Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La razza Maremmana: origini paletnologiche e caratteristiche produttive
Alessandro Giorgetti, Clara Sargentini, Andrea Martini, Roberto Tocci, Ferdinando Ciani

2
Definizione di ‘Razza Podolica’
La Maremmana è generalmente definita razza “podolica”, da Podolia, ampia e fertile pianura dell’Europa orientale, nell’attuale Ucraina. Comprende: razze bovine macrocere italiane, arrivate in Italia a seguito delle invasioni barbariche, caratterizzate da mantello grigio più o meno intenso e cute pigmentata: Maremmana, Romagnola e Podolica, ma anche razze mediocere e brachicere: Chianina, Calvana, Marchigiana, Piemontese, Modenese.

3
Presenze preistoriche di bovini macroceri domestici in Italia
I bovini domestici macroceri però erano presenti nel nostro Paese anche prima della caduta dell’Impero romano. Bovini domestici dalle lunghe corna sono documentati nel nostro Paese fin dal neolitico. Frammenti ossei di bovini domestici con caratteristiche dell’Uro sono stati trovati nel sito delle Arene Candide (SV) del VI millennio a.C., nei siti di Rendina (PZ) e Scamuso (BA), risalenti al VI millennio. Incisioni rupestri in Liguria risalenti all’Età del Bronzo (II millennio a.C.), rappresentano bovini dalle lunghe corna trainanti l’aratro e statuette di bronzo della civiltà nuragica rappresentano bovini macroceri domestici (circa 800 a.C.).
 del VI millennio a.C., nei siti di Rendina (PZ) e Scamuso (BA), risalenti al VI millennio. Incisioni rupestri in Liguria risalenti all’Età del Bronzo (II millennio a.C.), rappresentano bovini dalle lunghe corna trainanti l’aratro e statuette di bronzo della civiltà nuragica rappresentano bovini macroceri domestici (circa 800 a.C.).")
4
Incisioni rupestri della Valle delle Meraviglie (Liguria), VI millennio a. C..
, VI millennio a. C..")
5
Incisioni rupestri del Parco dell’Adamello (età del rame, III millennio a. C.)
")
6
Navicella nuragica, 800 a. C.

7
Presenze di bovini macroceri domestici in Italia in età Etrusca e Romana
Gli Etruschi lasciarono varie testimonianze artistiche di bovini macroceri: tomba dei Tori di Tarquinia ( a.C.) e urna funeraria della tomba del Duce di Vetulonia (VII secolo a.C.). Plinio il Vecchio (I secolo d.C.) descrisse un bovino rustico, dalle grandi corna, diffuso nei boschi e nelle macchie dell’Italia centrale e da lui definito Bos silvestris. Columella (I secolo d.C.) parla di bovini macroceri da lavoro diffusi in varie regioni italiane, la cui descrizione riporta, pur dopo 2000 anni, alle razze macrocere contemporanee (Maremmana, Romagnola, Podolica, Calvana primitiva e in generale tutte le popolazioni e razze grigie italiane). Vari documenti storici, materiale iconografico e reperti archeozoologici testimoniano la presenza nel nostro Paese, fin dai tempi più antichi, di bovini che più tardi saranno chiamati impropriamente “podolici”.
 e urna funeraria della tomba del Duce di Vetulonia (VII secolo a.C.). Plinio il Vecchio (I secolo d.C.) descrisse un bovino rustico, dalle grandi corna, diffuso nei boschi e nelle macchie dell’Italia centrale e da lui definito Bos silvestris. Columella (I secolo d.C.) parla di bovini macroceri da lavoro diffusi in varie regioni italiane, la cui descrizione riporta, pur dopo 2000 anni, alle razze macrocere contemporanee (Maremmana, Romagnola, Podolica, Calvana primitiva e in generale tutte le popolazioni e razze grigie italiane). Vari documenti storici, materiale iconografico e reperti archeozoologici testimoniano la presenza nel nostro Paese, fin dai tempi più antichi, di bovini che più tardi saranno chiamati impropriamente podolici .")
8
Tomba dei Tori di Tarquinia (530-520 a.C.)
")
9
Tomba dei Tori di Tarquinia (altro particolare)
")
10
Tomba del Duce di Vetulonia Vetulonia (VII secolo a.C.).
.")
11
Probabilmente la teoria dell’origine podolica dei bovini macroceri deriva dall’errata interpretazione di un passo dell’ Historia Longobardorum di Paolo Diacono ( d.C.) nella quale si fa riferimento a bovidi dalle grandi corna (che chiamava però Bubalus, bufalo). Sembra invece che la maggioranza dei bovini al seguito dei barbari fossero dei brachiceri. D’altro canto Bodò (1990), sull’origine della Grigia della Puzta, ha evidenziato che fino al Medioevo non esistono descrizioni o documentazioni di macroceri nella popolazione bovina dell’Ungheria, ed ha formulato 2 ipotesi: La popolazione grigia ungherese fu introdotta nel IX secolo d.C. dai conquistatori Magiari provenienti dai Carpazi La popolazione grigia ungherese fu introdotta in Ungheria dall’Europa meridionale (dall’Italia o dalla Penisola Balcanica) nel XIII – XIV secolo. In epoca più recente è certo il trasferimento di Maremmani in Ungheria e, forse anche se in minor misura, di bovini magiari in Italia, particolarmente durante il Granducato dei Lorena in Toscana (1737 –1859).
, sull’origine della Grigia della Puzta, ha evidenziato che fino al Medioevo non esistono descrizioni o documentazioni di macroceri nella popolazione bovina dell’Ungheria, ed ha formulato 2 ipotesi: La popolazione grigia ungherese fu introdotta nel IX secolo d.C. dai conquistatori Magiari provenienti dai Carpazi. La popolazione grigia ungherese fu introdotta in Ungheria dall’Europa meridionale (dall’Italia o dalla Penisola Balcanica) nel XIII – XIV secolo. In epoca più recente è certo il trasferimento di Maremmani in Ungheria e, forse anche se in minor misura, di bovini magiari in Italia, particolarmente durante il Granducato dei Lorena in Toscana (1737 –1859).")
12
La razza Maremmana si può quindi considerare autoctona del nostro Paese e qui presente da tempi preistorici. I bovini rappresentati nell’iconografia etrusca, il Bos silvestris di Plinio e il bovino da lavoro di Columella deriverebbero da popolazioni preesistenti ancora più antiche. Per cercare di ricostruire le reali origini della Maremmana è necessario partire dal progenitore selvatico comune a tutti i bovini: il Bos primigenius o Uro, a sua volta derivato dal pliocenico Leptobos, antenato sia delle diverse specie del genere Bos che delle specie appartenenti al genere Bison.

13
Il progenitore: l’Uro dalle grandi corna
Bos primigenius. Museo di Paleontologia di Roma

14
Berlino. Museo di Storia Naturale

15
L’Uro e le sue sottospecie
L’ Uro è una specie evolutasi in Asia Centrale circa 2 milioni di anni fa; da qui, in epoche successive, avrebbe colonizzato gran parte del continente e avrebbe infine raggiunto l’Europa nell’Holsteniano ( – anni fa), periodo caldo-temperato fra le glaciazioni del Mindel ( – anni fa) e del Riss ( – anni fa). Nell’ultimo periodo interglaciale Riss-Wurm ( – anni fa) e durante l’ultima glaciazione (Wurm, – anni fa), l’Uro avrebbe rivestito un ruolo di primo piano nella sopravvivenza dei popoli cacciatori-raccoglitori del Paleolitico e dell’alto Neolitico. Dal Pleistocene fino alla sua estinzione, avvenuta in tempi storici relativamente recenti in Polonia (1627 d. C.), la specie Bos primigenius raggiunse una notevole diffusione, mantenutasi per diversi millenni. Le sottospecie di Uro, tutte macrocere, riconosciute sono tre: Bos primigenius primigenius, Bojanus 1827, diffuso nella fascia temperata dell’Eurasia; Bos primigenius namadicus, Falconer 1859, distribuito nel subcontinente indiano; Bos primigenius ophistonomus, Pomel 1894, diffusosi in Africa settentrionale.
, periodo caldo-temperato fra le glaciazioni del Mindel ( – anni fa) e del Riss ( – anni fa). Nell’ultimo periodo interglaciale Riss-Wurm ( – anni fa) e durante l’ultima glaciazione (Wurm, – anni fa), l’Uro avrebbe rivestito un ruolo di primo piano nella sopravvivenza dei popoli cacciatori-raccoglitori del Paleolitico e dell’alto Neolitico. Dal Pleistocene fino alla sua estinzione, avvenuta in tempi storici relativamente recenti in Polonia (1627 d. C.), la specie Bos primigenius raggiunse una notevole diffusione, mantenutasi per diversi millenni. Le sottospecie di Uro, tutte macrocere, riconosciute sono tre: Bos primigenius primigenius, Bojanus 1827, diffuso nella fascia temperata dell’Eurasia; Bos primigenius namadicus, Falconer 1859, distribuito nel subcontinente indiano; Bos primigenius ophistonomus, Pomel 1894, diffusosi in Africa settentrionale.")
16
Uro su una pittura rupestre a Lascaux, Francia.

17
L’Uro europeo L’Uro europeo era di grandi dimensioni: i maschi adulti potevano superare l’altezza al garrese di m 1,80 e avevano un peso vivo che poteva oltrepassare i 900 kg. Il colore del mantello variava dal grigio chiaro, al fulvo, al grigio scuro, al bruno, con tonalità scure che aumentavano d’intensità fino al nerastro con l’invecchiamento. La femmina aveva il mantello variabile dal rosso al bruno-grigiastro, mentre i vitelli rimanevano fromentini nel primo anno di vita, carattere conservatosi nelle razze impropriamente definite “podoliche”. Specie caratteristica degli ecosistemi a foresta planiziaria e a pianure con macchie arboreo-arbustive lungo i corsi dei fiumi dell’ Eurasia, il suo comportamento alimentare può essere identificato con quello tipico del selettore intermedio, pascolatore - brucatore, diverso da quello caratteristico della maggioranza delle razze bovine attuali, pascolatrici tipiche, ma simile a quello di razze rustiche macrocere come la Maremmana e la Podolica.

18
Polonia. Monumento all’ultimo Uro

19
L’Uro in Italia La presenza dell’Uro nel nostro Paese è documentata da reperti ossei e iconografici che vanno dal Paleolitico superiore ( – anni fa), attraverso il Mesolitico ( anni fa), fino alla metà del Neolitico (IV millennio a.C.); in questo lungo intervallo temporale gli antichi abitanti dell’Europa ci hanno lasciato numerose raffigurazioni di bovini selvatici. Nel nostro Paese sono ben 11 i siti preistorici che in cui compaiono rappresentazioni di Uro; quasi tutti hanno anche restituito resti ossei di Bos p. primigenius, spesso relativamente abbondanti, a dimostrazione dell’importanza della specie come oggetto di caccia e nell’alimentazione. I reperti hanno evidenziato che gli Uro italici, e più in generale dell’Europa meridionale, erano più piccoli rispetto a quelli delle regioni settentrionali del continente, con arti più lunghi in relazione al peso e mantello più chiaro e meno fitto.
, attraverso il Mesolitico ( anni fa), fino alla metà del Neolitico (IV millennio a.C.); in questo lungo intervallo temporale gli antichi abitanti dell’Europa ci hanno lasciato numerose raffigurazioni di bovini selvatici. Nel nostro Paese sono ben 11 i siti preistorici che in cui compaiono rappresentazioni di Uro; quasi tutti hanno anche restituito resti ossei di Bos p. primigenius, spesso relativamente abbondanti, a dimostrazione dell’importanza della specie come oggetto di caccia e nell’alimentazione. I reperti hanno evidenziato che gli Uro italici, e più in generale dell’Europa meridionale, erano più piccoli rispetto a quelli delle regioni settentrionali del continente, con arti più lunghi in relazione al peso e mantello più chiaro e meno fitto.")
20
L’Uro

21
La domesticazione dell’Uro
I principali centri di domesticazione dell’Uro sono stati identificati nell’Asia sud-occidentale (mezzaluna fertile), in periodi corrispondenti almeno al Neolitico pre-ceramico. Da reperti ossei rinvenuti in siti Natufiani della Palestina, presso le rive del Mediterraneo, è emerso che i bovini furono probabilmente addomesticati nel IX millennio a.C., periodo confermato da altri reperti della stessa specie rinvenuti nell’isola di Cipro. In generale, una grande massa di ritrovamenti suggerisce la diffusione di macroceri domestici nel Neolitico lungo le coste asiatiche, africane ed europee del Mediterraneo, fino all’Europa centrale e settentrionale. Per quanto riguarda i brachiceri si ritiene che abbiano avuto origine da una o più varietà di bovini domestici macroceri e che le varie caratteristiche brachicere si siano sviluppate per l’effetto combinato di modificazioni genetiche spontanee e della selezione artificiale operata dall’uomo, agli inizi del V millennio a.C. in Medio Oriente.
, in periodi corrispondenti almeno al Neolitico pre-ceramico. Da reperti ossei rinvenuti in siti Natufiani della Palestina, presso le rive del Mediterraneo, è emerso che i bovini furono probabilmente addomesticati nel IX millennio a.C., periodo confermato da altri reperti della stessa specie rinvenuti nell’isola di Cipro. In generale, una grande massa di ritrovamenti suggerisce la diffusione di macroceri domestici nel Neolitico lungo le coste asiatiche, africane ed europee del Mediterraneo, fino all’Europa centrale e settentrionale. Per quanto riguarda i brachiceri si ritiene che abbiano avuto origine da una o più varietà di bovini domestici macroceri e che le varie caratteristiche brachicere si siano sviluppate per l’effetto combinato di modificazioni genetiche spontanee e della selezione artificiale operata dall’uomo, agli inizi del V millennio a.C. in Medio Oriente.")
22
Testa di Toro. Periodo Minoico/Cretese 1700-1450 a.C.

23
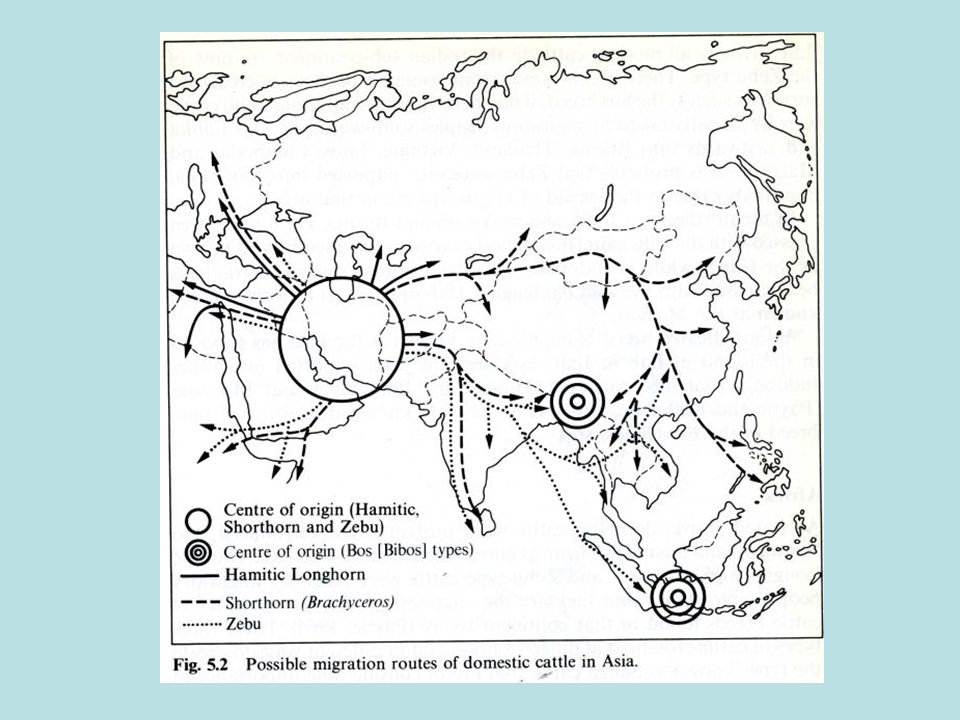
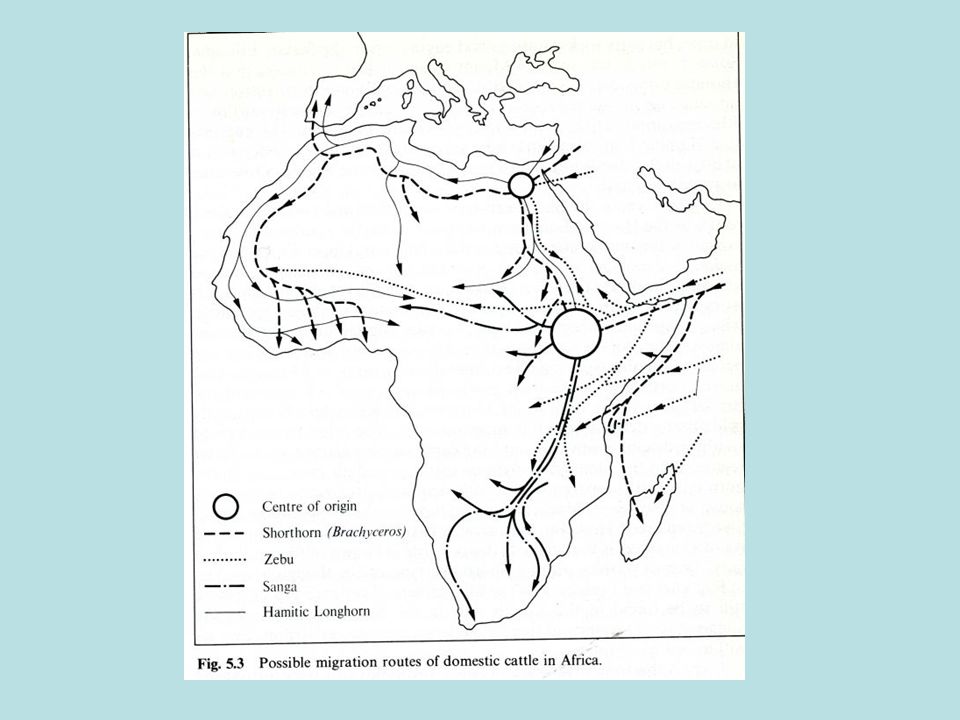
La diffusione di bovini macroceri
I bovini domestici macroceri e brachiceri (in un secondo tempo) si diffusero progressivamente in Europa (VI/VII millennio a.C.) e nel continente africano (V millennio a.C.). In Africa il bovino domestico dalle grandi corna per molto tempo costituì l’unico tipo di bovino presente, ampiamente raffigurato dagli Egizi soprattutto a partire dal III millennio a.C. Le prime immagini del dio Apis illustrano chiaramente il colore del mantello e alcune caratteristiche somatiche di questo bovino primitivo di medie o grandi dimensioni corporee.
 si diffusero progressivamente in Europa (VI/VII millennio a.C.) e nel continente africano (V millennio a.C.). In Africa il bovino domestico dalle grandi corna per molto tempo costituì l’unico tipo di bovino presente, ampiamente raffigurato dagli Egizi soprattutto a partire dal III millennio a.C. Le prime immagini del dio Apis illustrano chiaramente il colore del mantello e alcune caratteristiche somatiche di questo bovino primitivo di medie o grandi dimensioni corporee.")
25
Il dio Apis

26
La diffusione di bovini macroceri
Dall’Egitto verso a.C., gli allevatori di questo ceppo iniziarono una migrazione seguendo tre direttrici: verso sud, in direzione della Nubia e dell’altipiano etiopico verso sud-ovest, in direzione del lago Chad e verso ovest lungo la costiera mediterranea, in direzione delle attuali Libia, Tunisia, Algeria e Marocco. Le popolazioni di quest’ultima direttrice, giunte all’altezza dello stretto di Gibilterra, si divisero in due tronconi: uno diretto a sud, lungo la costa atlantica fino al golfo di Guinea (origine delle razze N’dama e Kouri), e l’altro a nord verso la penisola iberica (origine razze iberiche macrocere).
, e l’altro a nord verso la penisola iberica (origine razze iberiche macrocere).")
28
Macroceri europei: l’origine della Maremmana
Il popolamento di Bos primigenius taurus domestico in Europa è complesso: alle popolazioni arrivate durante il Neolitico, attraverso la Turchia e i Balcani, a partire dal VI-VII millennio a.C., si andarono più tardi ad aggiungere le popolazioni iberiche, a loro volta provenienti dall’Egitto e transitate lungo la sponda meridionale del Mediterraneo. E’ anche probabile un’introduzione, via mare, di quelli stessi bovini domestici macroceri in migrazione dall’Egitto. In Europa, agli inizi del 3000 a.C., venivano già allevati bovini brachiceri in associazione con esemplari di maggiori dimensioni di tipo macrocero. Non è chiaro se e in quale misura le diverse popolazioni bovine domestiche macrocere arrivate in Europa siano venute tra loro in contatto, e di conseguenza non è chiaro se le popolazioni centro-europee e soprattutto quelle italiane siano il risultato di una o più di tali introduzioni.

29
Macroceri europei: l’origine della Maremmana
Come accertato da molti siti neolitici del nostro Paese, erano presenti in Italia almeno anni prima della migrazione dall’Africa alla penisola iberica e almeno anni prima che la specie domestica mettesse piede in Egitto. Tutte indicano che i bovini allevati per il lavoro e per le attività ludiche dalle popolazioni pre-celtiche, villanoviane, etrusche, e nuragiche, fino dagli albori, avevano le corna lunghe ed una una somiglianza somatica molto accentuata fra di loro.

30
Macroceri europei: l’origine della Maremmana
Quali rapporti esistono tra l’Uro e i macroceri domestici europei? Sicuramente i secondi sono i discendenti del primo, ma gli Uro erano distribuiti ampiamente in quasi tutto il territorio eurasiatico, con notevoli differenze fenotipiche. Di norma viene accettata l’idea che il centro di domesticazione (o i centri) siano localizzati nella mezzaluna fertile. Le popolazioni domestiche arrivate in Europa già nel neolitico antico trovarono però uro già presenti nel continente da decine di migliaia di anni. Ma, gli Uro europei contribuirono o no alla formazione delle razze domestiche macrocere?
 siano localizzati nella mezzaluna fertile. Le popolazioni domestiche arrivate in Europa già nel neolitico antico trovarono però uro già presenti nel continente da decine di migliaia di anni. Ma, gli Uro europei contribuirono o no alla formazione delle razze domestiche macrocere")
31
Macroceri europei: l’origine della Maremmana
Ricerche condotte alcuni anni fa sulle sequenze di frammenti di DNA mitocondriale di due uro preistorici dell’Inghilterra, avevano dimostrato una netta diversità tra questi e tutte le razze bovine moderne europee. Se le sequenze degli uro inglesi si potessero considerare come rappresentative della composizione genetica della specie selvatica in Europa centro-settentrionale emergerebbe che questa popolazione non ha contribuito alla formazione delle attuali razze bovine europee. Gli allevatori di bovini del neolitico non sarebbero perciò mai stati interessati) ad introdurre nei loro allevamenti il genoma dei locali uro comunemente presenti allo stato selvatico.
 ad introdurre nei loro allevamenti il genoma dei locali uro comunemente presenti allo stato selvatico.")
32
L’albero filogenetico mostra le distanze tra le razze “ gibbute” (Bos indicus), le razze “taurine” (Bos taurus) e l’uro (Bos primigenius) (Troy et al., 2001).
, le razze taurine (Bos taurus) e l’uro (Bos primigenius) (Troy et al., 2001).")
33
Principali corridoi di migrazione delle razze bovine, suggeriti dall’analisi delle sequenze mitocondriali. L’area racchiusa all’interno del cerchio rappresenta il centro d’origine dal quale si sarebbero poi diffuse le razze “taurine”domestiche europee e nord-africane. Il quadrato corrisponde ad un secondo evento di domesticazione, all’interno del quale si colloca l’origine delle razze zebù asiatiche. Il collegamento tra i due centri è abbastanza incerto.

34
Macroceri europei: l’origine della Maremmana
Studi condotti su frammenti ossei di uro italiani (circa anni fa), comparati con le razze Chianina e Maremmana, indicherebbero che alcune sequenze di DNA mitocondriale osservate in queste razze bovine europee attuali erano già presenti negli antichi uro della penisola italiana. Mentre ulteriori studi hanno indicato che, almeno per quanto riguarda le popolazioni nord-europee di uro, non vi fu alcuna partecipazione alla formazione delle moderne razze.
, comparati con le razze Chianina e Maremmana, indicherebbero che alcune sequenze di DNA mitocondriale osservate in queste razze bovine europee attuali erano già presenti negli antichi uro della penisola italiana. Mentre ulteriori studi hanno indicato che, almeno per quanto riguarda le popolazioni nord-europee di uro, non vi fu alcuna partecipazione alla formazione delle moderne razze.")
35
L’albero neighbour joining (NJ) dei bovini antichi e moderni:
Il cerchio rosso rappresenta i Bos primigenius inglesi, quello viola i B. primigenius italiani e B. taurus europei, quello blu i B. taurus africani.

36
Macroceri europei: l’origine della Maremmana
Ricerche più recenti su frammenti di DNA mitocondriale di uro dell’Europa continentale, settentrionale e occidentale hanno infatti dimostrato la presenza dominante di un aplogruppo denominato P che è invece assente nelle razze e popolazioni bovine moderne, sia europee che nord-africane e medio-orientali. le razze europee sono caratterizzate quasi esclusivamente da un singolo gruppo di sequenze del DNA mitocondriale, l’aplogruppo T3; le nordafricane e medio orientali dalla presenza di quattro aplogruppi principali: T, T1, T2, T3, tutti nettamente divergenti dal P. Ciò conferma che, almeno per quanto riguarda le popolazioni nord-europee di uro, non vi fu alcuna partecipazione alla formazione delle moderne razze. Esemplari di uro dell’Italia centrale e meridionale risalenti ad un periodo compreso fra e anni a.C hanno evidenziato nel DNA mitocondriale di questi animali sequenze caratteristiche dell’aplogruppo T3, che li distingue geneticamente dagli uro centro-europei (che presentano l’aplogruppo P) e li avvicina sia alle moderne razze bovine che agli uro del medio-oriente, che hanno lo stesso aplogruppo T3.
 e li avvicina sia alle moderne razze bovine che agli uro del medio-oriente, che hanno lo stesso aplogruppo T3.")
37
European spread of agropastoralism
European spread of agropastoralism. Each black cattle figure represents a population sample point. Different hypothesized maritime routes (dashed line with arrow) and continental route (solid line with arrow) are indicated. Dash-dot lines are suggestive of the geographic limits of African cattle influence in Europe. Pie chart represents the frequencies of the four major mtDNA haplogroups, with circle sizes proportional to sample sizes.
 and continental route (solid line with arrow) are indicated. Dash-dot lines are suggestive of the geographic limits of African cattle influence in Europe. Pie chart represents the frequencies of the four major mtDNA haplogroups, with circle sizes proportional to sample sizes.")
38
Macroceri europei: l’origine della Maremmana
Le attuali razze grigie autoctone italiane, compresa la Maremmana, presentano l’aplotipo T3, comune a quello degli uro centro-meridionali italiani, in circa il 60% degli individui esaminati. Di conseguenza, si può ipotizzare che in Italia gli allevatori neolitici abbiano accettato o addirittura favorito moderati livelli di introgressione genetica di uri selvatici nelle loro popolazioni domestiche, almeno da parte di femmine di Bos primigenius primigenius, pur conservando un pool genetico assai simile a quello di alcune popolazioni medio-orientali. Infatti ulteriori ricerche condotte ancora sul DNA mitocondriale delle razze autoctone toscane Maremmana, Chianina e Calvana e di attuali popolazioni dell’Anatolia e del Medio Oriente, confermano la presenza in tutti questi bovini dei quattro aplogruppi T, T1, T2, T3, dei quali l’ultimo è predominante. Questa variabilità condivisa, non presente in altre razze autoctone italiane ed europee, tende a confermare la stessa origine genetica da popolazioni protostoriche bovine, di incipiente domesticazione, Anatoliche e Medio Orientali derivate da Uro locali.

39
La Maremmana e le sue caratteristiche
Razza rustica per eccellenza, con scheletro forte, incornatura poderosa, pelle spessa e robusta, grande sviluppo del treno anteriore, la Maremmana presenta performance produttive quantitativamente scadenti. Le modeste rese in carcassa hanno fatto sì che nel secolo scorso venissero avanzate proposte e fossero prese iniziative oggettivamente negative anche per la stessa sopravvivenza della razza.

47
La Maremmana e le sue caratteristiche
Dopo la bonifica della Maremma tosco-laziale molti volevano sostituire la rustica Maremmana con la più produttiva Chianina. La stessa stagionalità dei parti, risultato di una stagionalità nella monta naturale, era ed è visto come fattore limitante le possibilità di sviluppo della razza. Ma grazie alla caparbietà dei ricercatori della UNIFI ed agli allevatori, la razza fu salvata dall’estinzione e oggi, pur in presenza di una numerosità effettiva relativamente ridotta, è ancora presente al suo interno un livello soddisfacente di eterozigosi, premessa per una efficace opera di selezione.

48
La Maremmana e le sue caratteristiche
Tuttavia l’idea di una sostituzione parziale, attraverso la pratica dell’incrocio con la Chianina delle vacche maremmane continuò, tanto che fino agli anni cinquanta del secolo scorso si parlava di “razza” Chianino-Maremmana, meticcio relativamente stabilizzato. La pratica dell’incrocio, continuò, a partire dagli anni sessanta, con le francesi Charolaise e Limousine, per affievolirsi o fermarsi (in Toscana ma non nel Lazio) solo a partire dagli anni ottanta, con la presa di coscienza dell’importanza della conservazione e della valorizzazione in purezza dei genotipi autoctoni.
 solo a partire dagli anni ottanta, con la presa di coscienza dell’importanza della conservazione e della valorizzazione in purezza dei genotipi autoctoni.")
49
La Maremmana e le sue caratteristiche
D’altra parte, a fronte dell’oggettiva limitatezza delle produzioni dal punto di vista quantitativo, studi sulla qualità delle carni condotte UNIFI fin dagli anni novanta hanno dimostrato ottime caratteristiche qualitative, sia per quanto riguarda gli aspetti più propriamente sensoriali (sapore, tenerezza, succulenza), che per quelli dietetici (povertà in acidi grassi saturi, ricchezza in acidi grassi insaturi e particolarmente polinsaturi omega-6 e omega-3) Ne risultano valori di assoluta eccellenza degli indici che descrivono la capacità di esercitare un effetto protettivo nei confronti delle malattie cardiovascolari e che, in particolare nei vitelloni leggeri di mesi di età, non temono confronti con quelli di razze da carne “nobili”, Chianina compresa.
, che per quelli dietetici (povertà in acidi grassi saturi, ricchezza in acidi grassi insaturi e particolarmente polinsaturi omega-6 e omega-3) Ne risultano valori di assoluta eccellenza degli indici che descrivono la capacità di esercitare un effetto protettivo nei confronti delle malattie cardiovascolari e che, in particolare nei vitelloni leggeri di mesi di età, non temono confronti con quelli di razze da carne nobili , Chianina compresa.")
50
La Maremmana e le sue caratteristiche
Queste caratteristiche pregevoli si perdono, in gran parte, negli incroci e, oltre certe età, anche nei vitelloni in purezza. Ne deriva la necessità di trovare soluzioni innovative nei processi produttivi in grado di superare gli ostacoli che, a fronte della bontà qualitativa del prodotto, si oppongono a un auspicato sviluppo e successo di questa antica, nobile razza.

51
La Maremmana e le sue caratteristiche
18 mesi 15 mesi Acidi grassi saturi 43,3 42,9 Acidi grassi monoinsaturi 32,3 30,5 Acidi grassi polinsaturi ω-6 20,9 22,1 Acidi grassi polinsaturi ω-3 3,6 4,0 MONOINSATURI/SATURI 0,75 0,71 POLINSATURI/SATURI 0,57 0,62 Indice di aterogenesi (optimum < 1,0) 0,51 0,49 Indice di trombogenesi (optimum < 2,0) 1,09 0,98
 0,51. 0,49. Indice di trombogenesi. (optimum < 2,0) 1,09. 0,98.")
53
Grazie per l’attenzione!

Presentazioni simili