Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La glicolisi

2
Introduzione Una via metabolica molto antica (il nome, dove avviene e perché) Glucosio + 2 NAD+ + 2 ADP + 2 Pi → 2 piruvato + 2 NADH + 2 H+ + 2 ATP + 2 H2O Universale Specifica di alcuni organismi e di alcuni tessuti Classica via catabolica, ma alcuni suoi intermedi partecipano ad altre vie metaboliche

3
METABOLISMO CELLULARE

4
METABOLISMO CENTRALE DEL CARBONIO
Metabolismo ossidativo Gluconeogenesi Glicolisi

5
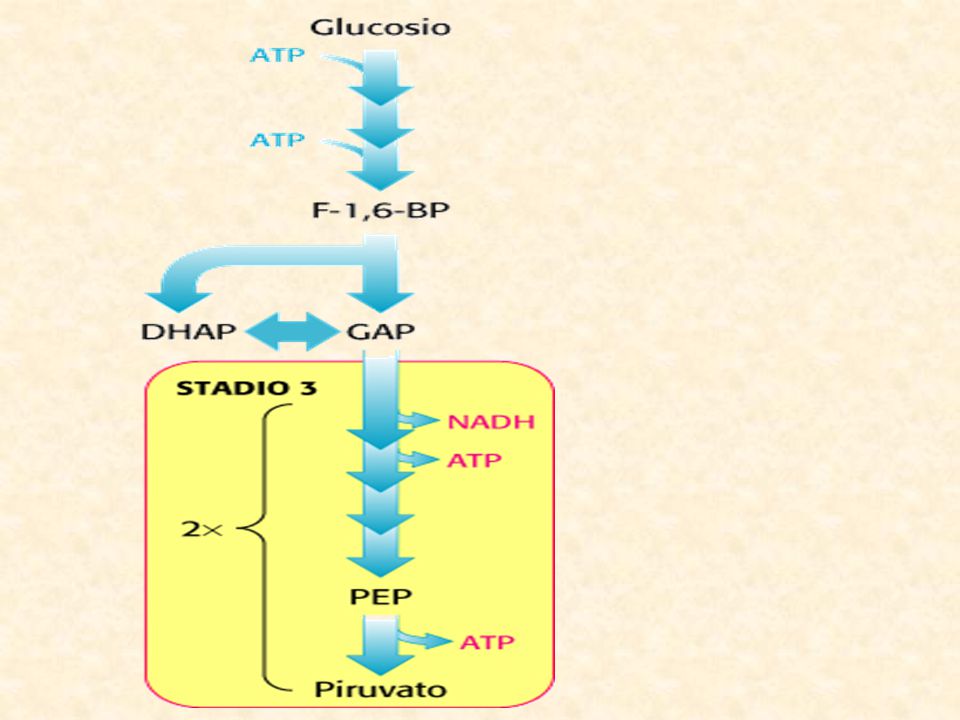
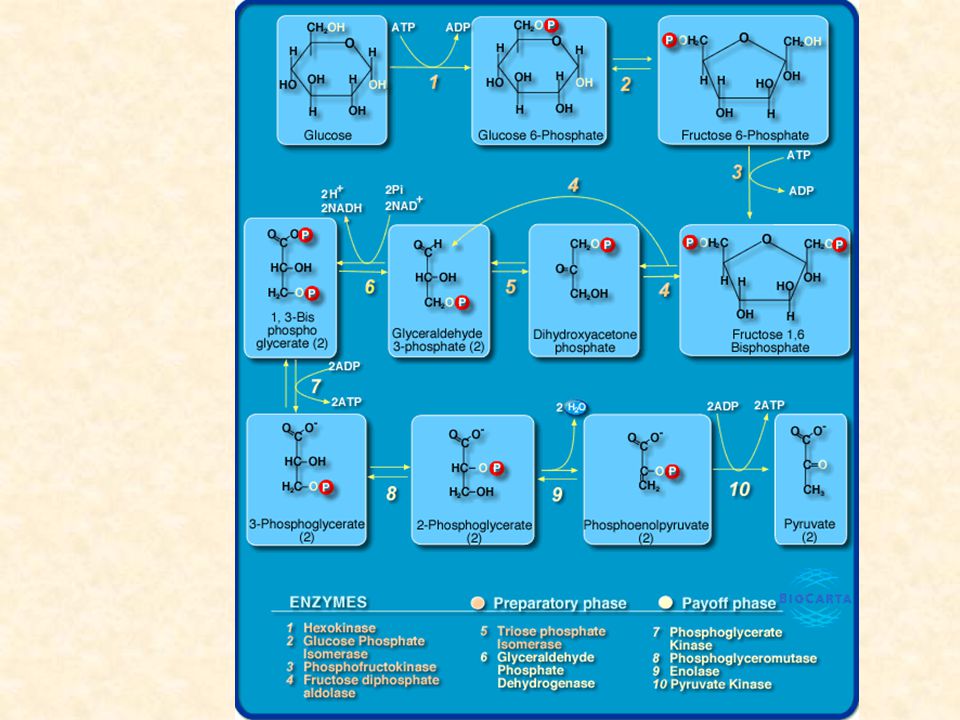
10 reazioni 2 FASI: fase preparatoria, reazioni 1-5 fase del recupero dell’energia, reazioni 6-10 Krebs O2

6
METABOLISMO GLUCIDICO
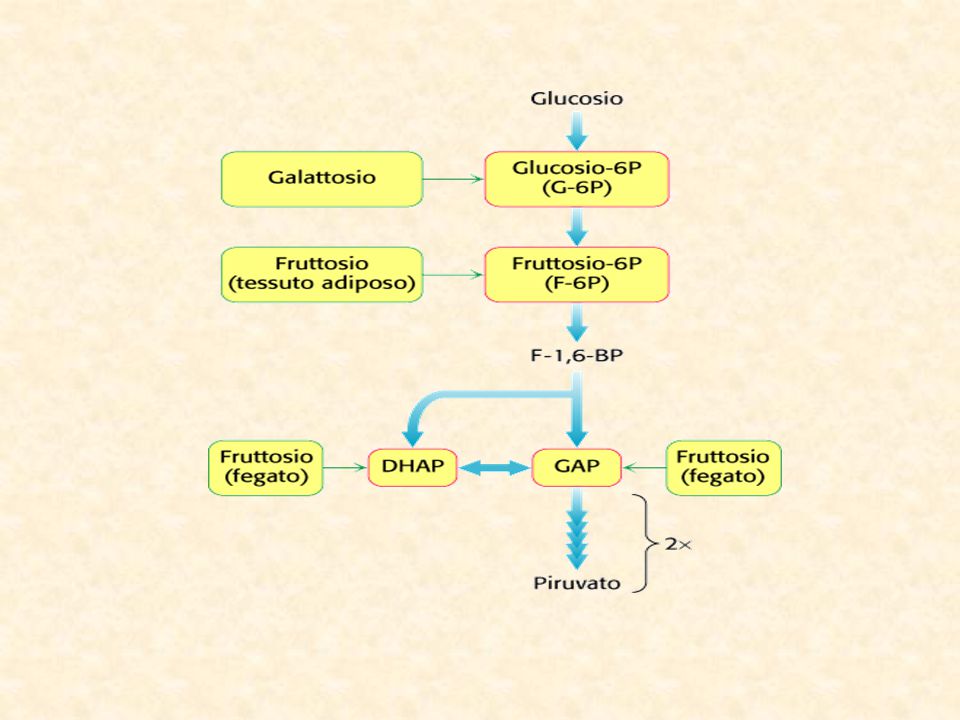
I prodotti finali della digestione dei carboidrati nel tubo digerente sono quasi esclusivamente glucosio, fruttosio e galattosio tra i quali il primo rappresenta circa l’80%. Dopo l’assorbimento intestinale quasi tutto il galattosio e buona parte del fruttosio è trasformato in glucosio. Più del 95% dei monosaccaridi presenti nel sangue sono rappresentati dal glucosio.

7
METABOLISMO GLUCIDICO
Il glucosio, con peso molecolare di 180, non potrebbe attraversare le membrane cellulari (max 100), ma penetra comunque tramite diffusione facilitata, secondo un gradiente di concentrazione e utilizzando diversi carriers come trasportatori. Non c’è quindi in questo caso accoppiamento con il sodio e dispendio quindi energetico. Solo nelle cellule epatiche e cerebrali il glucosio entra in quantità sufficiente al fabbisogno, nelle altre è la presenza di insulina a permetterne un ingresso quantitativamente utile.
, ma penetra comunque tramite diffusione facilitata, secondo un gradiente di concentrazione e utilizzando diversi carriers come trasportatori. Non c’è quindi in questo caso accoppiamento con il sodio e dispendio quindi energetico. Solo nelle cellule epatiche e cerebrali il glucosio entra in quantità sufficiente al fabbisogno, nelle altre è la presenza di insulina a permetterne un ingresso quantitativamente utile.")
8
LA RESPIRAZIONE CELLULARE
La respirazione è un processo di ossidoriduzione nel quale i riducenti sono molecole organiche altamente ridotte e ricche di energia e gli ossidanti sono molecole inorganiche come O2 Il glucosio è il principale combustibile per la maggior parte degli organismi. Il bilancio generale della respirazione può essere riassunto dall'equazione: C6H12O6 + 6 O2 = 6 H2O + 6 CO2 con DG = -686 kcal/mole di glucosio La respirazione avviene in tre stadi principali (stadi della respirazione cellulare); diverse sono anche, per i tre stadi, le sedi della respirazione. Il primo stadio, detto glicolisi, avviene in tutti gli organismi ed è indifferente alla presenza di ossigeno.
; diverse sono anche, per i tre stadi, le sedi della respirazione. Il primo stadio, detto glicolisi, avviene in tutti gli organismi ed è indifferente alla presenza di ossigeno.")
9
LA GLICOLISI PRODOTTI: piruvato, ATP, NADH
La Glicolisi, detta anche via Embden-Meyerhof (Warburg), è il meccanismo più importante di liberazione di energia dal glucosio. Nelle cellule eucariotiche avviene nel CITOPLASMA. Consiste nella scissione di una molecola di glucosio in 2 molecole di acido piruvico (piruvato) con la contemporanea produzione di 4 molecole di ATP, ma una resa netta finale di 2 molecole di ATP. Glucosio + 2ADP + 2NAD+ + 2Pi --> 2 Piruvato + 2ATP + 2 NADH + 2H+ Il processo consiste nella sequenza di 10 reazioni catalizzate da altrettanti enzimi, per ciascuno dei quali il substrato è costituito dal prodotto della reazione precedente. Gli enzimi sono localizzati nel citoplasma, dove sono trattenuti perché in forma fosforilata impermeabile alla membrana, i metaboliti intermedi. PRODOTTI: piruvato, ATP, NADH Il piruvato ha generalmente tre destini diversi
, è il meccanismo più importante di liberazione di energia dal glucosio. Nelle cellule eucariotiche avviene nel CITOPLASMA. Consiste nella scissione di una molecola di glucosio in 2 molecole di acido piruvico (piruvato) con la contemporanea produzione di 4 molecole di ATP, ma una resa netta finale di 2 molecole di ATP. Glucosio + 2ADP + 2NAD+ + 2Pi --> 2 Piruvato + 2ATP + 2 NADH + 2H+ Il processo consiste nella sequenza di 10 reazioni catalizzate da altrettanti enzimi, per ciascuno dei quali il substrato è costituito dal prodotto della reazione precedente. Gli enzimi sono localizzati nel citoplasma, dove sono trattenuti perché in forma fosforilata impermeabile alla membrana, i metaboliti intermedi. PRODOTTI: piruvato, ATP, NADH. Il piruvato ha generalmente tre destini diversi.")
10
Destino del piruvato Per gli organismi anaerobici la glicolisi costituisce il processo energetico fondamentale, il piruvato viene trasformato in acido lattico o alcool etilico Per gli organismi aerobici la glicolisi costituisce la fase anaerobica citoplasmatica preliminare alla fase aerobica mitocondriale in cui l’acido piruvico viene ossidato in CO2 e H2O con la produzione della maggior quota di energia Anche per gli organismi aerobi in assenza di ossigeno la glicolisi rappresenta il solo processo di utilizzazione del glucosio prodotto terminale acido lattico

11
Affinché la glicolisi avvenga il glucosio deve entrare nella cellula
Nella maggior parte delle cellule di mammifero, la concentrazione di glucosio è controllata da TRASPORTATORI DI MEMBRANA soggetti a regolazione (Insulina per Miociti, Adipociti e Neuroni) ed è inferiore alla sua concentrazione ematica 5mM ( mg/100ml) Al contrario il glucosio entra liberamente nelle cellule del Fegato e Pancreas ed in esse la sua concentrazione è simile a quella ematica.
 ed è inferiore alla sua concentrazione ematica 5mM ( mg/100ml) Al contrario il glucosio entra liberamente nelle cellule del Fegato e Pancreas ed in esse la sua concentrazione è simile a quella ematica.")
12
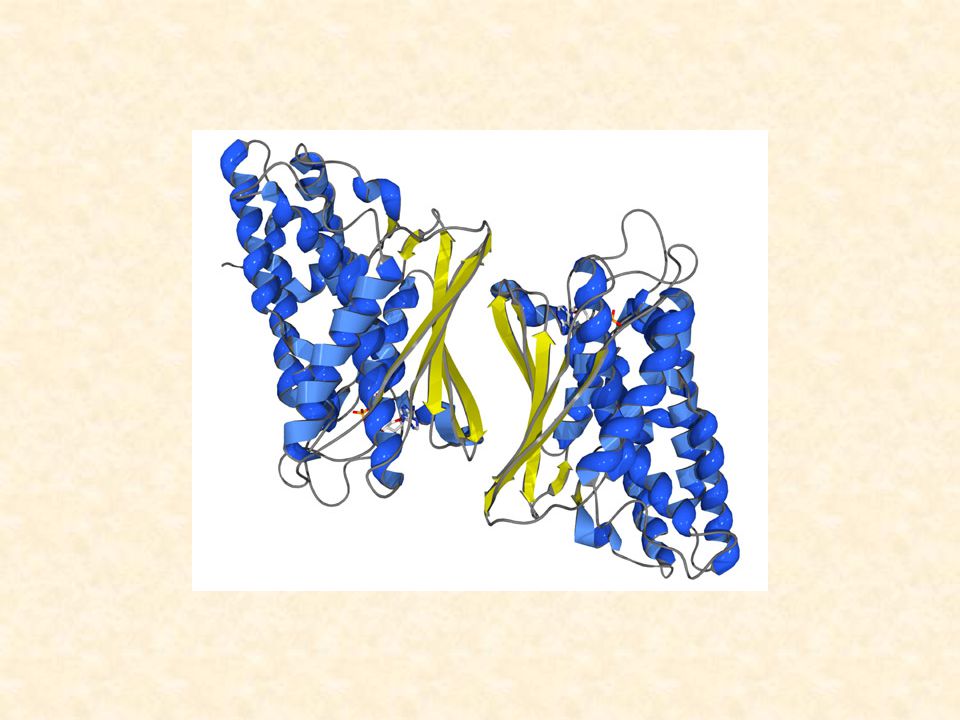
Trasportatore di glucosio nei mammiferi
I trasportatori di glucosio sono membri di una superfamiglia di trasportatori detta superfamiglia dei facilitatori maggiori (MF), che trasportano zuccheri in organismi molto diversi, come Escherichia coli, Trypanosoma brucei (che causa la Tripanosomiasi africana o malattia del sonno africana) e la specie umana.
, che trasportano zuccheri in organismi molto diversi, come Escherichia coli, Trypanosoma brucei (che causa la Tripanosomiasi africana o malattia del sonno africana) e la specie umana.")
13
La famiglia dei trasportatori di glucosio
Nome Localizzazione cellulare Km Commenti GLUT1 Tutti i tessuti dei mammiferi 1mM Captazione basale del glucosio GLUT2 Fegato e cellule del pancreas mM Nel Pancreas svolge un ruolo di regolazione dell’insulina Nel Fegato rimuove dal sangue il glucosio in eccesso GLUT3 Tutti i tessuti dei mammiferi 5 mM La quantità presente nelle membrane plasmatiche delle cellule muscolari aumenta in risposta all’esercizio fisico protratto GLUT5 Intestino tenue 1mM Captazione basale del glucosio GLUT4 Tessuto muscolare e cellule adipose Principalmente un trasportatore di Fruttosio

14
La Glicolisi Stadio 1 Stadio 2 Stadio 3

16
STEP 1: Fosforilazione del glucosio
LE TAPPE DELLA GLICOLISI STEP 1: Fosforilazione del glucosio ESOCHINASI G < 0 G’°= -16,7 KJ/mol Il glucosio-6-P (G6P) può tuttavia formarsi anche dal glucosio-1-P prodotto di degradazione del glicogeno
 può tuttavia formarsi anche dal glucosio-1-P prodotto di degradazione del glicogeno.")
17
Esocinasi Mg++ ΔG'°=-16,7 kJ/mole ΔG=-33,4 kJ/mole

18
Conformazione inattiva
L’esochinasi mostra proprietà che cambiano con il legame del substrato (induced fit ): il legame del glucosio al sito attivo causa variazioni conformazionali. glucoso Conformazione attiva E’ inibita allostericamente dal suo prodotto glucoso-6-P Se il gluc-6P non viene consumato con la stessa velocità con cui è prodotto, inibisce l'esochinasi fino a quando l'eccesso non sia consumato
: il legame del glucosio al sito attivo causa variazioni conformazionali. glucoso. Conformazione attiva. E’ inibita allostericamente. dal suo prodotto glucoso-6-P. Se il gluc-6P non viene consumato. con la stessa velocità con cui è prodotto, inibisce l esochinasi fino a quando l eccesso non sia consumato.")
19
Esochinasi è presente in tutte
le cellule Glucochinasi è presente nel fegato

20
Attraverso la fosforilazione il glucosio si mantiene all’interno della cellula non riuscendo ad oltrepassare la membrana citoplasmatica

21
E’ una reazione di isomerizzazione
STEP 2: Isomerizzazione del glucosio6P in fruttosio6P E’ una reazione di isomerizzazione reversibile Fosfoglucosio isomerasi G’°= 1,7 KJ/mol L’isomerizzazione attiva il C-3 per la scissione nella successiva reazione catalizzata dall’aldolasi

22
Fosfoglucoso isomerasi
ΔG'°=+1,7 kJ/mole ΔG=-2,5 kJ/mole

23
STEP 3: Fosforilazione del fruttosio-6-P
DGo’ = -14,2 kJ/mol Reazione irreversibile Fosfofruttochinasi 1

24
Fosfofruttocinasi 1 Mg++ ΔG'°=-14,2 kJ/mole ΔG=-22,2 kJ/mole

25
- + + FOSFOFRUTTOCHINASI (PFK)
ADP Fru 1,6- bisphoaphate Enzima oligomerico formato da 4 protomeri, a loro volta costituiti da varie subunità Due subunità del tetramero Regolazione allosterica dell’attività enzimatica della PFK FOSFO FRUTTOCHINASI AMP ADP + Pi Fruttosio 2,6 bifosfato Fruttosio 1,6 bifosfato + ATP Citrato - PFK l’attività quando lo stato energetico è basso PFK l’attività quando lo stato energetico è ALTO

26
FOSFOFRUTTOCHINASI (PFK)
Per la molteplicità degli effettori chimici l’enzima è denominato: enzima allosterico plurivalente. Infatti il sito allosterico cui si lega il citrato è distinto da quello per ATP, per cui l’effetto dei due effettori è additivo. Nel fegato la fosfofruttochinasi 1 è inibita anche dagli acidi grassi liberi. Mantenendo [fruttoso 6P] cost. la curva di velocità di sintesi del fruttoso 1,6bifosfato in funzione della concentrazione di ATP mostra un rapido decremento dopo aver raggiunto un valore limite. Quando la [ATP] supera un valore critico si ha INIBIZIONE dell’enzima Curva di saturazione della PFK in funzione della concentrazione di ATP

27
FOSFOFRUTTOCHINASI (PFK)
A basse concentrazioni l’ATP si lega al SITO CATALITICO (> affinità) in forma di complesso con Mg2+ Ad alte concentrazioni si lega al SITO ALLOSTERICO diminuendo l’attività dell’enzima per il fruttoso 6P (< affinità) Ad alte concentrazioni di ATP la cinetica della fosfofruttochinasi muta da iperbolica a sigmoidale A LIVELLO EPATICO il più potente attivatore della PFK è il fruttosio 2,6-bifosfato, la cui azione si manifesta a concentrazioni basse anche 10 volte inferiore agli altri regolatori (ATP;AMP,citrato..)
 in forma di complesso con Mg2+ Ad alte concentrazioni si lega al SITO ALLOSTERICO diminuendo l’attività dell’enzima per il fruttoso 6P (< affinità) Ad alte concentrazioni di ATP la cinetica della fosfofruttochinasi muta da iperbolica a sigmoidale. A LIVELLO EPATICO il più potente attivatore della PFK è il fruttosio 2,6-bifosfato, la cui azione si manifesta a concentrazioni basse anche 10 volte inferiore agli altri regolatori (ATP;AMP,citrato..)")
29
STEP 4: Demolizione dell’esoso in triosi
La reazione è reversibile e all’equilibrio è favorita la formazione di fruttoso 1,6-bifosfato (89% rispetto all’11% dei 2 triosi). Affinché la reazione possa procedere da sinistra a destra uno dei 2 triosi deve essere rapidamente rimosso. DGo’ = -23,9 kJ/mol
. Affinché la reazione possa procedere da sinistra a destra uno dei 2 triosi deve essere rapidamente rimosso. DGo’ = -23,9 kJ/mol.")
30
Aldolasi ΔG'°=+23,8 kJ/mole ΔG= -1,3 kJ/mole

31
Condensazione Aldolica

32
Aldolasi della classe 1 C6 trasformato in 2 C3 (DHAP, Gly-3-P)
Le aldolasi degli animali sono della classe I Queste aldolasi formano basi di Schiff intermedie legate covalentemente tra il substrato e una lisina nel sito attivo dell’enzima

33
STEP 5: Isomerizzazione dei triosofosfati
Diidrossiacetone fosfato e gliceraldeide 3 fosfato sono interconvertibili, in una reazione di isomerizzazione (chetoso) (aldoso) 4% 96% Una reazione redox intramolecolare con meccanismo eno-diolo La reazione procede comunque verso la G3P, in quanto questa ultima viene rimossa molto velocemente dalla reazione successiva. Da 1 molecola di fruttoso1,6 bifosfato, mediante reazioni sequenziali di Aldolasi e triso fosfato isomerasi, si ottengono 2molecole di G3P
 (aldoso) 4% 96% Una reazione redox intramolecolare con meccanismo eno-diolo. La reazione procede comunque verso la G3P, in quanto questa ultima viene rimossa molto velocemente dalla reazione successiva. Da 1 molecola di fruttoso1,6 bifosfato, mediante reazioni sequenziali di Aldolasi e triso fosfato isomerasi, si ottengono 2molecole di G3P.")
34
Trioso fosfato isomerasi
L’enzima è costituita da un nucleo centrale di otto b- foglietti e otto a- elica con regioni di connessione. Questo motivo strutturale, chiamato barile ab, è stato riscontrato anche in uno dei domini della piruvato chinasi Regioni di connessione b-foglietto a-elica

35
Triosofosfato isomerasi
ΔG'°=+7,5 kJ/mole ΔG= + 2,5 kJ/mole

37
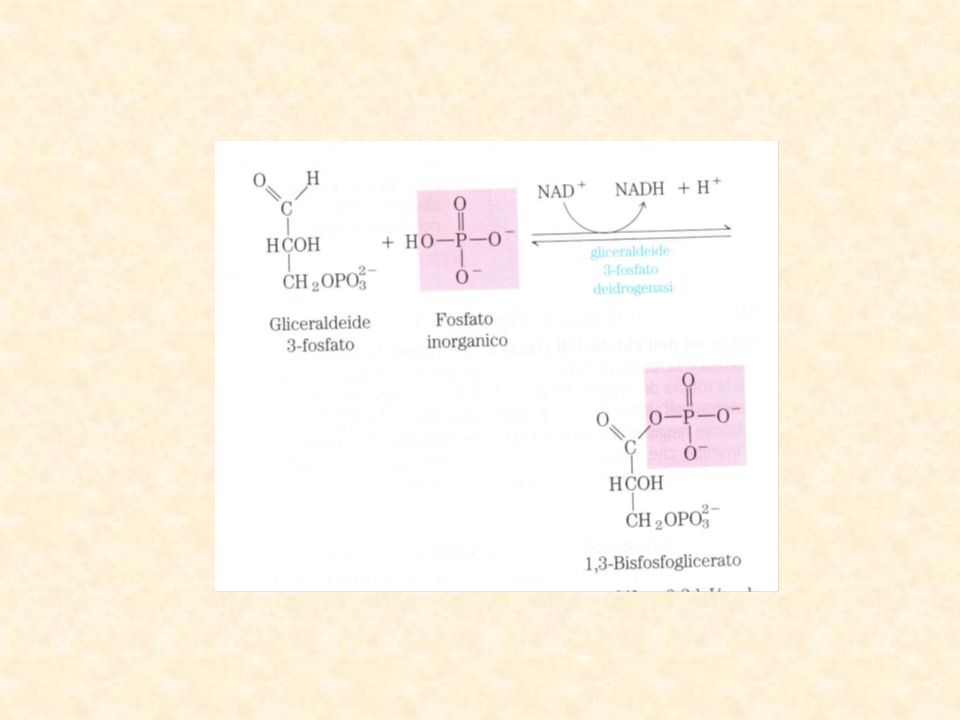
Ossidazione (deidrogenazione) seguita da fosforolisi
STEP 6: ossidazione della gliceraldeide 3 fosfato Ossidazione (deidrogenazione) seguita da fosforolisi DGo’ = +6,3 kJ/mol Legame ad alta energia tra l’acido fosforico e il gruppo carbossilico legame da anidride mista La reazione si dice accoppiata in quanto l’energia derivante dall’ossidazione dell’aldeide viene utilizzata per la formazione del leg.fosfo-anidridico Acil fosfato
 seguita da fosforolisi. DGo’ = +6,3 kJ/mol. Legame ad alta energia. tra l’acido fosforico e il. gruppo carbossilico legame. da anidride mista. La reazione si dice accoppiata in quanto l’energia derivante dall’ossidazione dell’aldeide viene utilizzata per la formazione del leg.fosfo-anidridico. Acil fosfato.")
39
Gliceraldeide-3-fosfato deidrogenasi
ΔG'°=+12,6 kJ/mole ΔG=-3,4 kJ/mole

40
La gliceraldeide 3fosfatodeidrogenasi è un tetramero che puo’ legare 4 molecole di NAD+ una per subunità La sua attività è dipendente dalla integrità di un gruppo –SH in corrispondenza di ogni sito attivo REAZIONE DI ADDIZIONE (1): il gruppo aldeidico della G3P forma con il gruppo SH dell’enzima un TIOSEMIACETALE che viene ossidato dal NAD+ legato all’enzima formando un TIOESTERE (ricco di en.) (3) FOSFOROLISI: Si libera l’acil-fosfato e l’enzima ripristina la sua conformazione iniziale(5)
: il gruppo aldeidico. della G3P forma. con il gruppo SH. dell’enzima un TIOSEMIACETALE che viene ossidato dal NAD+ legato all’enzima formando un TIOESTERE (ricco di en.) (3) FOSFOROLISI: Si libera l’acil-fosfato e l’enzima ripristina la sua conformazione iniziale(5)")
41
Un residuo di cisteina è coinvolto nella catalisi
fosforolisi deidrogenazione

42
Struttura della Gliceraldeide 3-fosfato deidrogenasi
Il polipeptide si ripiega in due domini distinti: Il primo dominio lega il NAD+ Il secondo dominio lega la gliceraldeide 3P

43
Subunità dell’enzima Gliceraldeide - 3 - fosfato deidrogenasi
Dominio a cui si lega la Gliceraldeide fosfato Subunità dell’enzima Gliceraldeide fosfato deidrogenasi

44
STEP 7: 1° Fosforilazione dell’ADP
Sebbene l’equilibrio sia termodinamicamente spostato verso dx, la reazione è tuttavia reversibile consentendo la generazione di ATP nel processo glicolitico: tuttavia quando viene richiesta la sintesi del glucoso (gluconeogenesi)ha luogo la formazione del BPG a spese di ATP DGo’ = -18,9 kJ/mol Fosforilazione a livello del substrato
ha luogo la formazione del 1-3BPG a spese di ATP. DGo’ = -18,9 kJ/mol. Fosforilazione a livello del substrato.")
45
STEP 8: Trasferimento intramolecolare del radicale fosforico
FOSFOGLICERATO MUTASI Scambio reversibile di un gruppo fosfato tra il C-2 e il C-3 del glicerato Mg++ Mutasi: enzimi che catalizzano il trasferimento di un raggruppamento da una parte all’altra della molecola che conserva la propria identità Isomerasi:catalizzano una modificazione di identità del substrato.

46
Il 2-3 bifosfoglicerato cede all’enzima il Pi in posizione 3, formando 2 fosfoglicerato
(reazione 3) Questo gruppo Pi si lega temporaneamente ad un residuo di istidina (His) dell’enzima (generando la forma attiva dell’enz.) per essere poi ceduto al 3 fosfoglicerato ( il substrato della reazione) che si converte in 2,3 bifosfoglicerato e così via FOSFOGLICERATO MUTASI
 Questo gruppo Pi si lega temporaneamente ad un residuo di istidina (His) dell’enzima (generando la forma attiva dell’enz.) per essere poi ceduto al 3 fosfoglicerato. ( il substrato della reazione) che si converte in 2,3 bifosfoglicerato e così via. FOSFOGLICERATO. MUTASI.")
47
Fosfoglicerato cinasi
Mg++ ΔG'°=-37 kJ/mole ΔG =+ 2,7 kJ/mole

48
Il gruppo P scambiato è trasportato temporaneamente da un residuo di His dell’enzima

49
Fosfoglicerato mutasi
Mg++ ΔG'°=+8,8 kJ/mole ΔG= +1,6 kJ/mole

50
La fosfogliceromutasi
richiede quantità catalitiche del 2-3-bifosfoglicerato Gli ERITROCITI hanno un contenuto di 2,3 bifosfoglicerato (4mM) più elevato delle altre, perché lo utilizzano non solo come cofattore nella glicolisi, ma anche come modulatore dell’affinità dell’emoglobina per l’ossigeno. Per questo ricavano il 2,3 BFG dal 1,3 BFG per azione di una MUTASI(bifosfoglicerato mutasi) 1,3 BFG ,3 BFG
 più elevato delle altre, perché lo utilizzano non solo come cofattore nella glicolisi, ma anche come modulatore dell’affinità dell’emoglobina per l’ossigeno. Per questo ricavano il 2,3 BFG dal 1,3 BFG per azione di una MUTASI(bifosfoglicerato mutasi) 1,3 BFG 2,3 BFG.")
51
Il 2,3 BFG può essere defosforilato in 3-fosfoglicerato dalla 2,3 bifosfoglicerato.
La trasformazione del 1,3 bifosfoglicerato in 3 fosfoglicerato per azione successiva di mutasi e fosfatasi implica l’abolizione di formare ATP nella reazione catalizzata dalla fosfoglicerato chinasi. Infatti in base alla richiesta maggiore di ATP o di 2,3-bisfofoglicerato, gli eritrociti utilizzano la normale via metabolica della glicolisi, oppure questo bypass metabolico.. G3P MUTASI 1,3 bifosfoglicerato ADP 2,3 bifosfoglicerato CHINASI ATP FOSFATASI 3 fosfoglicerato Pi

52
STEP 9:Formazione del fosfoenolpiruvato (PEP)
composto ad alta energia DGo’ = 1,8 kJ/mol G0’ per l’idrolisi del fosfato varia da –17.6 kJ/mol nel 2PG a –61.9 kJ/mol nel PEP.

53
STEP 10: 2° Fosforilazione dell’ADP
Piruvico Chinasi Forma chetonica Forma Enolica La seconda fosforilazione dell’ADP a livello del substrato

54
STEP 10: 2° Fosforilazione dell’ADP

55
LA PIRUVICO CHINASI è inibita da alti livelli di ATP(meccanismo allosterico) è attivata da parte del fruttosio-1,6-bifosfato (attivazione feedforward). In tal modo si assicura che il carbonio che fluisce attraverso la glicolisi completi il suo passaggio e non si abbia accumulo di intermedi è inibita dall’acetil-CoA (inibizione da prodotto). Questa inibizione permette di ridurre il flusso attraverso la glicolisi qualora sia disponibile un substrato non zuccherino
. In tal modo si assicura che il carbonio che fluisce attraverso la glicolisi completi il suo passaggio e non si abbia accumulo di intermedi. è inibita dall’acetil-CoA (inibizione da prodotto). Questa inibizione permette di ridurre il flusso attraverso la glicolisi qualora sia disponibile un substrato non zuccherino.")
56
LA PIRUVICO CHINASI E’ presente nei tessuti in tre forme isoenzimatiche: la M nel muscolo e cervello La L nel fegato che è influenzata dalla dieta e dagli ormoni. Il digiuno, condizione che favorisce la gluconeogenesi, ne diminuisce la quantità e il GLUCAGONE che pure stimola la gluconeogenesi, ne diminuisce l’attività stimolando la fosforilazione dell’enzima (Forma fosforilata è meno attiva) L’insulina fa aumentare per induzione,la quantità dell’enzima AZIONE ANTAGONISTA DEL GLUCAGONE E DELL’INSULINA Glucagone + Insulina - La A negli altri tessuti
 L’insulina fa aumentare per induzione,la quantità dell’enzima. AZIONE ANTAGONISTA DEL GLUCAGONE E DELL’INSULINA. Glucagone + Insulina - La A negli altri tessuti.")
57
Sito attivo e regolazione della Piruvico chinasi
Attivato allostericamente da AMP e Fru-1,6-bisP Inibito allostericamente da ATP e acetil-CoA

58
GLUCOSIO 2 ATP 2 NAD+ 2 ADP 2 NADH + 2H+ 2- Gliceraldeide -3- PO4
2 molecole di PIRUVATO 2 ATP 2 ADP 4 ADP 4 ATP 2 NAD+ 2 NADH + 2H+ La glicolisi genera: 2 molecole di ATP e 2 NADH + 2 H+ 2 ATP vengono usati per aggiungere gruppi fosfato al glucosio e al fruttosio -6-P (- 2 ATP) 4 ATP si generano per trasferimento diretto (+ 4 ATP) Glucosio + 2 ADP + 2 Pi + 2 NAD piruvati + 2 ATP + 2 NADH + 2 H+
 4 ATP si generano per trasferimento diretto (+ 4 ATP) Glucosio + 2 ADP + 2 Pi + 2 NAD+ 2piruvati + 2 ATP + 2 NADH + 2 H+")
59
Gli enzimi glicolitici possono formare complessi multienzimatici nelle cellule
Quando le proteine sono purificate da estratti di cellule lisati in soluzioni diluite, le interazioni non-convalenti possono essere distrutte Evidenze cinetiche e fisiche suggeriscono che gli enzimi della glicolisi possono assemblarsi in complessi multienzimatici, dove i prodotti intermedi sono incanalati direttamente da un enzima ad un altro, senza entrare nelle soluzioni acquose, un fenomeno chiamato “substrate channeling”.

60
Gli enzimi glicolitici molto
probabilmente formano complessi multienzimatici.

61
REGOLAZIONE DELLA GLICOLISI
Solitamente le reazioni della glicolisi sono facilmente reversibili, eccetto quelle catalizzate dall'esochinasi, dalla fosfofruttochinasi (PFK) e dalla piruvatochinasi (PK), che fungono da punti di regolazione della glicolisi.(DG < 0 e grande)Il tasso di glicolisi nei mammiferi è controllato proncipalmente a livello della PFK, e parzialmente a livelli dell’esochinasi e della PK. La fosfofruttochinasi, principale elemento di controllo della glicolisi, catalizza una reazione esoergonica irreversibile che permette al glucosio di entrare nella via glicolitica (escludendo il ciclo dei pentosi-fosfati), è attiva quando è necessario produrre energia oppure precursori, ma viene inibita da alte concentrazioni di ATP e di citrato, (CIA) La piruvatochinasi, altro sito di controllo, è inibita allostericamente da Acetil CoA.
 e dalla piruvatochinasi (PK), che fungono da punti di regolazione della glicolisi.(DG < 0 e grande)Il tasso di glicolisi nei mammiferi è controllato proncipalmente a livello della PFK, e parzialmente a livelli dell’esochinasi e della PK. La fosfofruttochinasi, principale elemento di controllo della glicolisi, catalizza una reazione esoergonica irreversibile che permette al glucosio di entrare nella via glicolitica (escludendo il ciclo dei pentosi-fosfati), è attiva quando è necessario produrre energia oppure precursori, ma viene inibita da alte concentrazioni di ATP e di citrato, (CIA) La piruvatochinasi, altro sito di controllo, è inibita allostericamente da Acetil CoA.")
62
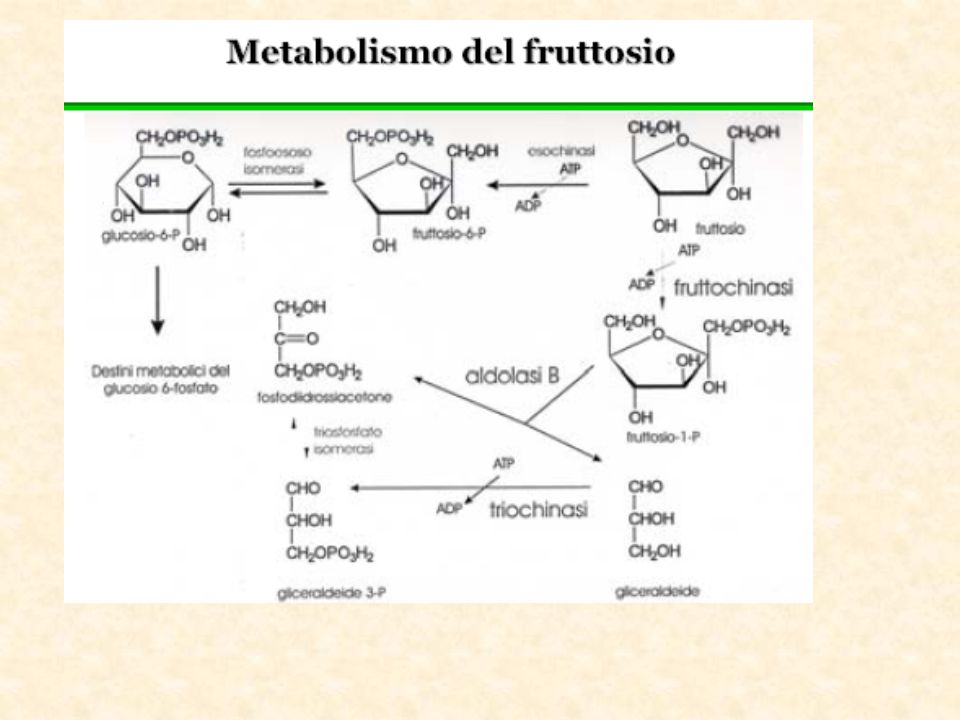
Altri substrati della glicolisi Metabolismo del fruttosio

63
Il fruttosio Il fruttosio (o levulosio) è un monosaccaride contenuto normalmente nella frutta. Presenta basso potere cariogeno, fornisce 4 Kcal /g, e il suo potere edulcorante, circa 1,5 volte superiore a quello del saccarosio, consente un risparmio calorico minimo. Sebbene il metabolismo del fruttosio sia indipendente dall'insulina, quando viene superata la quantità di 40 g/die esso viene trasformato in glucosio. Per questo motivo le persone diabetiche non devono oltrepassare il limite massimo di assunzione giornaliera di 40 g di fruttosio; in questo calcolo va tenuto presente anche la quota presente negli alimenti specifici per diabetici, spesso molto elevata. In quantità elevate il fruttosio può causare diarrea, dolori addominali e flatulenza. Trattandosi di una sostanza naturale, è perfettamente innocua (motivo per cui non ne è stata stabilita la DGA), ma deve essere usato con precauzione nelle persone con alterata funzione renale e gravi disturbi al fegato.
 è un monosaccaride contenuto normalmente nella frutta. Presenta basso potere cariogeno, fornisce 4 Kcal /g, e il suo potere edulcorante, circa 1,5 volte superiore a quello del saccarosio, consente un risparmio calorico minimo. Sebbene il metabolismo del fruttosio sia indipendente dall insulina, quando viene superata la quantità di 40 g/die esso viene trasformato in glucosio. Per questo motivo le persone diabetiche non devono oltrepassare il limite massimo di assunzione giornaliera di 40 g di fruttosio; in questo calcolo va tenuto presente anche la quota presente negli alimenti specifici per diabetici, spesso molto elevata. In quantità elevate il fruttosio può causare diarrea, dolori addominali e flatulenza. Trattandosi di una sostanza naturale, è perfettamente innocua (motivo per cui non ne è stata stabilita la DGA), ma deve essere usato con precauzione nelle persone con alterata funzione renale e gravi disturbi al fegato.")
64
Assorbimento del fruttosio
L’assorbimento del fruttosio avviene per diffusione facilitata ed è mediato dal trasportatore GLUT-5. Il fruttosio viene rapidamente rimosso dal lume intestinale, tuttavia elevati carichi alimentari possono superare la capacità del trasportatore con conseguente incompleto assorbimento e rapida Fermentazione

68
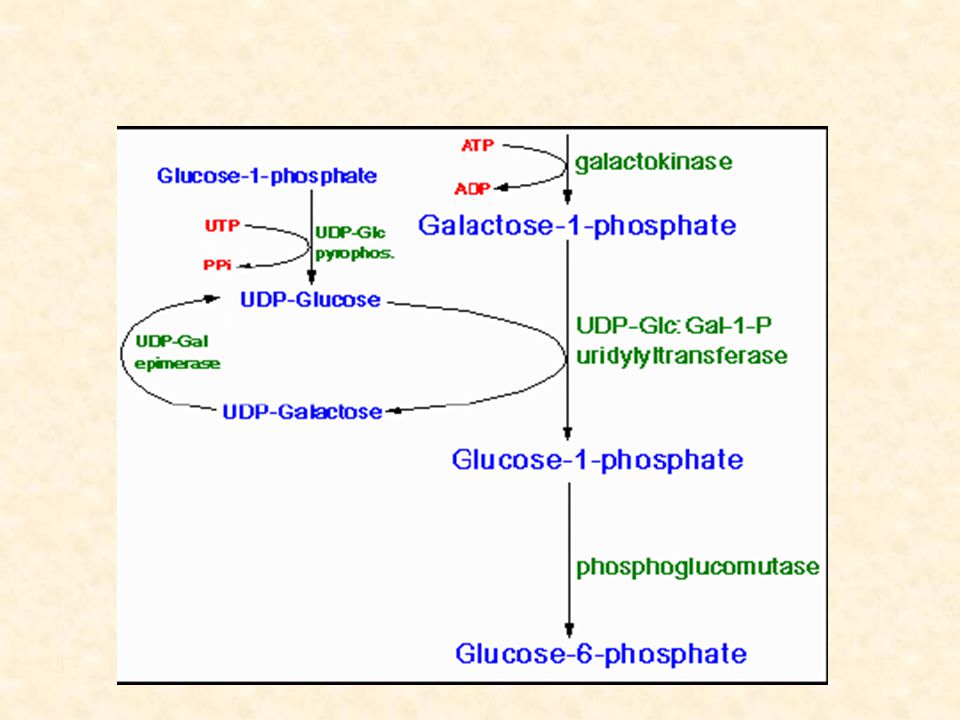
Metabolismo del galattosio

71
Metabolismo del mannosio

72
DESTINO DEL PIRUVATO….. Il piruvato è anche un precursore in
molte reazioni anaboliche. Quando l’ossigeno sarà disponibile, questo debito sarà pagato con la riossidazione del Lattato a Piruvato e con la completa ossidazione di quest’ultimo.

73
Lieviti Eucarioti superiori
L’acido piruvico rappresenta un composto centrale del metabolismo Ha numerosi destini alternativi specie nelle cellule e nei Microorganismi anaerobi. In tutti i casi, è il substrato di eccellenza da ridurre per smaltire l’eccesso di NADH prodotto dalla glicolisi Lieviti Eucarioti superiori

74
Pathways del Piruvato In condizioni ANAEROBIE il piruvato
è convertito in acido lattico o etanolo

75
Fermentazione Lattica
Rigenerazione del NAD+ quando manca O2 (la glicolisi continua comunque) Il NADH proviene dall’OX della G3P Nei muscoli,globuli rossi e nei lattobacilli è molto attiva. Se O2 è disponibile invece, NADH è Ossidato nel trasporto elettronico, producendo ATP nell’ossidazione fosforilativa.
 Il NADH proviene dall’OX della G3P. Nei muscoli,globuli rossi e. nei lattobacilli è molto. attiva. Se O2 è disponibile invece, NADH è. Ossidato nel trasporto elettronico, producendo ATP nell’ossidazione. fosforilativa.")
76
I lattobacilli fermentano il Glucosio trasformandolo in acido lattico.
Il batterio Lactobacillus è anche un componente della flora batterica umana normale del tratto urogenitale dove, grazie alla sua capacità di generare un ambiente acido, impedisce la crescita di batteri nocivi. La straordinaria efficienza della GLICOLISI ANAEROBIA delle Cellule Embrionali e Tumorali spiega la loro capacità a sopravvivere e moltiplicarsi anche in condizioni di scarso apporto di ossigeno.

77
Fermentazione alcolica
Presente solo negli organismi alcool fermentativi Da ox della G3P Presente in molti organismi, tra cui l’uomo La rigenerazione del NAD+ (nella fermentazione lattica o etanolo) mantiene così il flusso di Glicolisi in condizioni anaerobiche
 mantiene così il flusso di Glicolisi in condizioni anaerobiche.")
78
EFFETTO PASTEUR Quando le cellule, capaci di metabolizzare il glucoso sia anaerobicamente che aerobicamente, sono mantenute in condizioni aerobiche, consumano meno glucoso e formano meno acido lattico di quando sono in condizioni anaerobiche. L’osservazione di tale fenomeno oggi spiegabile dalla maggior resa energetica della glicolisi aerobica rispetto a quella anaerobica e dal fatto che in presenza di ossigeno il piruvato non viene ridotto a lattato ma ossidato a CO2 portò Pasteur ad enunciare che “L’OSSIGENO INIBISCE LA GLICOLISI” In condizioni aerobiche la fosforilazione ossidativa mitocondriale del piruvato produce una elevata quantità di ATP. Se si considera che l’aumento di ATP e la concomitante diminuzione di ADP inibisce l’attività della fosfofruttocinasi 1 si comprende come in condizioni aerobiche il flusso glicolitico abbia a diminuire.

79
DESTINO AEROBICO DEL PIRUVATO
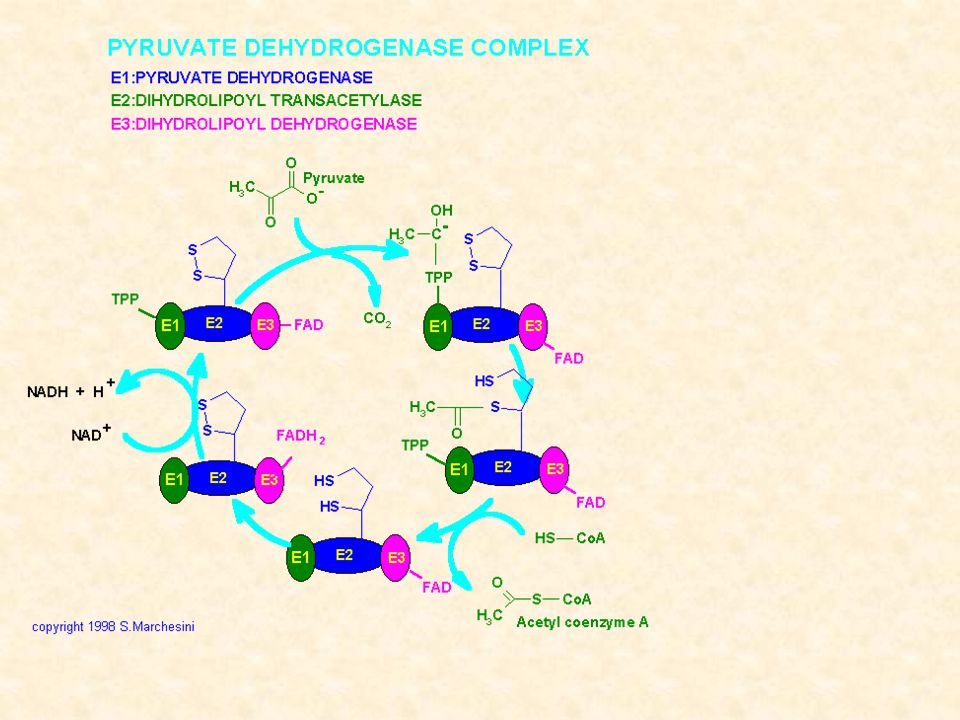
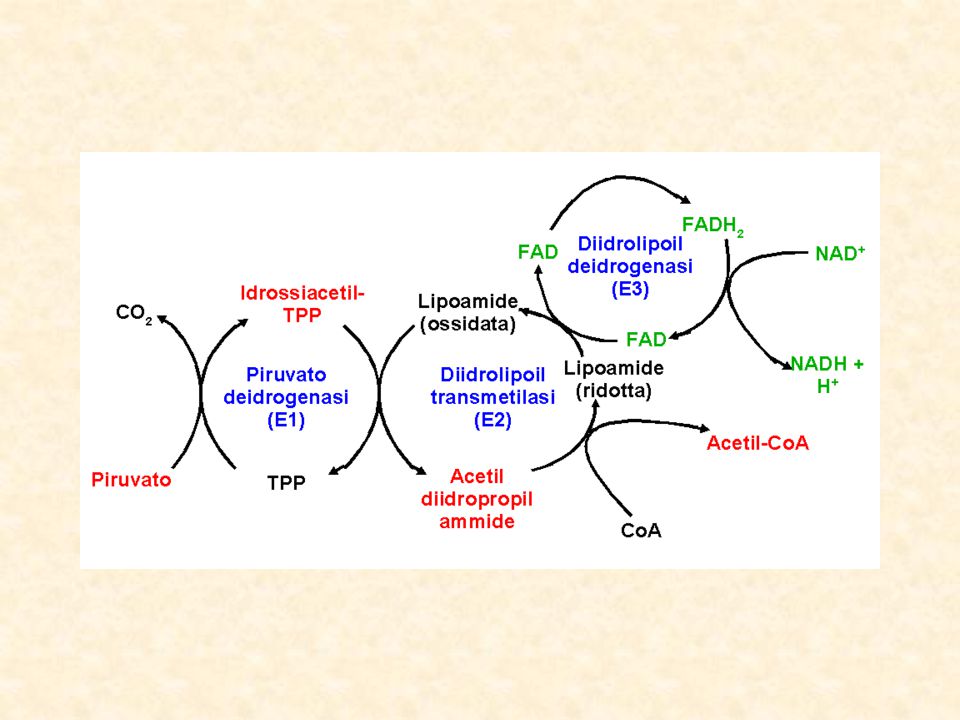
DEIDROGENASI DECARBOSSILAZIONE OSSIDATIVA Il piruvato prodotto dalla glicolisi o dall’ox del lattato passa nei mitocondri attraverso la membrana mitocondriale interna in forma protonata tramite carrier specifico. La PIRUVICO DEIDROGENASI è un complesso enzimatico sito nella porzione più interna della membrana mitocondriale interna e costituito da3 unità: piruvato deidrogenasi, deidrolipoil acetiltransferasi, deidrolipoil deidrogenasi

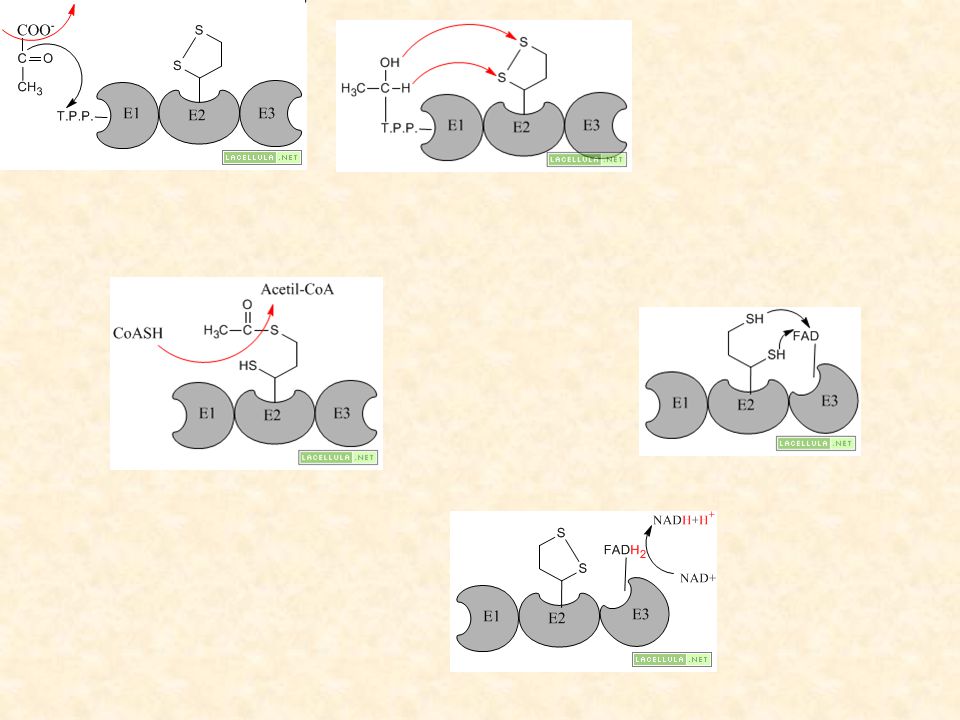
80
PIRUVATO DEIDROGENASI
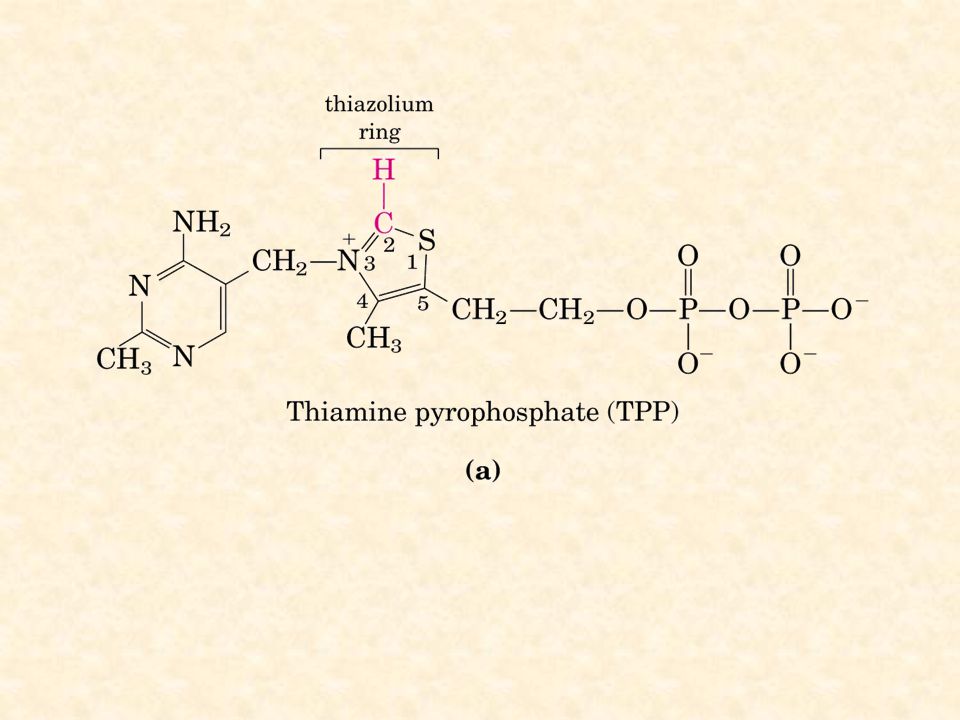
E1 = piruvato deidrogenasi cui si lega al tiamina pirofosfato (TPP) E2 = deidrolipoil acetiltransferasi cui è legato l’acido lipoico E3 = deidrolipoil deidrogenasi cui è legato il FAD
 E2 = deidrolipoil acetiltransferasi cui è legato l’acido lipoico. E3 = deidrolipoil deidrogenasi cui è legato il FAD.")
83
Destino del piruvato…….

85
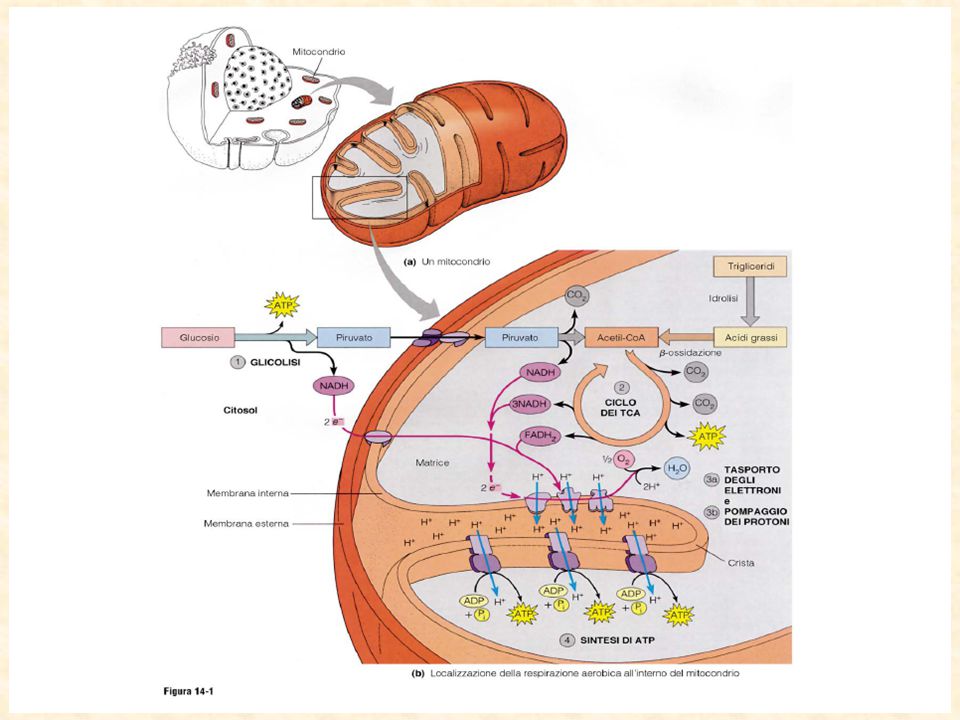
Il piruvato entra nel mitocondrio grazie ad una proteina trasportatrice,per essere decarbossilato ad Acetil CoA ed entrare nel ciclo di krebs

89
ΔG'°=-33,4 kJ/mol

91
REGOLAZIONE DELLA PDH La PDH è controllata mediante inibizione feedback dai suoi prodotti di reazione (acetilCoA e NADH); questo può essere razionalizzato pensando che l’enzima funzioni in modo da produrre acetil-CoA e NADH in modo che vengano utilizzati immediatamente. L’enzima è stimolato da PEP, presumibilmente per segnalare la necessità di degradare il piruvato velocemente ed evitare un intasamento del flusso glicolitico. PDH è stimolata anche da AMP, il cui accumulo indica carenza di ATP.
; questo può essere razionalizzato pensando che l’enzima funzioni in modo da produrre acetil-CoA e NADH in modo che vengano utilizzati immediatamente. L’enzima è stimolato da PEP, presumibilmente per segnalare la necessità di degradare il piruvato velocemente ed evitare un intasamento del flusso glicolitico. PDH è stimolata anche da AMP, il cui accumulo indica carenza di ATP.")
92
+ - + - REGOLAZIONE DELLA PDH FOSFORILAZIONE ad opera di una CHINASI
Forma fosforilata INATTIVA Forma defosforilata ATTIVA PYA= piruvato FOSFORILAZIONE ad opera di una CHINASI Acetil CoA ADP Mg Piruvato NADH NAD+ ATP TPP + - DEFOSFORILAZIONE ad opera di una FOSFATASI Ca NADH Mg Acetil CoA INSULINA + -

93
Funzioni tessuto-specifiche della glicolisi negli animali
Globuli rossi Esclusivamente per energia Muscoli scheletrici Fonte di energia, soprattutto durante lo sforzo intenso Tessuto adiposo Fonte di glicerolo-P per la sintesi di trigliceridi Fonte di acetil-CoA per la sintesi di acidi grassi Fegato

94
IL CICLO DI CORI Nei vari tessuti animali la glicolisi e la gluconeogenesi si svolgono in modo differenziato, nel senso che in alcuni tessuti la glicolisi è molto attiva mentre la gluconeogenesi molto poco. Questo è il caso del muscolo scheletrico. Nel fegato la situazione è opposta. Quando il muscolo lavora in anaerobiosi il lattato formato va in circolo e giunge al fegato dove viene convertito in glucosio, che tramite il circolo giunge al muscolo.

96
I prodotti della glicolisi anaerobica
2 molecole di piruvato 4 molecole di ATP (meno le due usate per l’attivazione del glucosio) 2 molecole di NADH
 2 molecole di NADH.")
97
Destino del piruvato In presenza di ossigeno: il piruvato entra nel mitocondrio, viene trasformato in acetil-CoA che, a sua volta, entra nel ciclo di Krebs. L’ossidazione continua poi in catena respiratoria. In assenza di ossigeno (anossia): nel citoplasma il piruvato può essere convertito in molteplici prodotti finali a seconda degli enzimi presenti; tutte queste reazioni hanno comunque lo scopo di riossidare il NADH, coenzima della Gliceraldeide-3-PDH.
: nel citoplasma il piruvato può essere convertito in molteplici prodotti finali a seconda degli enzimi presenti; tutte queste reazioni hanno comunque lo scopo di riossidare il NADH, coenzima della Gliceraldeide-3-PDH.")
Presentazioni simili




















>")



