Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Farmacologia Molecolare
Mer 7/4 pomeriggio dalla Prof. Curti per discussione orario Tentativamente, inizio lezioni: Mer 14/4 ore 9-11

2
Canale ionico in 3D – KcsA

3
Famiglie di canali ionici
I canali ionici sono raggruppati in diverse famiglie Canali della stessa famiglia tipicamente hanno: Stessa topologia di membrana della subunità che costituisce il poro del canale Somiglianze significative nelle sequenze

4
Canali ionici voltaggio-dipendenti
Famiglia di canali a 6 domini transmembrana (TM) Questa è una delle 4 (pseudo)subunità H5 Il poro è delineato da 4 subunità o pseudosubunità Ciascuna contiene 6 segmenti TM (S1-S6) e una regione H5 (loop P) Vi appartengono: Canali voltaggio-dipendenti del Na+, Ca2+ e K+ Canali del K+ Ca2+-attivati Canali cationici attivati dall’iperpolarizzazione, ecc.
 Questa è una delle 4 (pseudo)subunità. H5. Il poro è delineato da 4 subunità o pseudosubunità. Ciascuna contiene 6 segmenti TM (S1-S6) e una regione H5 (loop P) Vi appartengono: Canali voltaggio-dipendenti del Na+, Ca2+ e K+ Canali del K+ Ca2+-attivati. Canali cationici attivati dall’iperpolarizzazione, ecc.")
5
X 4

6
Canali del K inward rectifier
Famiglia di canali a 2 domini transmembrana (TM) Questa è una delle 4 (subunità Il poro è delineato da 4 subunità (Kir) Ciascuna contiene 2 segmenti TM e un loop interno Permettono una corrente di K+ inward maggiore della outward
 Questa è una delle 4 (subunità. Il poro è delineato da 4 subunità (Kir) Ciascuna contiene 2 segmenti TM e un loop interno. Permettono una corrente di K+ inward maggiore della outward.")
7
Recettori Ionotropici per il glutammato
Sono costituiti da 4 subunità Ciascuna ha 3 segmenti TM e un loop interno

8
Recettori Nicotinici e Recettori Ionotropici correlati
(5HT3, GABAA, Glicina) Ciascuna subunità ha 4 segmenti TM Sono costituiti da 5 subunità
 Ciascuna subunità ha 4 segmenti TM. Sono costituiti da 5 subunità.")
9
Come fa il voltaggio ad aprire i canali?
Il gating dei canali ovvero Come fa il voltaggio ad aprire i canali? Il controllo dell’attività dei canali per mezzo del voltaggio è la chiave che sta alla base dell’eccitabilità neuronale e del signalling

10
Il segmento S4 corrisponde al sensore del voltaggio
I canali ionici voltaggio-dipendenti contengono sensori del voltaggio intrinseci Il segmento S4 corrisponde al sensore del voltaggio

11
Il movimento del Sensore del Voltaggio Produce una Corrente di Gating
+50 mV -100 mV Corrente Ionica 1000 pA

12
Il movimento del Sensore del Voltaggio Produce una Corrente di Gating
+50 mV -100 mV Corrente Ionica 1000 pA 10 pA Corr. di Gating 1 ms Soluzione senza K+

13
Il movimento del Sensore del Voltaggio Produce una Corrente di Gating
+50 mV -100 mV Corrente Ionica 1000 pA 10 pA Corr. di Gating 1 ms Soluzione senza K+ Carica totale di gating = cariche e- /canale Numero dei canali

14
Attiva la spirale Nel canale del K+ “Shaker”, ciascun segmento S4 contiene 7 residui basici regolarmente distribuiti ogni 3 aa. Canale chiuso: la 2a Arg di S4 è seppellita nel bilayer Canale aperto: la 2a Arg fuoriesce dal lato extracell., mentre il 4o e 5o residui basici si muovono dal citosol all’interno del bilayer

15
Nel canale del K+ “Shaker” la presenza di 4 segmenti S4 che fungono da sensori del voltaggio intrinseci spiega la “ripida” dipendenza dell’attivazione del canale dal voltaggio. Le 4 subunità identiche si muovono verso il lato extracellulare in due fasi distinte: Prima indipendentemente l’una dall’altra; Poi in maniera cooperativa, portando all’apertura.

16
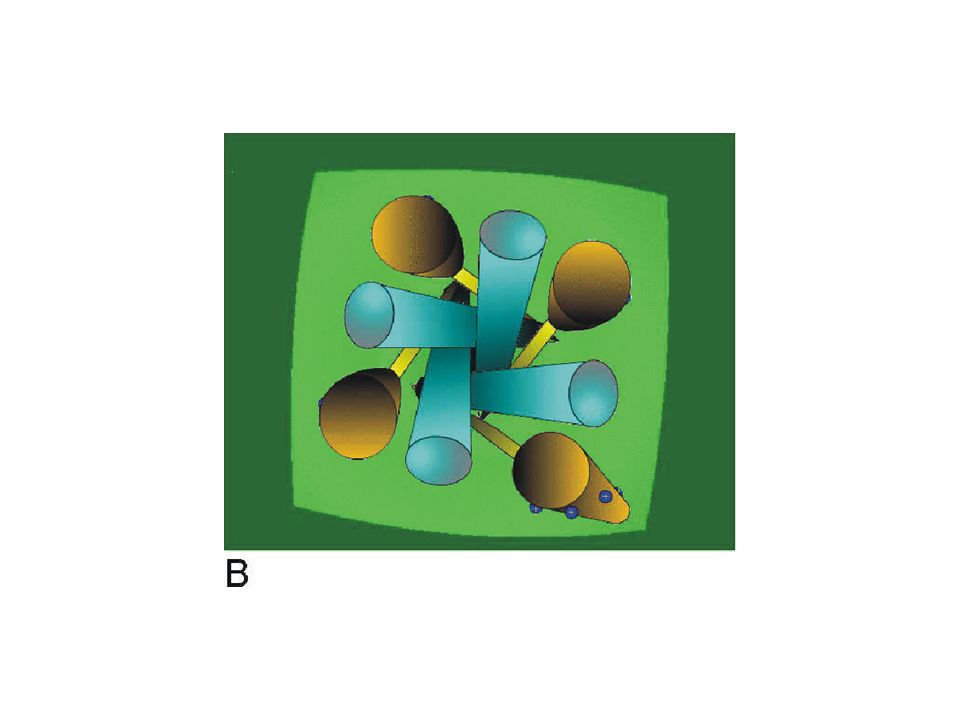
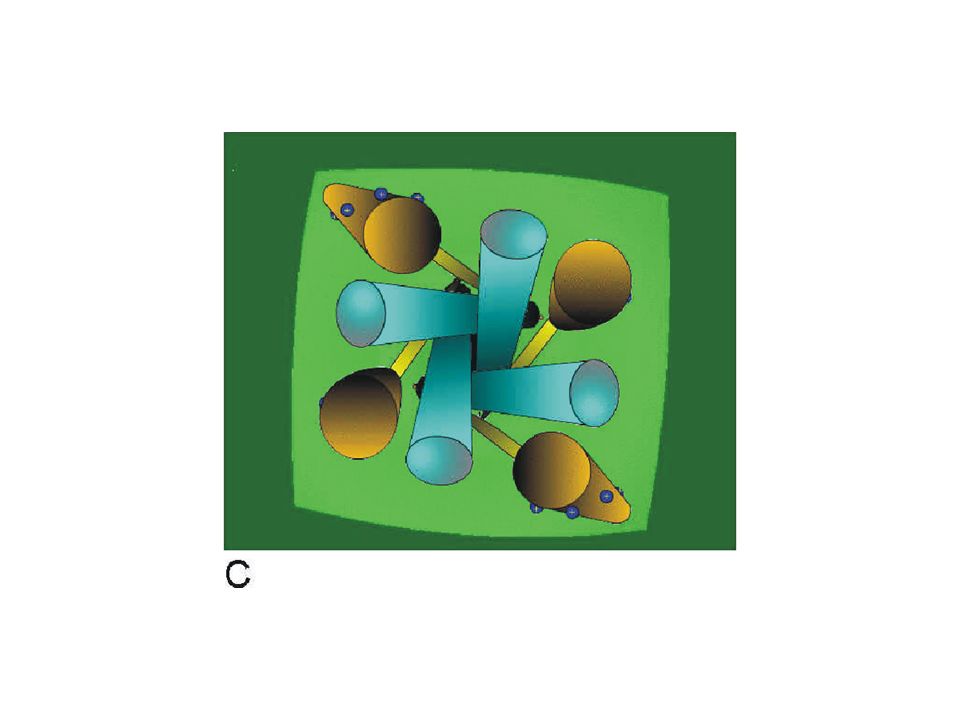
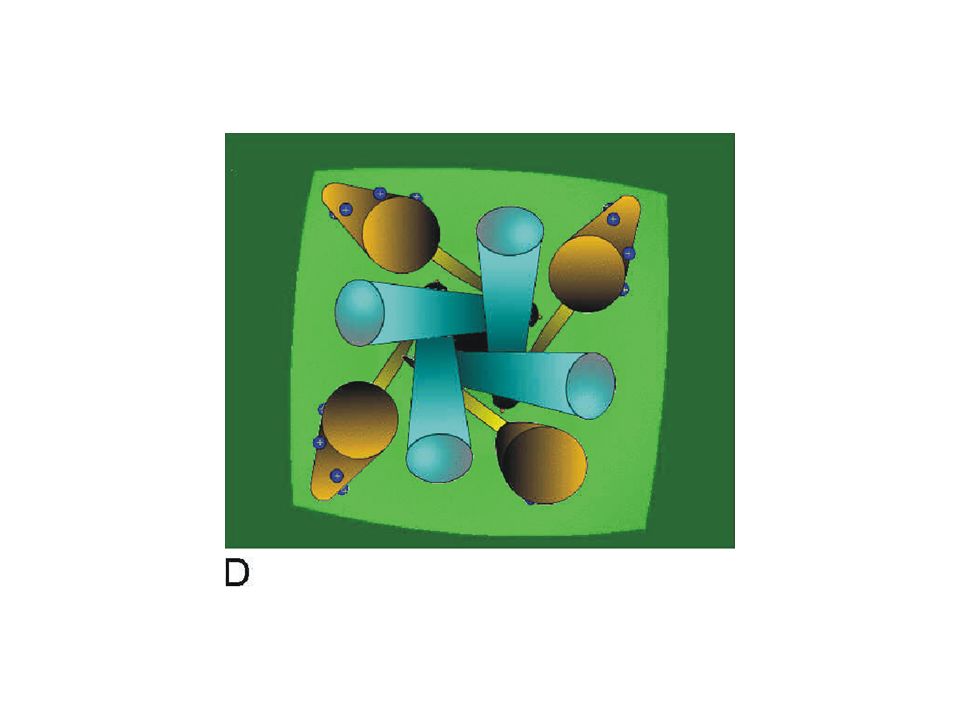
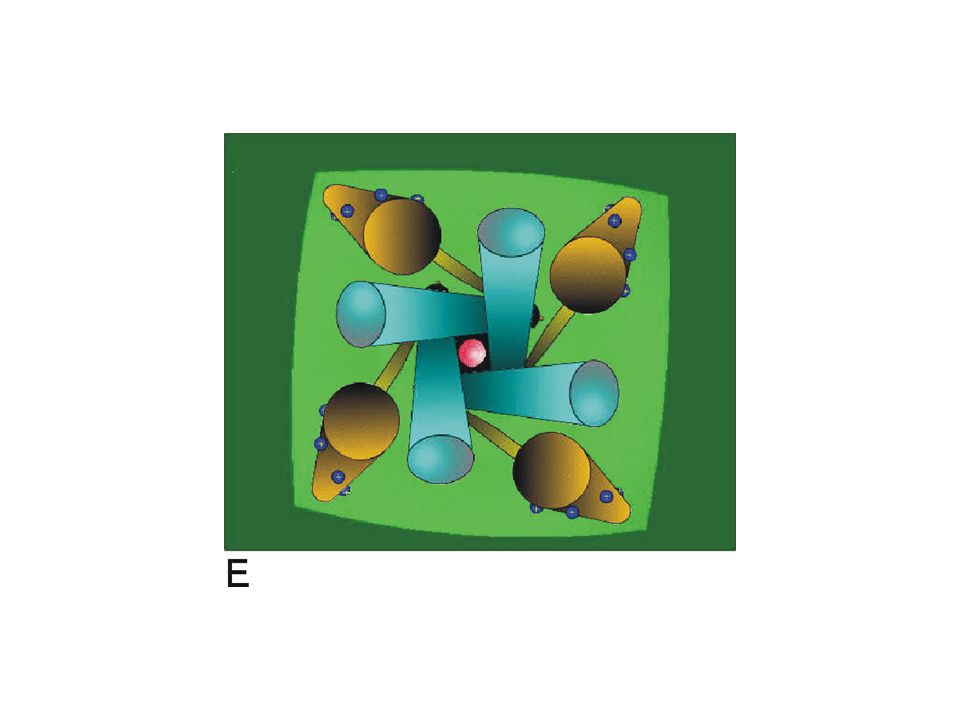
This simulation shows a simplified view of the channel from the outside of the cell membrane. The S6 segments of the pore (blue) represent the conduction pathway and the voltage sensors are represented by the S4 segments (brown). The link between the S4 and the pore is simply indicated as a bar (yellow) connecting the inner end of the S4 with the inner end of the S6 segment.
 represent the conduction pathway and the voltage sensors are represented by the S4 segments (brown). The link between the S4 and the pore is simply indicated as a bar (yellow) connecting the inner end of the S4 with the inner end of the S6 segment..")
21
Mutazioni in S4 Riducono il Movimento di Carica
1 2 3 4 5 6 7 IL R VI R LV R VF R IF K LS R HS K GL 4 R K 2 q/n R neutral AA - (e charges) -2 -4
")
22
Mutazioni in S4 Riducono il Movimento di Carica
1 2 3 4 5 6 7 IL R VI R LV R VF R IF K LS R HS K GL 4 R K 2 q/n R neutral AA - (e charges) -2 -4
")
23
MacKinnon_Nature01473.pdf MacKinnon_Nature01580.pdf MacKinnon_Nature01581.pdf

24
Struttura del Canale K+ Voltaggio-Dipendente KvAP da Aeropyrum pernix
La visione del tetramero del canale K+ è dal lato intracellulare. Ciascuna subunità è colorata separatamente e contiene 6 a-eliche (etichettate S1-S6 alle loro estremità N-terminali nella subunità blu). Le eliche S5-S6 formano il poro e il filtro di selettività per il K+. Le eliche S1-S4 formano i sensori del voltaggio. Gli ioni K+ sono colorati in verde.
. Le eliche S5-S6 formano il poro e il filtro di selettività per il K+. Le eliche S1-S4 formano i sensori del voltaggio. Gli ioni K+ sono colorati in verde.")
25
Struttura del Canale K+ Voltaggio-Dipendente KvAP da Aeropyrum pernix
Visione laterale. È indicato lo spessore della membrana (40 A)
")
26
Conformazioni del Sensore del Voltaggio
Vista Laterale: Le linee orizzontali demarcano approssimativamente i contorni della membrana. Si intravvedono 6 ioni K+; K1-K4 sono nel filtro di selettività. I residui comprendono il filtro di selettività e ne è mostrata l’ossatura in bianco. I gruppi C=O della catena principale di tali residui formano legami di coordinazione con gli ioni K+. Chiuso: Vr=-100 mV

27
Conformazioni del Sensore del Voltaggio
Vista Laterale: Le linee orizzontali demarcano approssimativamente i contorni della membrana. Si intravvedono 6 ioni K+; K1-K4 sono nel filtro di selettività. I residui comprendono il filtro di selettività e ne è mostrata l’ossatura in bianco. I gruppi C=O della catena principale di tali residui formano legami di coordinazione con gli ioni K+. Aperto: depolarizz.

28
Lo stato chiuso corrisponde al potenziale di riposo della cellula (~ -100 mV).
Lo stato aperto è favorito da una depolarizzazione della membrana. In questo modello, S2 è assunto rimanere inalterato durante lo switch; S3a e S3b subiscono dei riarrangiamenti sostanziali; si ipotizza che anche la connessione tra S4-S5 sia rimodellata.

29
Ingrandimento del Sensore del Voltaggio
Una Subunità, Chiusa: Delle eliche S2-S4 è mostrata l’ossatura. Le arginine che conferiscono la sensibilità al voltaggio, sono mostrate come bastoncini colorati. Sono mostrati anche l’ossatura del filtro di selettività e gli ioni K+. chiuso

30
Ingrandimento del Sensore del Voltaggio
aperto In aggiunta al movimento della “paletta sensore del voltaggio", notare nello stato aperto la formazione di un ponte salino (legame ionico) tra l’Asp 80 e l’Arg 133.
 tra l’Asp 80 e l’Arg 133.")
31
Accessibilità dei Residui
Nella conformazione chiusa, i residui evidenziati in magenta sono esposti allo spazio interno. Interno

32
Accessibilità dei Residui
Nella conformazione aperta, i residui evidenziati in rosso sono vicini allo spazio esterno (10 Å) o si estendono verso di esso. Esterno
 o si estendono verso di esso. Esterno.")
33
La struttura del canale del K+ KvAP è descritta in: A
La struttura del canale del K+ KvAP è descritta in: A. Jiang Y, Lee A, Chen J, Ruta V, Cadene M, Chait BT, & MacKinnon, R. (2003) "X-Ray Structure of a Voltage-Dependent K+ Channel". Nature 423: 33 I movimenti delle palette del voltaggio sono descritti in: B. Jiang Y, Ruta V, Chen J, Lee A, & MacKinnon, R. (2003) "The principle of gating charge movement in a voltage-dependent K+ Channel". Nature 423: 42 [1ORQ.pdb & 1ORS.pdb]
 X-Ray Structure of a Voltage-Dependent K+ Channel . Nature 423: 33. I movimenti delle palette del voltaggio sono descritti in: B. Jiang Y, Ruta V, Chen J, Lee A, & MacKinnon, R. (2003) The principle of gating charge movement in a voltage-dependent K+ Channel . Nature 423: 42. [1ORQ.pdb & 1ORS.pdb]")
34
La permeabilità agli ioni
Funzioni fisiologiche appropriate dei canali ionici dipendono dalla loro straordinaria selettività agli ioni Per raggiungere un’alta selettività senza trattenere lo ione troppo a lungo, i canali ionici hanno siti di legame multipli per lo ione permeante

35
Evidenze per la presenza di siti di legame:
Una dipendenza della permeabilità allo ione dalla composizione e dalla concentrazione dello ione implica la presenza di siti di legame nel canale Conduttanza Concentrazione del Na+ o K+ K+ Na+

36
Evidenze per l’esistenza di più di un sito di legame:
La presenza di più ioni permeanti nel poro del canale è visualizzabile da misure di cristallografia

37
La selettività ionica dei canali del Na+ e del Ca2+
I canali del Na+ e Ca2+ hanno pori più ampi di quelli dei canali del K+ e tuttavia favoriscono il passaggio di ioni più piccoli del K+ Come sono disegnati questi canali per essere selettivamente permeabili a Na+ e Ca2+ ?

38
In soluzione acquosa tutti gli ioni sono circondati da un alone di molecole d’acqua

39
Il poro dei canali del Na+ e Ca2+ è effettivamente più ampio di quello dei canali del K+, ma non sufficientemente grande da permettere la permeazione di ioni completamente idratati. Raggio anidro e idrato e spessore dell’alone idrico di solvatazione (idratazione) di alcuni ioni di interesse fisiologico Il processo di deidratazione e l’energia coinvolta costituiscono un fattore importante nel determinare la selettività. Probabilmente il poro di un canale approssima la conchiglia di idratazione del suo ione permeante, facilitando la perdita di molecole d’acqua durante il movimento dello ione lungo il canale.
 di alcuni ioni di interesse fisiologico. Il processo di deidratazione e l’energia coinvolta costituiscono un fattore importante nel determinare la selettività. Probabilmente il poro di un canale approssima la conchiglia di idratazione del suo ione permeante, facilitando la perdita di molecole d’acqua durante il movimento dello ione lungo il canale.")
40
La selettività di un canale K+ nei confronti del K+ rispetto al Na+
K+ in H2O Na+ in H2O K+ nel poro Na+ nel poro Gli ioni K+, idrati in soluzione, perdono le molecole di H2O quando passano attraverso il filtro di selettività e formano dei legami di coordinazione con quattro O di gruppi carbonilici C=O. Gli ioni Na+, essendo più piccoli, non possono coordinarsi perfettamente con questi O e quindi attraversano il canale solo raramente.

41
Quindi: Il canale K+ è selettivo per il K+ rispetto all’Na+ perchè gli ioni K+ formano legami più forti col filtro di selettività, compensando la perdita di energia dovuta alla deidratazione. Però, come si concilia il legame forte con il fatto che gli ioni K+ diffondono attraverso il canale quasi altrettanto rapidamente che nel solvente?

42
1. Nel filtro di selettività della proteina cristallizzata si possono vedere due ioni K+.

43
2. I due ioni K+ sono estremamente vicini tra di loro.
La repulsione tra questi due ioni carichi positivamente deve essere piuttosto forte, e tende a spingerne uno o entrambi fuori dal filtro.

44
canale-recettore per l’Ach
Dimensioni del Poro canale del potassio canale del sodio canale-recettore per l’Ach

Presentazioni simili


 (Ito) (IKr, IKs) (ICa-L) (INa).>")
















