolori addominali associati a diarrea Ispessimento dell’intestino Febbre Manifestazioni extraintestinali a carico delle articolazioni, della cute, del fegato e degli occhi Nei malati di Crohn si ha un’incidenza dell’80% di un polimorfismo in Nod2

48
Effetti della mutazione Nod2
In vitro: si osserva una diminuzione dell’attivazione di NF-kB In vivo: si osserva una over-espressione di NF-kB con conseguente sintesi e rilascio di citochine infiammatorie Un difetto a livello della segnalazione di Nod2 potrebbe portare ad una proliferazione incontrollata di microorganismi nell’intestino e conseguentemente una prolungata infiammazione (es: assenza di peptidi antimicrobici a livello delle cripte) Nod2 alterato potrebbe non indurre la secrezione di molecole anti-infiammatorie quali IL-10 Alcuni autori riportano un ruolo negativo di Nod2 sul TLR2; in assenza di Nod2 si potrebbe avere una continua stimolazione del TLR2 e quindi un’induzione di molecole infiammatorie
 Nod2 alterato potrebbe non indurre la secrezione di molecole anti-infiammatorie quali IL-10. Alcuni autori riportano un ruolo negativo di Nod2 sul TLR2; in assenza di Nod2 si potrebbe avere una continua stimolazione del TLR2 e quindi un’induzione di molecole infiammatorie.")
49
TLRs Diseases Features Prostate Cancer Systemic Lupus erythematosus
TLR-4 Asp299Gly TLR-4 sequence variant TLR-5 stop codon Thr399Ile TLR-2 TLR-3 Alzheimer’s Disease; Diabetes; Coronary Artery Disease Gene polymorphism is associated with protection from the development of disease Prostate Cancer Systemic Lupus erythematosus Ulcerative colitis/Crohn’s Disease The mutation is increased in patients Atherosclerosis Diabetes The treatment with TLR agonist to animals induced experimental disease model Lee and Hwang, 2006 Molecules and Cells.

Presentazioni simili








 SPECIFICA/SELETTIVA:>")









 Pontieri - Elementi di Patologia Generale (ed. Piccin) Majno-Joris - Cellule,>")
