Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
emoglobina : trasporto dell’Ossigeno.

2
La transizione dalla vita anaerobica alla vita aerobica rese disponibile una grande riserva di energia. In presenza di O2 è possibile estrarre dal glucoso una quantità di energia 18 volte maggiore, ed è possibile usare come fonte di energia anche lipidi e protidi. I vertebrati hanno selezionato dei meccanismi per rifornire in modo adeguato tutte le cellule di O2 : -- Un sistema circolatorio, indispensabile per organismi di dimensioni maggiori di un mm3. -- Molecole trasportatrici di ossigeno che permettono di aumentare la solubilità nell’acqua del gas (1 l di H2O solubilizza 5 ml di O2 , 1 l di sangue 250 ml) e di ridurre la produzione dei radicali liberi dell’ossigeno,molto dannosi per tutte le molecole biologiche.
 e di ridurre la produzione dei radicali liberi dell’ossigeno,molto dannosi per tutte le molecole biologiche.")
3
CROMOPROTEINE - Emoglobina - mioglobina - citocromi - enzimi eminici

4
MIOGLOBINA ED EMOGLOBINA SONO PROTEINE CONIUGATE COSTITUITE DA UNA PARTE PROTEICA (GLOBINA) E DA UN GRUPPO PROSTETICO (FERROPROTOEME) CHE E’ ALLOGGIATO IN UNA TASCA DELLA GLOBINA E CHE LEGA REVERSIBILMENTE L’OSSIGENO
 E DA UN GRUPPO PROSTETICO (FERROPROTOEME) CHE E’ ALLOGGIATO IN UNA TASCA DELLA GLOBINA E CHE LEGA REVERSIBILMENTE L’OSSIGENO")
5
LA MIOGLOBINA E’ UNA PROTEINA MONOMERICA COSTITUITA DA UNA SINGOLA CATENA POLIPEPTIDICA LEGATA AD UN SINGOLO GRUPPO EME

6
L’EMOGLOBINA E’ UNA PROTEINA TETRAMERICA COSTITUITA DA QUATTRO CATENE POLIPEPTIDICHE CIASCUNA LEGATA AD UN GRUPPO EME

7
LA MIOGLOBINA E’ COSTITUITA DA UNA CATENA DI 153 AMMINOACIDI (STRUTTURA PRIMARIA) CHE SI SVILUPPA NELLO SPAZIO FORMANDO OTTO ELICHE (STRUTTURA SUPERSECONDARIA) ORGANIZZATE IN UN UNICO DOMINIO (STRUTTURA TERZIARIA)
 CHE SI SVILUPPA NELLO SPAZIO FORMANDO OTTO ELICHE (STRUTTURA SUPERSECONDARIA) ORGANIZZATE IN UN UNICO DOMINIO (STRUTTURA TERZIARIA)")
8
L’EMOGLOBINA HA UNA STRUTTURA QUATERNARIA
L’EMOGLOBINA HA UNA STRUTTURA QUATERNARIA. CONTIENE 4 CATENE GLOBINICHE: 2 (CON 141 AMMINOACIDICI) RIPIEGATE IN 7 REGIONI AD -ELICA) E 2 (CON 146 AMMINOACIDI) RIPIEGATE IN 8 REGIONI AD -ELICA). A CIASCUNA CATENA GLOBINICA E’ LEGATO UN FERROPROTOEME
 RIPIEGATE IN 7 REGIONI AD -ELICA) E 2 (CON 146 AMMINOACIDI) RIPIEGATE IN 8 REGIONI AD -ELICA). A CIASCUNA CATENA GLOBINICA E’ LEGATO UN FERROPROTOEME.")
9
IL GRUPPO EME ORIGINA DALLA PORFINA

10
IL FERROPROTOEME DERIVA DALLA PROTOPORFIRINA IX

11
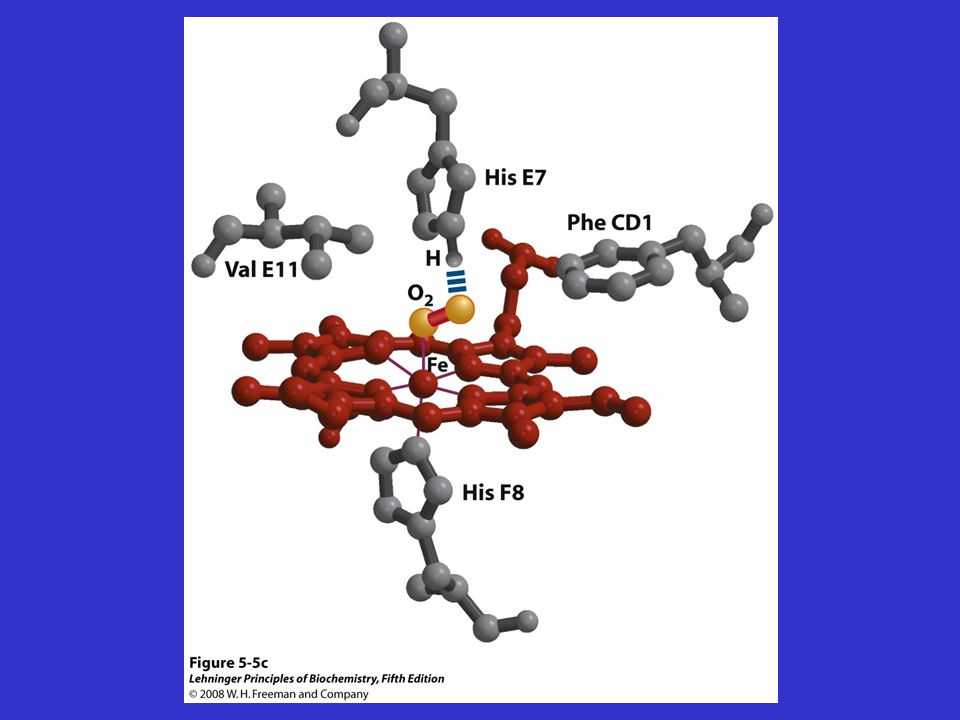
L’OSSIGENO SI LEGA REVERSIBILMENTE AL FERRO DELL’EME
Il Fe (allo stato ferroso) forma sei legami di coordinazione: 4 con gli N dell’anello porfirinico Il 5° con l’His F8 Il 6° con l’O2
 forma sei legami di coordinazione: 4 con gli N dell’anello porfirinico. Il 5° con l’His F8. Il 6° con l’O2.")
13
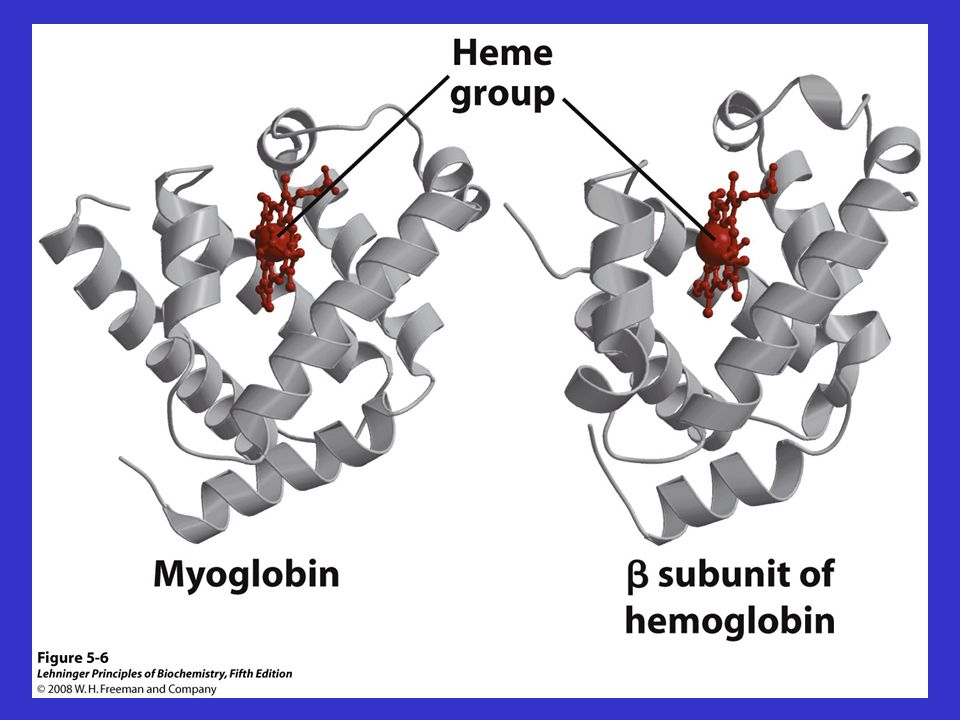
COMPLESSO DELL’EME NELLA MIOGLOBINA

14
STRUTTURA DELLA MIOGLOBINA
Le strutture della mioglobina, delle catene e di tutte le catene delle Hb di tutti i vertebrati sono molto simili nella struttura terziaria, anche se le strutture primarie sono differenti: LA CONFORMAZIONE E’ RESPONSABILE DELLA FUNZIONE

17
RUOLO DELLA GLOBINA Crea un ambiente idrofobico che impedisce l’ossidazione del Fe++ a Fe+++; His E7 (distale) diminuisce l’affinità del CO da volte maggiore dell’O2 a solo 210 volte
 diminuisce l’affinità del CO da volte maggiore dell’O2 a solo 210 volte.")
18
L’Hb è un tetramero
Deossi Hb L’Hb è un tetramero Ossi Hb

19
Durante la vita si susseguono diverse emoglobine
Emoglobine embrionali Hb Emoglobina fetale HbF Emoglobine dell’adulto HbA HbA2dell’emoglobina totale)
")
20
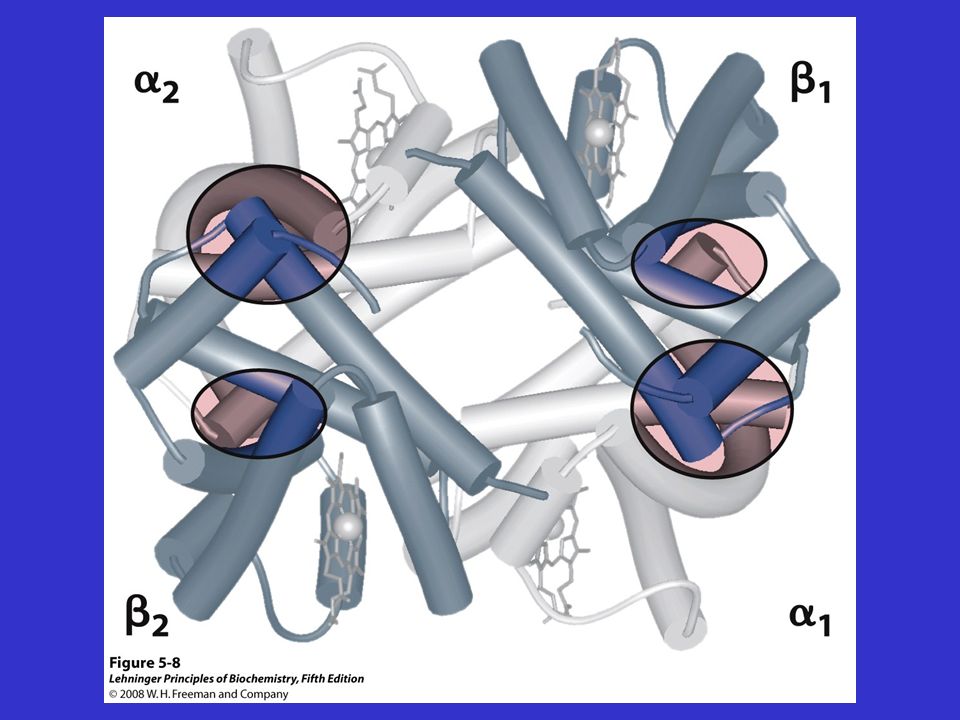
L’Hb è un tetramero in cui ogni subunità ha una struttura tutta ad -elica.
I contatti tra le subunità - sono di due tipi: -- Le associazioni e sono contatti estesi ed importanti per l’impacchettamento delle subunità (non si modificano quando l’Hb lega l’O2). -- Le regioni di contatto e sono chiamate regioni di scorrimento
. -- Le regioni di contatto e sono chiamate regioni di scorrimento.")
22
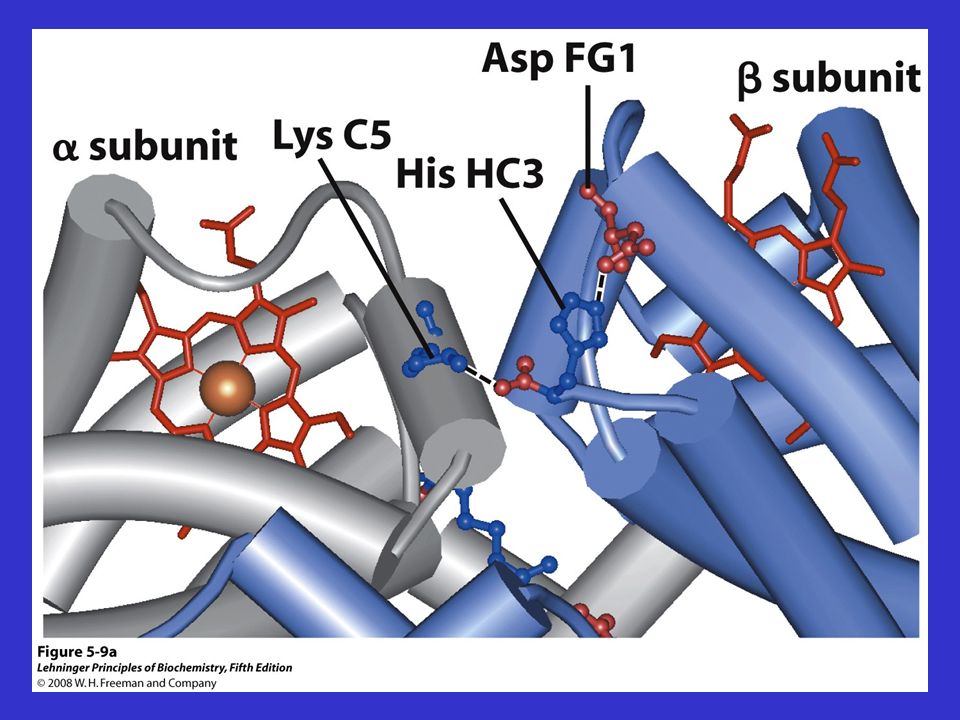
Ponti salini tra subunità differenti della Hb
Ponti salini tra subunità differenti della Hb. Sono interazioni elettrostatiche, non covalenti, che vengono rotte in seguito all’ossigenazione.

25
Curve di saturazione della Mb e della Hb
Questa diapositiva riporta le curve di ossigenazione di Hb e Mb NOTE la fig riporta un errore nella parte finale del grafico (zona asindotica per Mb ed HB che devono coincidere) La curva di ossigenasione della Hb va osservata in rapporto alla funzione che tale proteina deve svolgere. Questa cromoproteina infatti si sposta continuamente, all’interno degli eritrociti, dai polmoni dove a livello dei capillari arteriosi si è ossigenata e presenta una pressione parziale di O2 (pO2) prossima 110 a Torr, ai distretti tissutali dove in corrispondenza dei capillari venosi dovrà rilasciare l’O2. Tale curva pertanto va considera da ds verso sn, cioè dalla porzione asindotica verso sn. In grafico è riportata la pO2 contro la % di saturazione: pO2: esprime la [O2]; % saturazione: esprime la quantità di O2 legato alla Hb (HbO2). In condizioni di RIPOSO la Hb giunge ai capillari venosi dove rilascia solo il 25-30% dell’O2 legato. In tali condizioni la pO2 è di 40 TORR. Contestualmente però la Hb lega la Co2 per poi ritornare ai polmoni per riossigenarsi. Pertanto questa proteina in condizioni di riposo fa continuamente questo percorso dai capillari arteriosi dei polmoni dove si ossigena ai capillari venosi periferici dove rilascia l’ossigeno e ritorna indietro. Ma una proteina che fa questo percorso rilasciando solo il % dell’O2 legato funziona bene? Certamente sì perché in condizioni di attività quando si incrementa la concentrazione di H+ e il pH si abbassa questa curva è fortemente spostata verso ds e la Hb rilascia una quantità di O2 nettamente superiore. In condizioni di ATTIVITA’(per es. contrazione muscolare dopo uno sforzo fisico) si raggiunge una pO2 pari a 20 Torr, ovvero la Hb cede circa il 66% dell’O2 legato. E’ importante notare che in tali condizioni il muscolo produce metaboliti acidi, l’incremento della [H+] determina pertanto una riduzione del pH, e questo effetto (noto come EFFETTO BOHR) sposta verso ds la curva di ossigenazione della Hb incrementandone la sigmoidicità e riducendo l’affinità della Hb per l’O2 che è in tal modo rilasciato ai tessuti. la p50 è riferita a quella pO2 a cui il 50% della Hb è satura al 50%. Questo parametro è comparare l'affinità della Hb e della Mb per l'O2. la Mb è circa 26 volte più affine della Hb per l'O2. Questo consente alla Mb che deve trasportare l'O2 all'interno del muscolo di prelevarlo dalla Hb. p50 Hb = 26 torr p50 Mb = 2,8 torr pO2 = pressione parziale di O2 YO2 = saturazione frazionale 25
 La curva di ossigenasione della Hb va osservata in rapporto alla funzione che tale proteina deve svolgere. Questa cromoproteina infatti si sposta continuamente, all’interno degli eritrociti, dai polmoni dove a livello dei capillari arteriosi si è ossigenata e presenta una pressione parziale di O2 (pO2) prossima 110 a Torr, ai distretti tissutali dove in corrispondenza dei capillari venosi dovrà rilasciare l’O2. Tale curva pertanto va considera da ds verso sn, cioè dalla porzione asindotica verso sn. In grafico è riportata la pO2 contro la % di saturazione: pO2: esprime la [O2]; % saturazione: esprime la quantità di O2 legato alla Hb (HbO2). In condizioni di RIPOSO la Hb giunge ai capillari venosi dove rilascia solo il 25-30% dell’O2 legato. In tali condizioni la pO2 è di 40 TORR. Contestualmente però la Hb lega la Co2 per poi ritornare ai polmoni per riossigenarsi. Pertanto questa proteina in condizioni di riposo fa continuamente questo percorso dai capillari arteriosi dei polmoni dove si ossigena ai capillari venosi periferici dove rilascia l’ossigeno e ritorna indietro. Ma una proteina che fa questo percorso rilasciando solo il % dell’O2 legato funziona bene Certamente sì perché in condizioni di attività quando si incrementa la concentrazione di H+ e il pH si abbassa questa curva è fortemente spostata verso ds e la Hb rilascia una quantità di O2 nettamente superiore. In condizioni di ATTIVITA’(per es. contrazione muscolare dopo uno sforzo fisico) si raggiunge una pO2 pari a 20 Torr, ovvero la Hb cede circa il 66% dell’O2 legato. E’ importante notare che in tali condizioni il muscolo produce metaboliti acidi, l’incremento della [H+] determina pertanto una riduzione del pH, e questo effetto (noto come EFFETTO BOHR) sposta verso ds la curva di ossigenazione della Hb incrementandone la sigmoidicità e riducendo l’affinità della Hb per l’O2 che è in tal modo rilasciato ai tessuti. la p50 è riferita a quella pO2 a cui il 50% della Hb è satura al 50%. Questo parametro è comparare l affinità della Hb e della Mb per l O2. la Mb è circa 26 volte più affine della Hb per l O2. Questo consente alla Mb che deve trasportare l O2 all interno del muscolo di prelevarlo dalla Hb. p50 Hb = 26 torr. p50 Mb = 2,8 torr. pO2 = pressione parziale di O2. YO2 = saturazione frazionale. 25.")
26
EFFETTO COOPERATIVO Il legame dell’effettore alle subunità non avviene in maniera indipendente perché l’affinità dei singoli protomeri verso lo stesso effettore può essere diversa (aumentata o diminuita) in relazione allo stato della conformazione quaternaria dell’enzima. L’effetto cooperativo deriva dalle interazioni tra protomeri.
 in relazione allo stato della conformazione quaternaria dell’enzima. L’effetto cooperativo deriva dalle interazioni tra protomeri.")
27
EFFETTO COOPERATIVO POSITIVO
L’affinità dell’oligomero verso l’effettore aumenta quando una o più subunità dell’oligomero stesso hanno già legato l’effettore. EFFETTO COOPERATIVO NEGATIVO L’affinità dell’oligomero verso l’effettore diminuisce quando una o più subunità dell’oligomero stesso hanno già legato l’effettore.

28
La maggior parte degli enzimi allosterici sono regolati dallo stesso substrato con effetto omotropico positivo, cooperativo positivo.

29
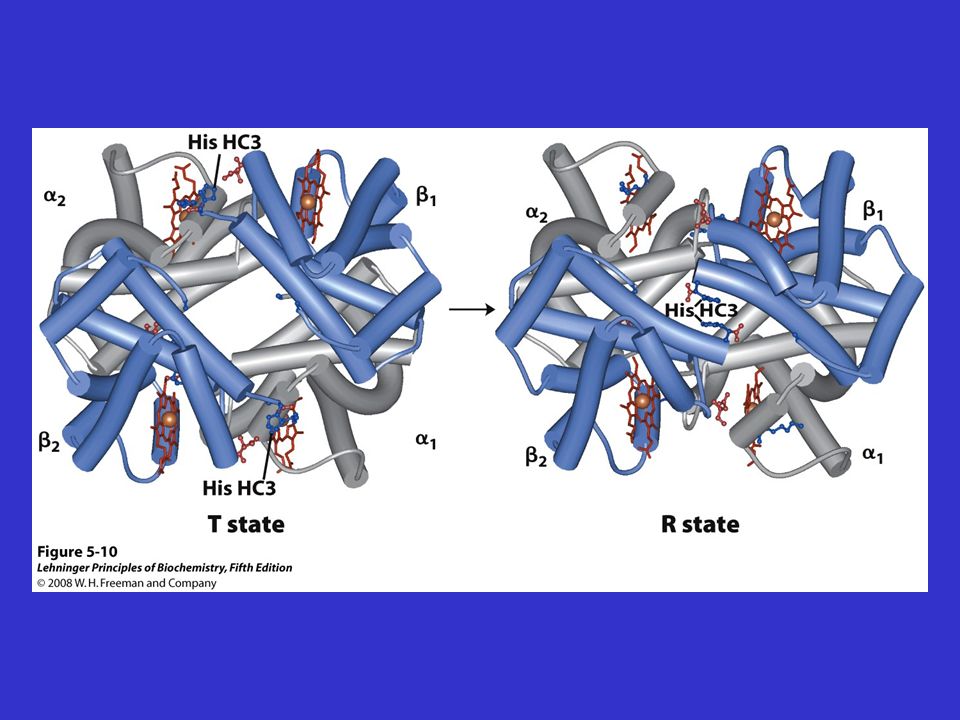
Modifiche conformazionali dell’Hb conseguenti all’ossigenazione
Blu forma T deossiHb Rossa forma R ossiHb

31
Movimenti dell’interfaccia durante la transizione T R

32
MODELLO SIMMETRICO DI MONOD

33
MODELLO SEQUENZIALE DI KOSHLAND

34
EFFETTORI ALLOSTERICI NEGATIVI
-pH -CO2 -2,3 bisfosfoglicerato

35
EFFETTO BOHR L’affinità dell’Hb per l’O2 aumenta all’aumentare del pH.
Un incremento della concentrazione idrogenionica (H+) corrispondente ad una riduzione del pH, comporta uno spostamento verso ds della curva di ossigenazione della Hb. determina pertanto una riduzione del pH, e questo effetto (noto come EFFETTO BOHR) che sposta verso ds la curva di ossigenazione della Hb incrementandone la sigmoidicità e riduce l’affinità della Hb per l’O2 che è in tal modo rilasciato ai tessuti. L’affinità dell’Hb per l’O2 aumenta all’aumentare del pH. 35
 corrispondente ad una riduzione del pH, comporta uno spostamento verso ds della curva di ossigenazione della Hb. determina pertanto una riduzione del pH, e questo effetto (noto come EFFETTO BOHR) che sposta verso ds la curva di ossigenazione della Hb incrementandone la sigmoidicità e riduce l’affinità della Hb per l’O2 che è in tal modo rilasciato ai tessuti. L’affinità dell’Hb per l’O2 aumenta all’aumentare del pH. 35.")
36
TRASPORTO CO2

37
Ruolo dell’Hb nel trasporto della CO2

39
Effetto del BPG e della CO2 sulla curva di dissociazione dell’Hb
Il BPG è presente nel sangue alla stessa [Molare] della Hb. In assenza di BPG la p50 dell’Hb è 1 torr. In presenza di BPG sale a 26 torr. Quindi abbassa l’affinità dell’Hb di 26 volte.

40
L’EMOGLOBINA FETALE CONTIENE DUE CATENE E DUE CATENE
LE CATENE LEGANO IL 2,3-DIFOSFOGLICERATO MENO SALDAMENTE DELLE DUE CATENE

41
CINETICA DI OSSIGENAZIONE DELL’EMOGLOBINA FETALE

42
EMOGLOBINA S ANEMIA A CELLULE FALCIFORMI
L’EMOGLOBINA S DIFFERISCE DALL’EMOGLOBINA NORMALE PERCHE’ NELLE CATENE (IN POSIZIONE 6) AL POSTO DELL’ACIDO GLUTAMMICO E’ PRESENTE LA VALINA
 AL POSTO DELL’ACIDO GLUTAMMICO E’ PRESENTE LA VALINA.")
43
La polimerizzazione della HbS attraverso le interazioni tra le catene laterali idrofobiche delle Val in posizione 6 e le tasche idrofobiche nei ripiegamenti EF delle catene in molecole adiacenti di Hb Ossi HbA Deossi HbA Ossi HbS Deossi HbS La deossiHbS polimerizza formando dei filamenti

44
FORMAZIONE DI FIBRE DI DEOSSIEMOGLOBINA A CAUSA DELLA VALINA MUTATA

45
FORMAZIONE DI FIBRE DI DEOSSIEMOGLOBINA

46
ERITROCITI NORMALI

47
ERITROCITI A FALCE

Presentazioni simili



















>")

>")
>")