Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Sviluppo e Differenziamento Delle Piante

2
Libri di testo: Taiz-Zeiger “Fisiologia vegetale” Piccin
VENERDI’ aula 8 MERCOLEDI’ aula 8 Dal 12 Marzo al 1 Giugno Prof. Mauro Marra stanza 349 Libri di testo: Taiz-Zeiger “Fisiologia vegetale” Piccin Buchanan “Biochimica e biologia molecolare delle piante” Zanichelli Articoli da riviste scientifiche

3
Programma del corso Sviluppo delle piante: basi molecolari della crescita e del differenziamento Embriogenesi, meristemi e sviluppo degli organi. Basi genetiche dello sviluppo del fiore. Controllo della fioritura. La Senescenza e la morte programmata nelle piante La maturazione dei frutti

4
DEFINIZIONI

5
Sviluppo: ha inizio negli eucarioti multicellulari con la fecondazione
Zigote: prolifera a formare un EMBRIONE multicellulare; si stabilisce una POLARITA’ e degli ASSI DI SIMMETRIA: quadro di riferimento per il differenziamento di gruppi di cellule in ORGANI e TESSUTI Come viene generata l’asimmetria? Come da una singola cellula iniziale, con poche divisioni si arriva ad una progenie di cellule con proprietà diverse le une dalle altre?

6
ASIMMETRIA: può essere generata in maniera diversa
da organismo a organismo Cellula uovo simmetrica: acquisizione asimmetria dipende dai cicli iniziali di divisione (mammiferi) Cellula uovo asimmetrica (componenti distribuiti asimmetricamente) (Drosophila, piante)
 Cellula uovo asimmetrica (componenti distribuiti. asimmetricamente) (Drosophila, piante)")
7
Da un punto di vista molecolare: diversi tipi cellulari = diversi pattern di
espressione genica Controllo attraverso i fattori di trascrizione Asimmetria iniziale tradotta in controllo della espressione di geni specifici, in modo che regioni specifiche della cellula uovo acquisiscano proprietà differenti Diversa localizzazione di fattori di trascrizione Controllo localizzato della loro attività CONTROLLO SPAZIALE E TEMPORALE DELL’ESPRESSIONE GENICA

8
STADIO INIZIALE seguito da uno stadio successivo in cui si determinano
le IDENTITA’ delle varie parti dell’EMBRIONE, dal quale si formeranno le diverse parti dell’organismo adulto Geni regolatori: GENI OMEOTICI (mutazioni causano l’assenza la duplicazione o lo scambio di organi) codificano per fattori di trascrizione AZIONE GERARCHICA CONTROLLO A CASCATA Un gene attivato in un certo stadio controlla l’espressione di un set genico dello stadio successivo
 codificano per fattori di trascrizione. AZIONE GERARCHICA. CONTROLLO A CASCATA. Un gene attivato in un certo stadio controlla l’espressione di un set genico. dello stadio successivo.")
9
Lo sviluppo di un organismo adulto da una cellula uovo fecondata
segue una via predeterminata in cui geni specifici sono attivati o repressi in momenti particolari Meccanismi cellulari possono essere diversi in specie diverse ma principi di base (drosophila) validi in generale Una cascata di eventi regolativi determina il pattern appropriato di espressione genica che porta all’organismo adulto
 validi in generale. Una cascata di eventi regolativi determina il pattern appropriato. di espressione genica che porta all’organismo adulto.")
10
PIANTE: Differenze rispetto agli animali nel pattern di sviluppo
Sviluppo meno determinato: le cellule vegetali mantengono un certo grado di TOTIPOTENZA. Tessuti differenziati possono essere indotti a sdifferenziarsi (calli) e di qui a rigenerare organi specifici e l’intera pianta (embriogenesi somatica)
 e di qui a rigenerare organi specifici e l’intera pianta. (embriogenesi somatica)")
11
Il movimento di cellule che è un aspetto importante
dell’embriogenesi animale, nelle piante è fortemente limitato dalla presenza della parete cellulare. Capacità di divisione cellulare ristretta alle ZONE MERISTEMATICHE;attività meristematica associata anche al differenziamento cellulare

12
Struttura delle piante

13
Il corpo vegetativo delle piante consiste di due parti:
Il sistema radicale Il sistema di parti aeree Sistema di parti aeree: fusto primario, rami Sistema radicale: radice primaria e radici secondarie e terziarie

14
Caratteristiche strutturali comuni a tutte le angiosperme, tuttavia
tra monocotiledoni e dicotiledoni alcune differenze anatomiche M: orchidee, gigli, palme, riso, mais D: rose fagioli, spinaci, girasole, querce

15
Ogni organo vegetale consiste di diversi tessuti e ogni tessuto
contiene molti tipi di cellule Gli organi vegetali consistono di tre diversi tessuti DERMICO VASCOLARE FONDAMENTALE In complesso questi tessuti contengono circa 40 diversi tipi cellulari Il corpo umano contiene diverse centinaia di tipi cellulari Piante organismi più semplici

16
Organizzazione dei tre sistemi di tessuti nel corpo della pianta

17
Tessuti dermici Epidermide: in piante giovani: singolo strato
di cellule con parete cellulare ispessita rivestita dalla cuticola (differenziamento in tricomi o cellule di guardia (stomi) nelle foglie e peli radicali nella radice) Periderma: in piante mature, comprende la corteccia; compare all’inizio dell’ispessimento e dopo la caduta dell’epidermide
 nelle foglie e peli radicali nella radice) Periderma: in piante mature, comprende la corteccia; compare all’inizio dell’ispessimento. e dopo la caduta dell’epidermide.")
18
Tessuti fondamentali Parenchima: cellule con parete sottile,
si trovano in tutti i tessuti. Foglie: fotosintesi (mesofillo) Fusto e radice: accumulo di amido e saccarosio Semi: amiloplasti, corpi proteici e corpi oleosi Floema: cellule compagne Collenchima: pareti cellulari più spesse, allungate, raggruppate in file verticali al di sotto dell’epidermide, con funzione di supporto meccanico. Sclerenchima: cellule morte con pareti ispessite e lignificate. Formano fibre che sostengono e proteggono il floema nei fusti
 Fusto e radice: accumulo di amido e saccarosio. Semi: amiloplasti, corpi proteici e corpi oleosi. Floema: cellule compagne. Collenchima: pareti cellulari più spesse, allungate, raggruppate in file verticali al di sotto dell’epidermide, con funzione di supporto meccanico. Sclerenchima: cellule morte con pareti ispessite. e lignificate. Formano fibre che sostengono e proteggono. il floema nei fusti.")
19
Tessuti vascolari Xilema: elementi dei vasi (tracheidi),
cellule allungate, morte con pareti ispessite e lignificate;Trasporto di acqua e soluti dalle radici alle foglie Floema: elementi dei tubi cribrosi cellule cribrose), cellule vitali prive di nucleo e Tonoplasto. Trasporto dei fotoassimilati nelle regioni sink della pianta.
, cellule vitali prive. di nucleo e Tonoplasto. Trasporto dei fotoassimilati. nelle regioni sink della pianta.")
20
Sviluppo e differenziamento
Piante: immobilità Maggiore capacità di adattamenti fisiologici Minore complessità anatomica rispetto agli animali Anatomia rigida: cellule incapaci di migrare (parete cellulare) Negli animali cellule migrano negli stadi precoci di sviluppo per formare gli organi Crescita mediante attività dei meristemi durante tutto il ciclo vitale (sviluppo vegetativo) Negli animali sviluppo stabilito essenzialmente durante l’embriogenesi
 Negli animali cellule migrano negli stadi precoci di sviluppo per formare. gli organi. Crescita mediante attività dei meristemi durante tutto il ciclo vitale. (sviluppo vegetativo) Negli animali sviluppo stabilito essenzialmente durante l’embriogenesi.")
21
Tutte le piante a seme passano attraverso tre stadi
di sviluppo dello sporofita EMBRIOGENESI SVILUPPO VEGETATIVO SVILUPPO RIPRODUTTIVO

22
Ciclo vitale di una pianta a fiore

23
Crescita indeterminata
La fase vegetativa dello sviluppo della pianta comincia con l’embriogenesi ma continua per tutta la vita della pianta. Le piante a differenza degli animali hanno tessuti specializzati, i meristemi, che generano continuamente nuove cellule ed organi Crescita indeterminata Ciò fornisce alle piante un modo semplice di risolvere il problema dell’invecchiamento: quando un organo è vecchio viene sostituito da uno nuovo abscissione fogliare

24
I Meristemi Meristema apicale del germoglio (SAM):
all’apice del germoglio una piccola massa di cellule a forma di cupola in continua divisione. Sono le progenitrici di tutte le cellule del germoglio. Le cellule immediatamente al di sotto sono anche meristematiche e la loro divisione contribuisce alla formazione degli organi in particolare il fusto. Dalla superfice del SAM emergono delle piccole protrusioni, i primordi, che evolveranno in piccole foglie. Inoltre si formano nuovi meristemi che daranno luogo alle gemme ascellari, quiescenti fino all’arrivo di uno stimolo ormonale Nell’insieme il meristema apicale (SAM) e i primordi formano la gemma apicale Stimoli ambientali, in primo luogo luce e temperatura regolano l’attività del SAM, attraverso la variazione dei livelli di alcuni ormoni Gli ormoni a loro volta attivano cascate geniche che controllano nel meristema la velocità di divisione cellulare, la dimensione del meristema, l’esatta posizione in cui si formano gli organi (primordio fogliare) e la velocità di crescita dell’organo
 e i primordi formano la gemma apicale. Stimoli ambientali, in primo luogo luce e temperatura regolano l’attività del SAM, attraverso la variazione. dei livelli di alcuni ormoni. Gli ormoni a loro volta attivano cascate geniche che controllano nel meristema la velocità di divisione cellulare, la dimensione del meristema, l’esatta posizione in cui si formano gli organi (primordio fogliare) e la velocità. di crescita dell’organo.")
25
Meristema apicale della radice (RAM) :
Il meristema apicale della radice è presente all’apice di ogni radice, appena al disotto della cuffia, la struttura che protegge il meristema quando la radice cresce nel suolo. La cuffia ha le proprie cellule meristematiche o iniziali che continuano a produrre cellule della cuffia, le quali poi cadono durante la crescita mantenendo costanti le dimensioni della cuffia. La radice stessa si origina da poche cellule iniziali (da tre a sei). Durante la crescita le radici laterali non si formano dal meristema apicale ma da un meristema secondario che si differenzia dal periciclo per effetto dell’auxina. Come nei fusti, il pattern di sviluppo è quello di continuo accrescimento della radice e di formazione di RAM.
. Durante la crescita. le radici laterali non si formano dal. meristema apicale ma da. un meristema secondario. che si differenzia dal periciclo. per effetto dell’auxina. Come nei fusti, il pattern di sviluppo. è quello di continuo. accrescimento della radice e. di formazione di RAM.")
26
SAM e RAM: cellule più piccole di quelle parenchimatiche ( volte). Hanno nuclei prominenti e citoplasma denso ricco di ribosomi e plastidi poco sviluppati. La loro funzione principale è quella di dividersi (ogni ore). Hanno parete cellulare sottile che viene riformata dopo la divisione. Alla periferia dei meristemi le cellule cominciano ad uscire dal programma di divisione e cominciano ad espandersi e poi a differenziarsi mediante l’espressione di migliaia di geni non espressi nelle cellule meristematiche. SAM ha la funzione di produrre la parte aerea del corpo vegetativo della pianta (foglie e fusto) ma per effetto di stimoli ambientali e/o ormonali il SAM può convertirsi nel meristema fiorale che produrrà i primordi fiorali e poi la struttura riproduttiva matura (fiore) con i vari organi ( sepali, petali, stami , carpelli)
. Hanno parete cellulare sottile che viene riformata. dopo la divisione. Alla periferia dei meristemi le cellule cominciano ad uscire dal. programma di divisione e cominciano ad espandersi e poi a differenziarsi mediante. l’espressione di migliaia di geni non espressi nelle cellule meristematiche. SAM. ha la funzione di produrre la parte aerea del corpo vegetativo della pianta (foglie e fusto) ma per effetto di stimoli ambientali e/o ormonali il SAM può convertirsi nel meristema. fiorale che produrrà i primordi fiorali e poi la struttura riproduttiva matura (fiore) con i vari organi ( sepali, petali, stami , carpelli)")
27
Gli stimoli ambientali e gli ormoni attivano programmi genetici
che regolano lo sviluppo delle piante

28
Specifici stimoli ambientali sono richiesti per far procedere una pianta da uno stadio di sviluppo al successivo Interruzione dormienza luce Deeziolatura Fioritura fotoperiodo vernalizzazione

29
Nel primo stadio di sviluppo, dalla cellula
uovo fecondata si origina il seme Nelle piante, a differenza degli animali, gli organi riproduttivi maschili e femminili si trovano sullo stesso individuo Celule specializzate nello stame e nel carpello vanno incontro a meiosi, producendo cellule spermatiche (stame) e cellule uovo (carpello) aploidi. Nell’ovulo si forma una singola cellula uovo Nel tubetto pollinico sono presenti due cellule spermatiche La fecondazione produce lo zigote diploide che si sviluppa nell’embrione, parte del seme. Un secondo evento di fertilizzazione produrrà il nucleo polare da cui si genera l’endosperma del seme. La formazione dell’embrione e del seme avviene nei tessuti (ovario) della pianta madre.
 e cellule uovo (carpello) aploidi. Nell’ovulo si forma una singola cellula uovo. Nel tubetto pollinico sono presenti due cellule. spermatiche. La fecondazione produce lo zigote diploide che si sviluppa nell’embrione, parte del seme. Un secondo evento di fertilizzazione produrrà il nucleo polare da cui si genera l’endosperma. del seme. La formazione dell’embrione e del seme avviene nei tessuti (ovario) della pianta madre.")
30
aspetto chiave: formazione dell’asse embrionale
EMBRIOGENESI: il processo in generale Dopo la fecondazione, lo zigote va incontro a numerose divisioni fino a formare l’embrione La crescita dell’embrione ha luogo dentro l’ovulo in un mezzo ricco di nutrienti: l’endosperma liquido (crescita eterotrofica) L’endosperma sintetizza i nutrienti da saccarosio e aminoacidi forniti dalla pianta madre. Mano a mano che si accresce, l’embrione acquisisce capacità biosintetiche ma rimane dipendente dal rifornimento di saccarosio e aminoacidi dalla pianta madre. aspetto chiave: formazione dell’asse embrionale SAM e RAM situati ai poli opposti dell’embrione In quale stadio avviene il differenziamento? studi sullo stadio a 16 cellule di Arabidopsis thaliana formazione del seme: Formato l’embrione, vengono sintetizzate le riserve di nutrienti (proteine, lipidi, amido) e si accumulano nelle cellule parenchimatiche di riserva dei cotiledoni o di altri tessuti. Nelle dicotiledoni: accumulo nei cotiledoni che diventano molto grandi Nei cereali e altre erbacee accumulo nell’endosperma Accumulo in organelli specializzati: amiloplasti (amido), corpi proteici (proteine), corpi oleosi (lipidi)
 L’endosperma sintetizza i nutrienti da saccarosio e aminoacidi forniti dalla pianta madre. Mano a mano che si accresce, l’embrione acquisisce capacità biosintetiche ma rimane. dipendente dal rifornimento di saccarosio e aminoacidi dalla pianta madre. aspetto chiave: formazione dell’asse embrionale. SAM e RAM situati ai poli opposti dell’embrione. In quale stadio avviene il differenziamento studi sullo stadio a 16 cellule di Arabidopsis thaliana. formazione del seme: Formato l’embrione, vengono sintetizzate le riserve di nutrienti (proteine, lipidi, amido) e si accumulano nelle cellule parenchimatiche di riserva dei cotiledoni o di altri tessuti. Nelle dicotiledoni: accumulo nei cotiledoni che diventano molto grandi. Nei cereali e altre erbacee accumulo nell’endosperma. Accumulo in organelli specializzati: amiloplasti (amido), corpi proteici (proteine), corpi oleosi (lipidi)")
31
Dallo zigote al seme maturo:
Attivazione di complessi programmi genetici: Divisione cellulare, formazione di tessuti ed organi dell’embrione Espansione degli organi di riserva e biosintesi dei composti di riserva Resistenza al dessiccamento (accumulo di proteine, zuccheri e oligosaccaridi idrofilici con funzione protettiva)
")
32
EMBRIOGENESI Dà inizio allo sviluppo della pianta
Comincia con l’unione di una cellula uovo e una cellula spermatica a formare lo zigote, ma in casi particolari anche cellule somatiche possono andare incontro a embriogenesi (embriogenesi somatica) Trasforma lo zigote a singola cellula in una piantina multicellulare, microscopica, embrionale. L’embrione completo ha la stessa organizzazione del corpo vegetale della pianta matura e la gran parte dei tessuti, anche se alcuni di questi sono presenti in forme più rudimentali. La fecondazione da luogo in realtà ad altri tre processi di sviluppo: formazione dell’endosperma, del seme e del frutto. Nelle angiosperme oltre alla formazione dello zigote c’è un secondo evento di fertilizzazione in cui un’altra cellula spermatica si unisce con due nuclei polari e da luogo all’endosperma triploide L’Embriogenesi ha luogo nel sacco embrionale all’interno dell’ovulo. Altre strutture associate all’ovulo danno luogo alle altre parti del seme. L’ embriogenesi e lo sviluppo dell’endosperma decorrono parallelamente allo sviluppo del seme l’embrione è parte del seme.L’endosperma è anche parte del seme ma in alcune specie può scomparire nel seme maturo.
 Trasforma lo zigote a singola cellula in una piantina multicellulare, microscopica, embrionale. L’embrione completo ha la stessa organizzazione del corpo vegetale della pianta matura e la gran parte. dei tessuti, anche se alcuni di questi sono presenti in forme più rudimentali. La fecondazione da luogo in realtà ad altri tre processi di sviluppo: formazione dell’endosperma, del seme e del frutto. Nelle angiosperme oltre alla formazione dello zigote c’è un secondo evento. di fertilizzazione in cui un’altra cellula spermatica si unisce con due nuclei polari e da luogo. all’endosperma triploide. L’Embriogenesi ha luogo nel sacco embrionale all’interno dell’ovulo. Altre strutture associate all’ovulo danno luogo alle altre parti del seme. L’ embriogenesi e lo sviluppo dell’endosperma decorrono parallelamente allo sviluppo del seme. l’embrione è parte del seme.L’endosperma è anche parte del seme ma in alcune specie può. scomparire nel seme maturo.")
33
Arabidopsis thaliana appartiene alla famiglia delle brassicacee Piccola, dal ciclo vitale breve, particolarmente adatta agli studi di genetica e biologia molecolare Genoma piccolo e completamente sequenziato Sforzo comune della ricerca per comprendere la funzione di ogni suo gene entro il 2010 Arabidopsis 107 bp Fritillaria 1011 bp

35
Pattern di sviluppo apicale-basale (assiale)
L’Embriogenesi stabilisce le caratteristiche essenziali della pianta adulta Nelle piante, diversamente dagli animali l’embriogenesi non genera direttamente i tessuti e gli organi dell’adulto Nelle angiosperme formazione di un corpo rudimentale: asse embrionale e due cotiledoni (dicotiledone) Tuttavia definite le caratteristiche dello sviluppo che si riscontrano nella pianta matura Pattern di sviluppo apicale-basale (assiale) Pattern di sviluppo radiale Inoltre l’embriogenesi determina la formazione dei meristemi primari, i quali dopo la germinazione daranno luogo agli organi e ai tessuti della pianta adulta.
 Tuttavia definite le caratteristiche dello sviluppo che si riscontrano nella pianta matura. Pattern di sviluppo apicale-basale (assiale) Pattern di sviluppo radiale. Inoltre l’embriogenesi determina la formazione dei meristemi primari, i quali dopo la germinazione daranno luogo agli organi e ai tessuti della pianta adulta.")
36
Sviluppo post embrionale di
arabidopsis

37
Pattern assiale Pattern radiale

38
Formazione del pattern assiale:
Le piante esibiscono una polarità assiale nella quale organi e tessuti sono organizzati In un ordine preciso lungo un asse lineare o polarizzato Meristema apicale Ipocotile Radice Meristema radicale Cuffia Viene stabilito durante l’embriogenesi Ogni segmento a sua volta esibisce una polarità con caratteristiche fisiologiche distinte tra parte basale e apicale (radici avventizie: si sviluppano dalla estremità basale dei fusti)
")
39
Ovulo di arabidopsis con il sacco embrionale
lo zigote è polarizzato

40
Caratteristiche cellulari dell’embriogenesi di arabidopsis

41
Formazione del pattern radiale:
Tessuti differenti sono organizzati secondo un pattern riconoscibile in un organo Fusti, radici: pattern radiale dall’esterno verso il centro Epidermide Cortex Cilindro vascolare (endodermide, periciclo, floema, xilema) Protoderma (epidermide) Meristema fondamentale (cortex, endodermide) Procambio (tessuto vascolare primario, cambio vascolare)
 Protoderma. (epidermide) Meristema fondamentale. (cortex, endodermide) Procambio. (tessuto vascolare primario, cambio vascolare)")
42
Sviluppo dell’embrione (Arabidopsis)
4 stadi morfologici Stadio globulare Stadio a cuore (cotiledoni, simmetria bilaterale) Stadio a siluro (espansione delle cellule, sviluppo dei cotiledoni Stadio di maturazione (perdita di acqua, tolleranza al dessiccamento 1 protoderma Cellula apicale Cellula basale cotiledone asse 2 3 4
 Stadio a siluro (espansione delle cellule, sviluppo dei cotiledoni. Stadio di maturazione (perdita di acqua, tolleranza al dessiccamento. 1. protoderma. Cellula apicale. Cellula basale. cotiledone. asse")
43
Il pattern embriogenico è diverso nelle monocoltiledoni
Pattern più variabile tuttavia evidenziabili analoghi stadi di sviluppo Stadio globulare Stadio di Coleoptile Stadio vegetativo giovanile Stadio di maturazione Embriogenesi in riso

44
La polarità assiale si genera già durante la prima divisione dello zigote
Lo zigote si espande e diventa polarizzato esso stesso prima di dare luogo alla prima divisione La parte apicale è con citoplasma denso La parte basale contiene il vacuolo centrale Prima divisione: asimmetrica perpendicolarmente all’asse principale dello zigote Si formano 2 cellule Cellula apicale: forma tutte le strutture dell’embrione; due divisioni verticali e una orizzontale danno luogo allo stadio globulare dell’embrione (8 cellule) Cellula basale: divisioni orizzontali; perpendicolari all’asse principale; si forma un filamento da 6 a 9 cellule (sospensore: extraembrionale). La prima di queste cellule formerà invece l’ipofisi che è parte dell’embrione (cuffia) Il sospensore connette l’embrione al sistema vascolare della pianta madre
 Cellula basale: divisioni orizzontali; perpendicolari all’asse principale; si forma un. filamento da 6 a 9 cellule (sospensore: extraembrionale). La prima di queste cellule. formerà invece l’ipofisi che è parte dell’embrione (cuffia) Il sospensore connette l’embrione al sistema vascolare della pianta madre.")
45
La polarità assiale è chiaramente distinguibile nello stadio a cuore
Tre regioni assiali: Regione apicale: forma i cotiledoni e il meristema apicale Regione mediana: forma l’ipocotile, la radice e la maggior parte del meristema radicale L’ipofisi: forma il resto del meristema radicale

46
l’ipofisi contribuisce alla formazione dell’embrione:
La cellula derivata dalla cellula basale più vicina all’embrione, l’ipofisi contribuisce alla formazione dell’embrione: forma la columella, cioè la parte centrale della cuffia radicale e il centro quiescente

47
Anche La polarità radiale si stabilisce durante l’embriogenesi ed
è distinguibile già nello stadio globulare: Tre tessuti Protoderma (epidermide) Meristema fondamentale (cortex, endodermide) Procambio (tessuti vascolari, periciclo)
 Meristema fondamentale (cortex, endodermide) Procambio (tessuti vascolari, periciclo)")
48
Formazione del pattern radiale

49
Meccanismi di signaling posizionale guidano l’embriogenesi
Negli animali Pattern riproducibile di divisioni cellulari programma di divisioni fisso per ogni cellula Nelle piante In piante diverse da Arabidopsis pattern di divisioni cellulari più variabili Probabilmente operante un sistema di signaling posizione dipendente Il destino differenziativo della cellula determinato dalla sua posizione nell’embrione

51
MORFOGENI il principale morfogeno durante l’embriogenesi
Negli animali segnali chimici svolgono un ruolo determinante durante l’embriogenesi MORFOGENI Informazione posizionale mediante gradienti di concentrazione Nelle piante ci sono evidenze che sia L’AUXINA il principale morfogeno durante l’embriogenesi

52
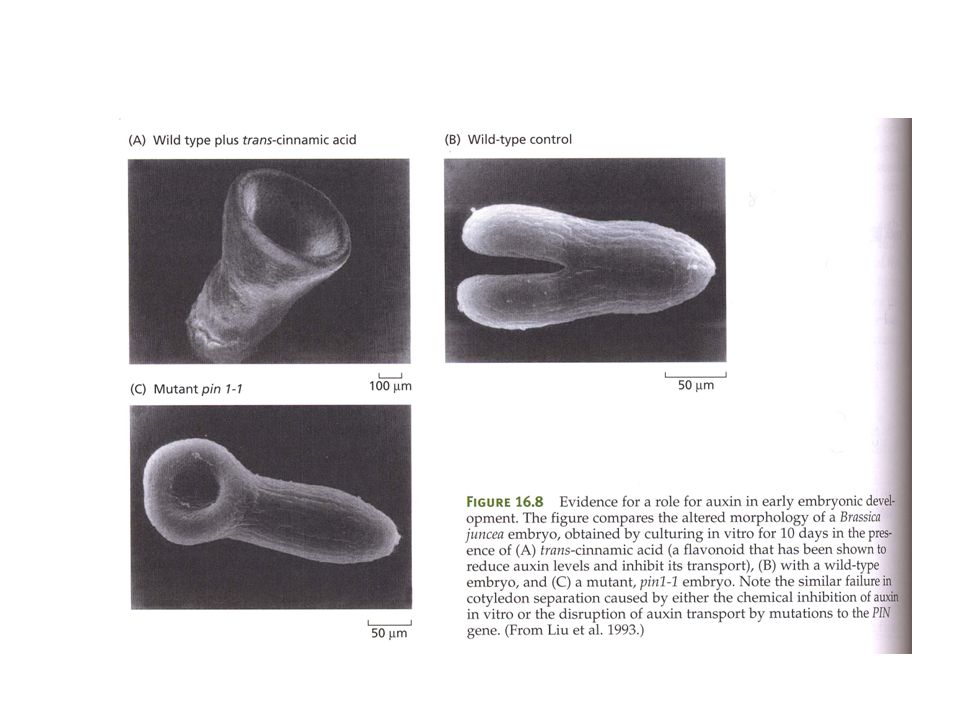
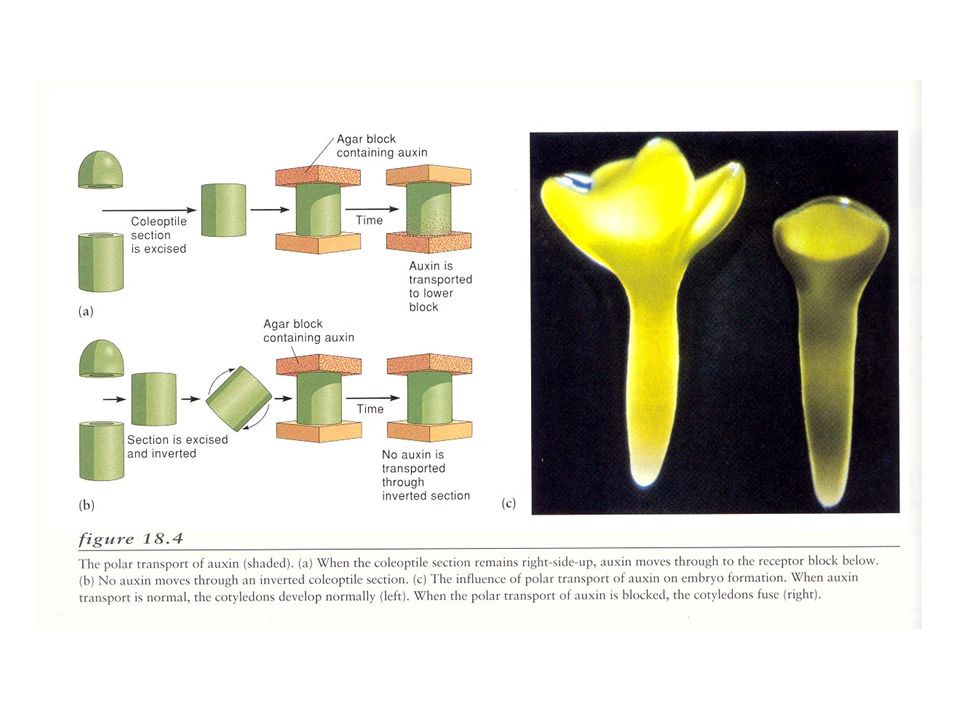
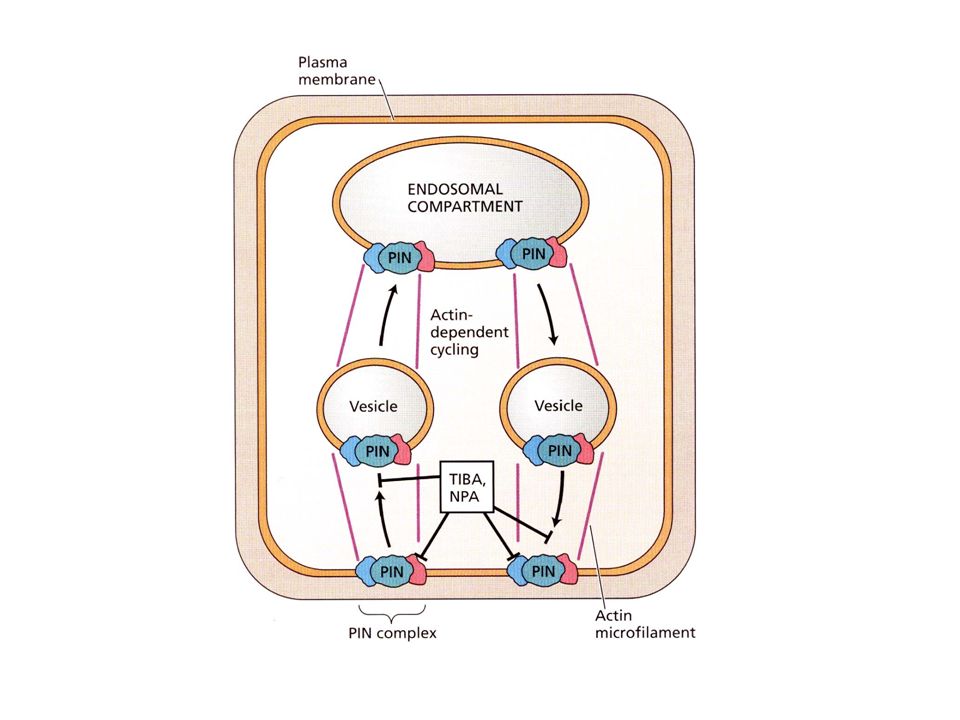
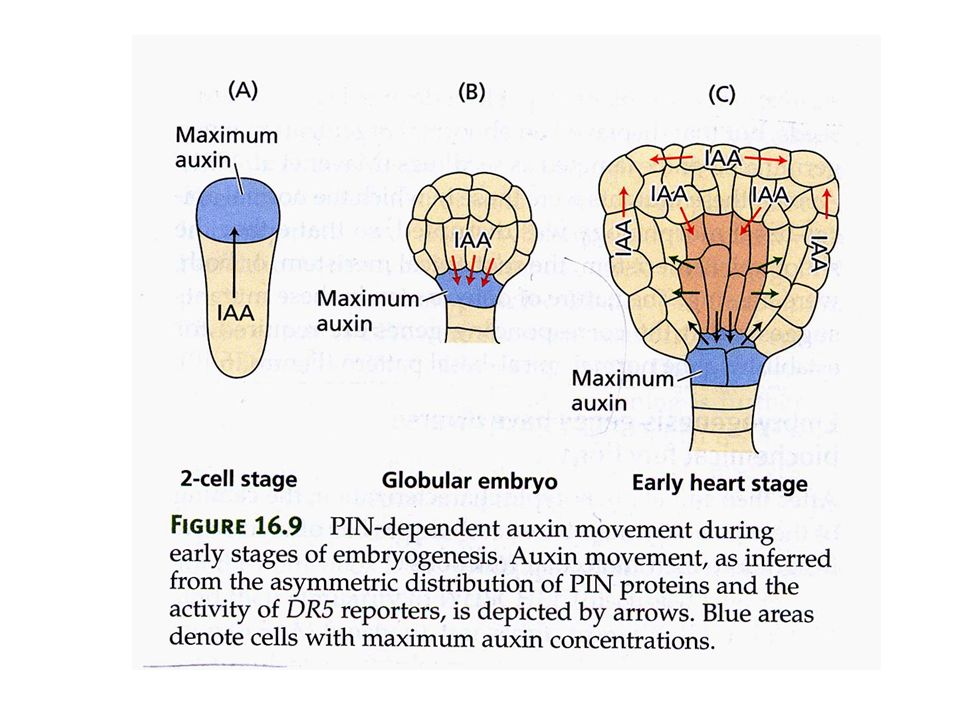
AUXINA COME MORFOGENO: EVIDENZE
L’auxina può indurre la formazione di embrioni da cellule somatiche L’auxina nell’embrione in sviluppo mostra gradienti di concentrazione (gene reporter) Proteine di trasporto dell’auxina nell’embrione mostrano una localizzazione asimmetrica
 Proteine di trasporto dell’auxina nell’embrione mostrano una localizzazione. asimmetrica.")
53
pianta Arabidopsis trasformata con il gene DR5::GUS
DR5 = promotore gene GH3

58
GENI CHE CONTROLLANO LA
FORMAZIONE DEL PATTERN APICALE-BASALE

59
L’analisi genetica del “pattern formation” in embrioni di Arabidopsis
ha evidenziato che: L’Embriogenesi richiede l’espressione di geni specifici L’analisi di mutanti di Arabidopsis che non riescono a stabilire una polarità assiale o si sviluppano in maniera anomala durante la germinazione ha portato alla identificazione di geni coinvolti nella formazione del pattern assiale pattern radiale meristemi radicale e apicale

61
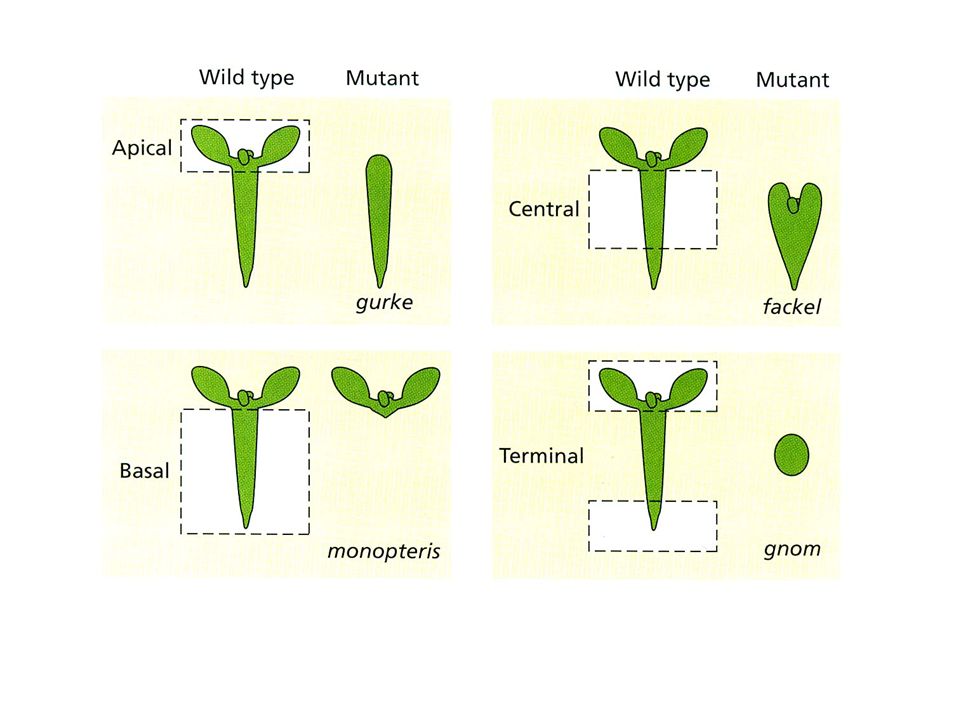
GURKE: deriva il nome dalla forma a cetriolo del mutante che è privo
dei cotiledoni e del meristema apicale. Il gene codifica per una acetil-CoA carbossilasi FACKEL: il mutante manca di ipocotile. Il gene codifica per una sterolo C14 reduttasi MONOPTEROS (MP): Necessario per la formazione degli elementi basali ipocotile e radice. Il gene codifica per un fattore di risposta all’auxina (ARF) GNOM (GN): richiesto per la formazione degli elementi terminali apicale e basale Codifica per un fattore di scambio di guanin nucleotidi (GEF):
: Necessario per la formazione degli elementi. basali ipocotile e radice. Il gene codifica per un fattore di risposta. all’auxina (ARF) GNOM (GN): richiesto per la formazione degli elementi terminali apicale e basale. Codifica per un fattore di scambio di guanin nucleotidi (GEF):")
Presentazioni simili