Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Insufficienza cardiaca

2
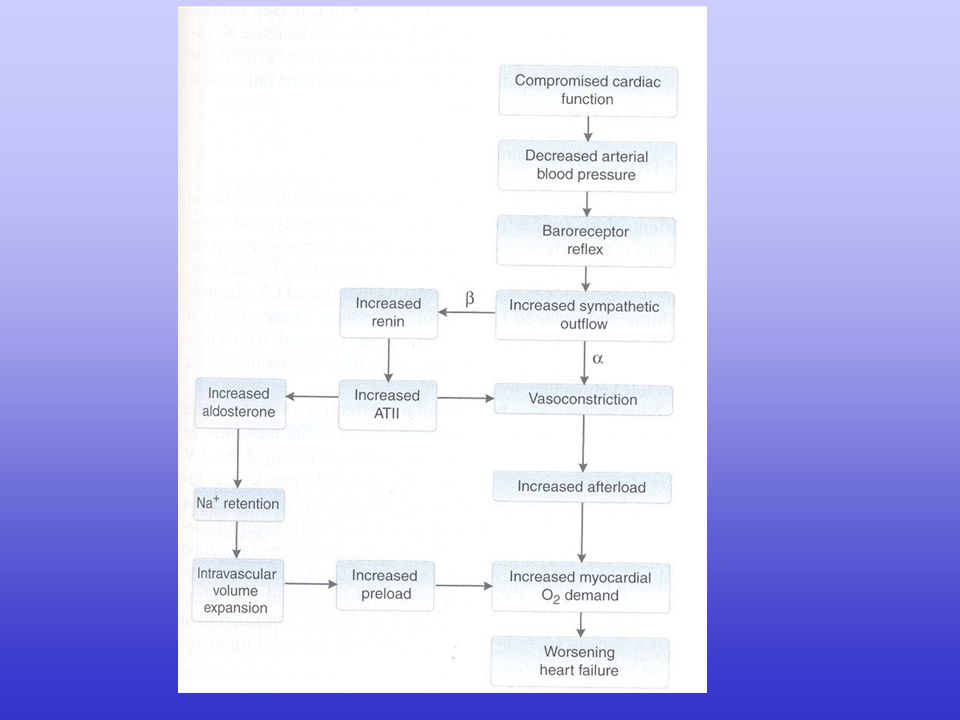
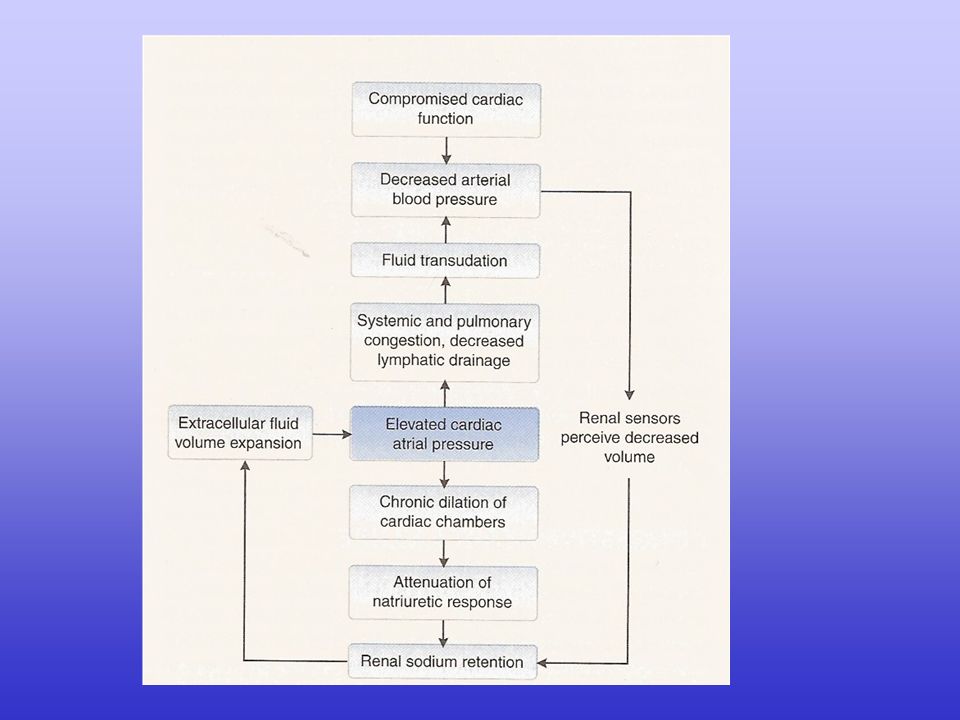
L’insufficienza cardiaca comporta alterazioni a carico di
funzionalità cardiaca status neuro-umorale funzionalità vascolare sistemica volume ematico

3
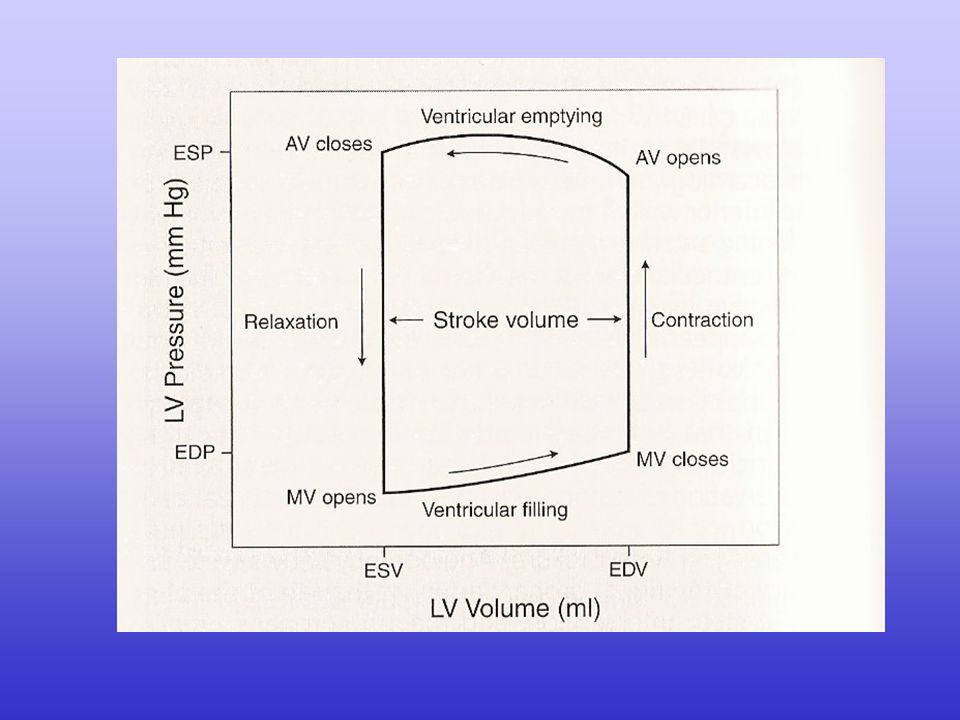
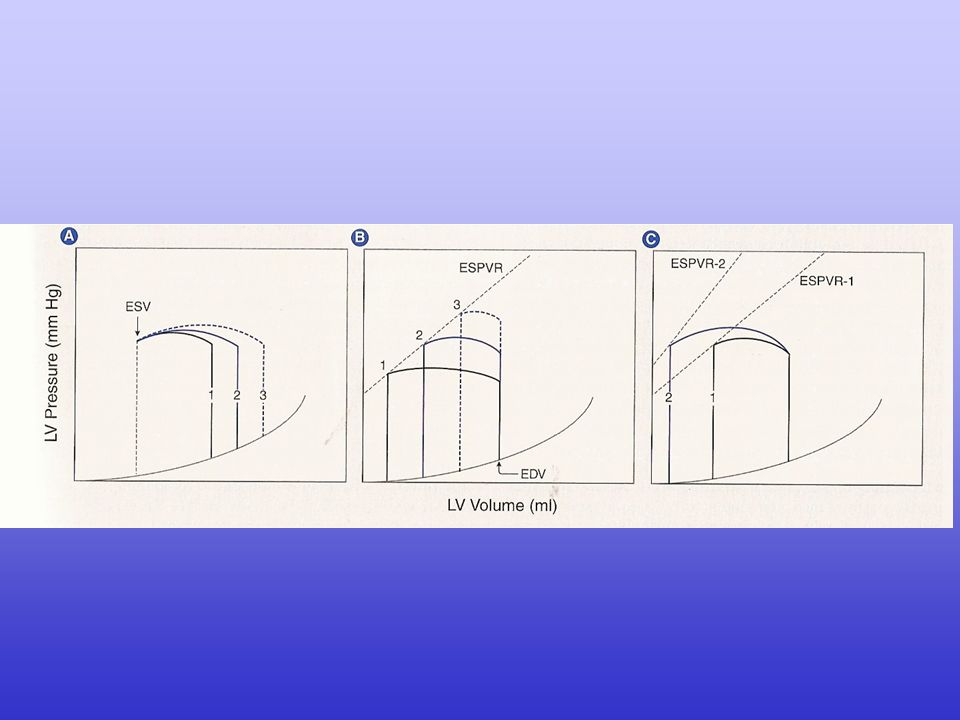
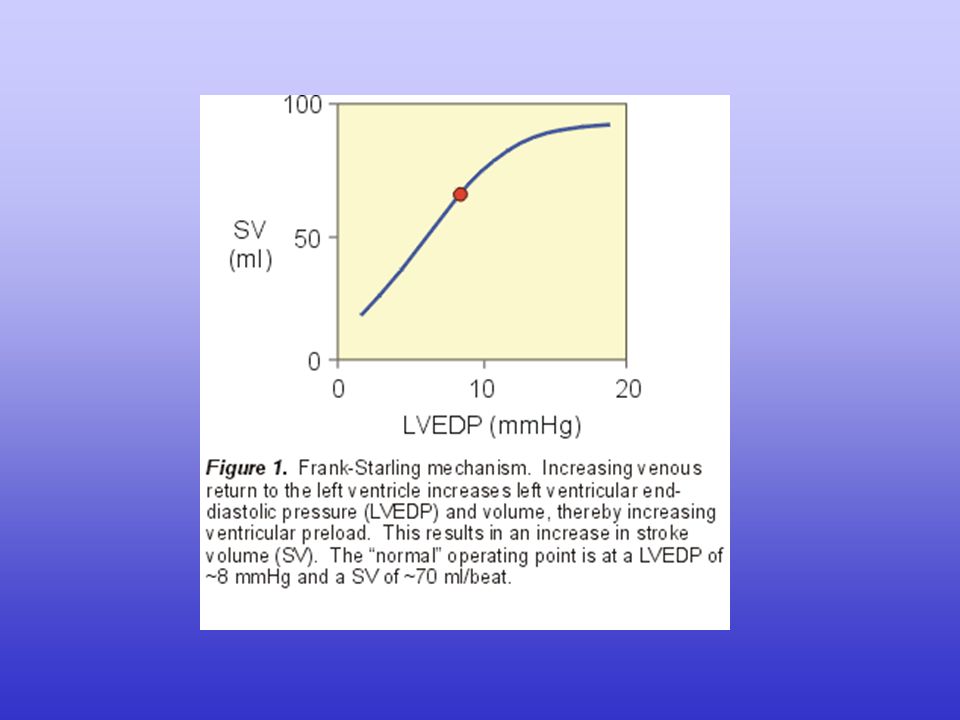
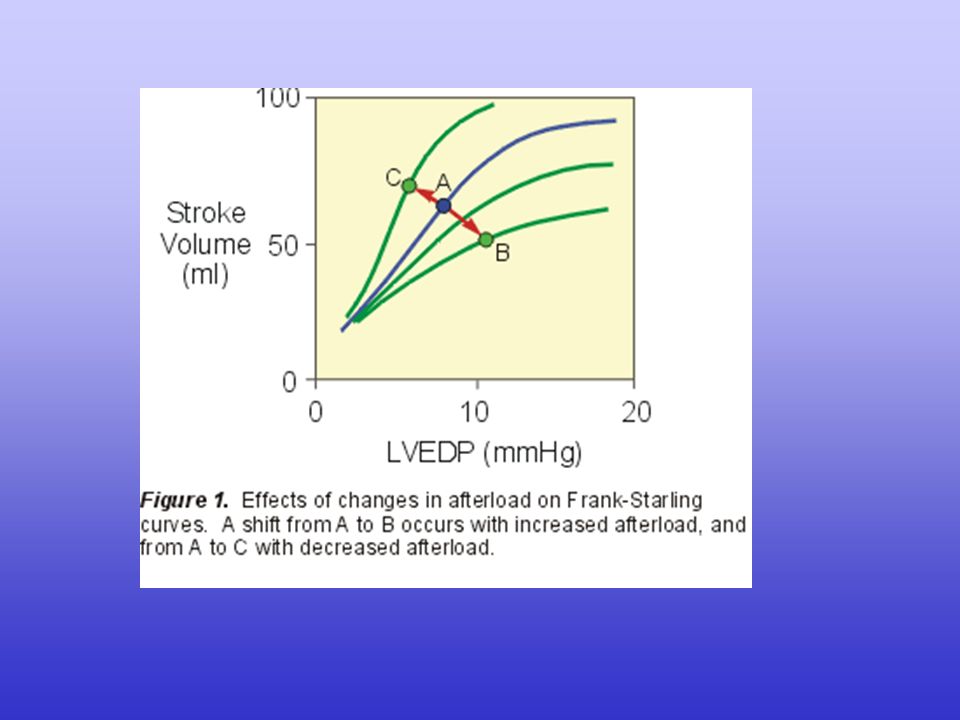
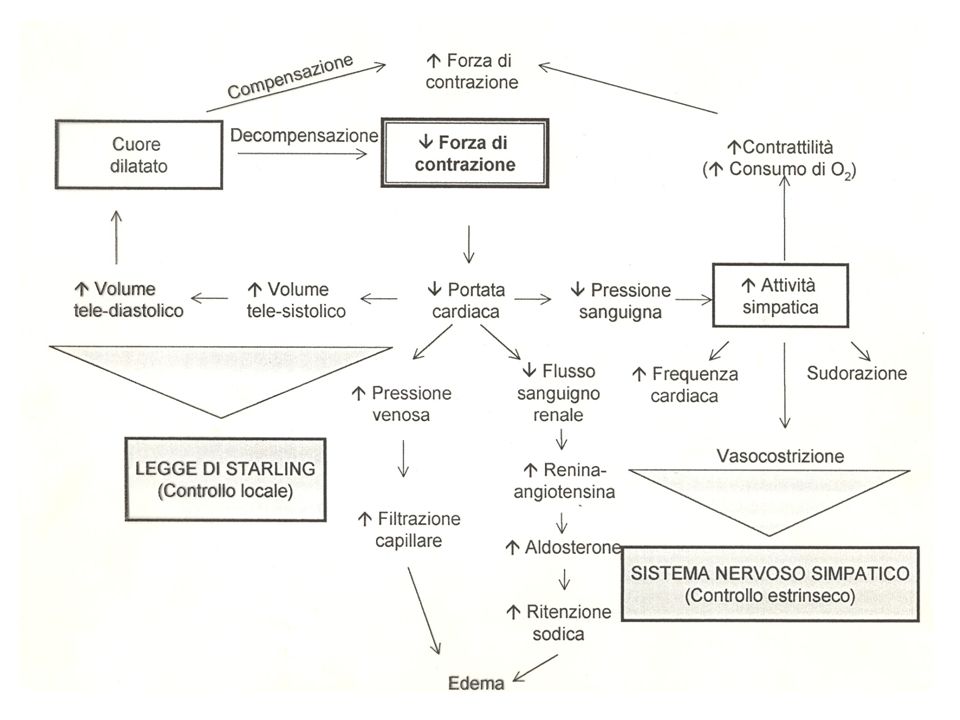
La gittata cardiaca (CO, cardiac output) dipende dalla gittata sistolica (SV, stroke volume) e dalla frequenza cardiaca (HR, heart rate): CO = SV · HR

5
A sua volta, la gittata sistolica dipende dalla differenza tra il volume telediastolico e il volume telesistolico: SV = EDV - ESV

13
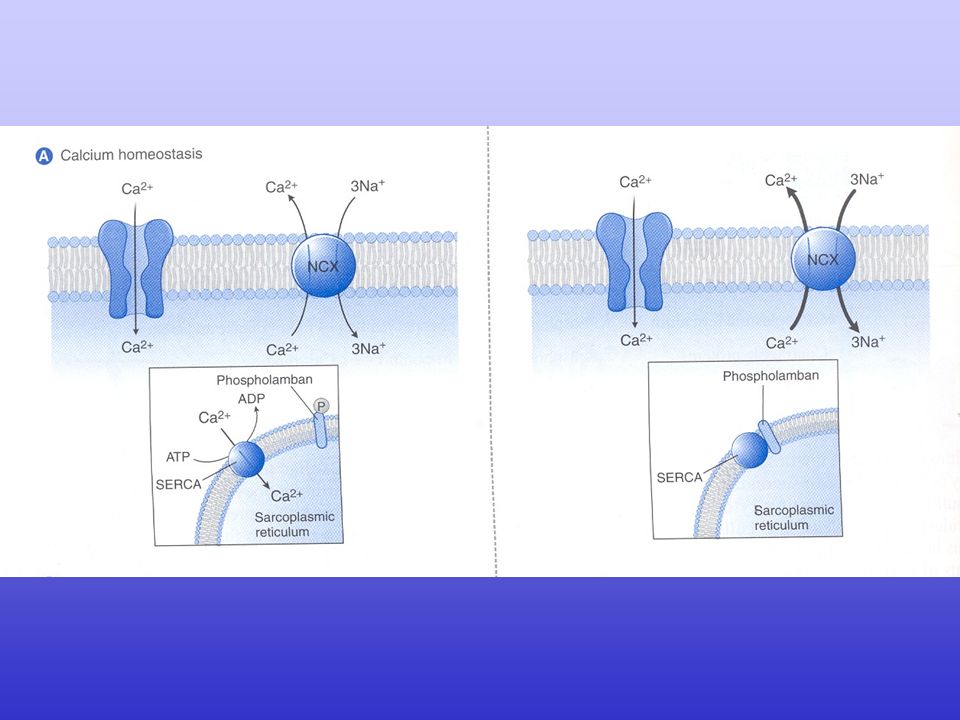
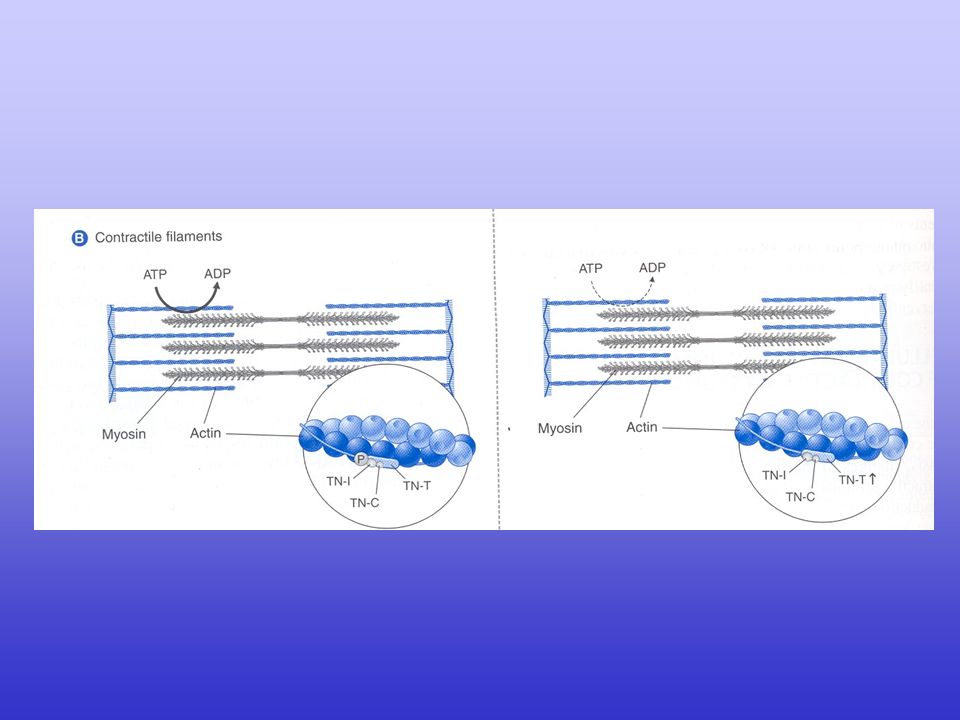
Excitation–contraction coupling in the heart
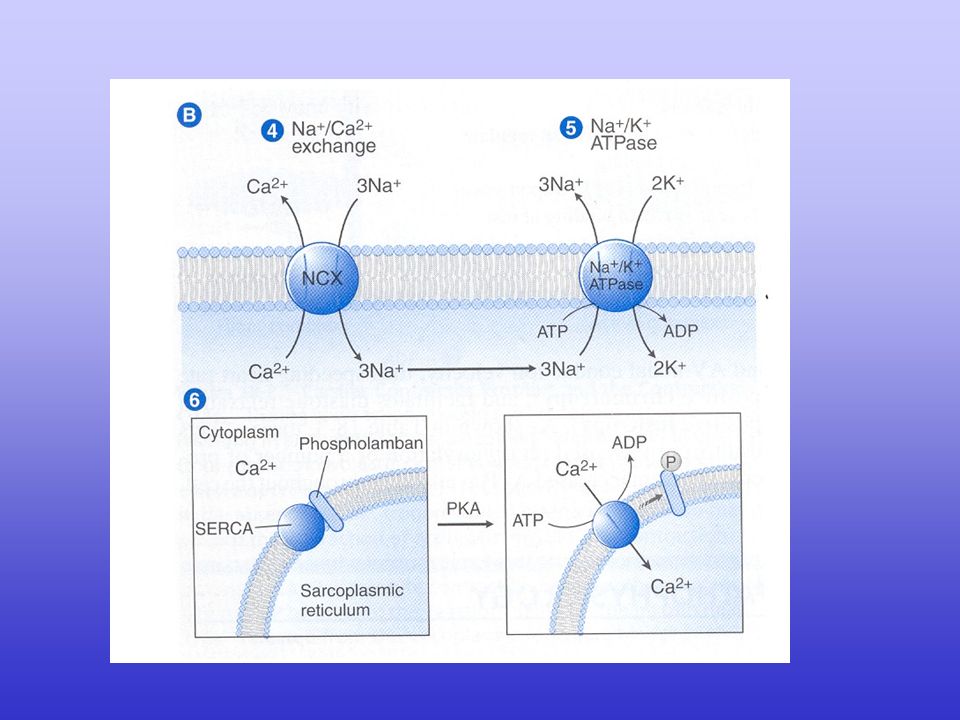
Excitation–contraction coupling in the heart. Excitation–contraction (EC) coupling in the heart involves depolarization of the transverse tubule (T-tubule), which activates voltage-gated L-type Ca2+ channels (LTCC). The small amount of Ca2+ influx through LTCC triggers a large-scale Ca2+ release from the sarcoplasmic reticulum (SR) through ryanodine receptors (RyR2). The increase in cytoplasmic Ca2+ concentration will induce muscle contraction. To enable relaxation, intracellular Ca2+ is pumped back into the SR via SR Ca2+-ATPase (SERCA2a), which is regulated by phospholamban (PLB), or extruded from the cell via the Na+/Ca2+-exchanger (NCX).
 coupling in the heart involves depolarization of the transverse tubule (T-tubule), which activates voltage-gated L-type Ca2+ channels (LTCC). The small amount of Ca2+ influx through LTCC triggers a large-scale Ca2+ release from the sarcoplasmic reticulum (SR) through ryanodine receptors (RyR2). The increase in cytoplasmic Ca2+ concentration will induce muscle contraction. To enable relaxation, intracellular Ca2+ is pumped back into the SR via SR Ca2+-ATPase (SERCA2a), which is regulated by phospholamban (PLB), or extruded from the cell via the Na+/Ca2+-exchanger (NCX).")
15
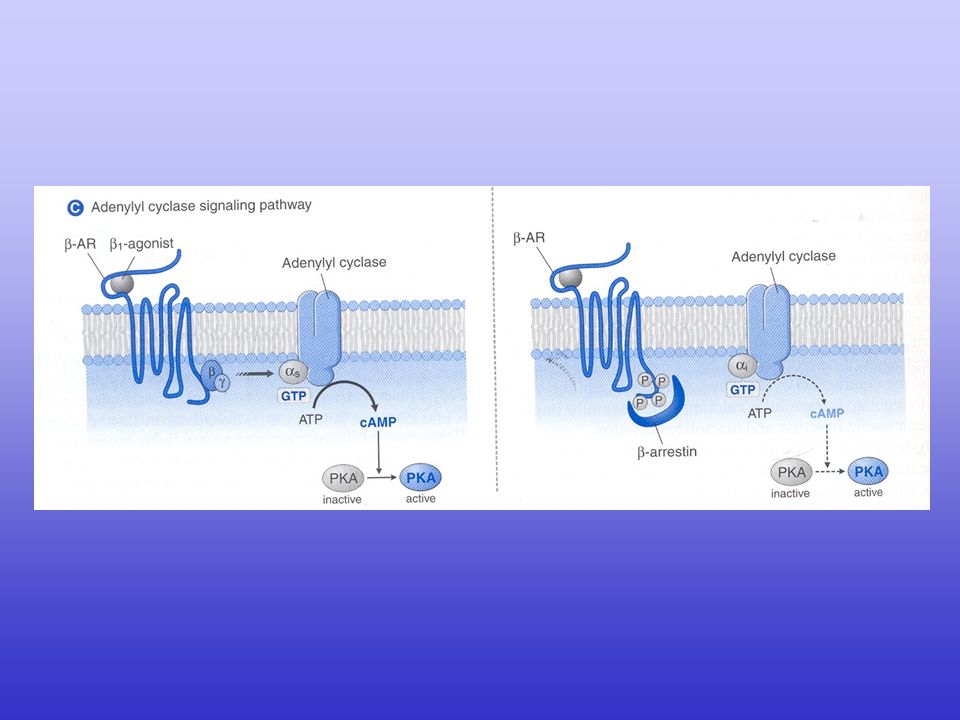
Regulation of intracellular Ca2+ signalling in the heart
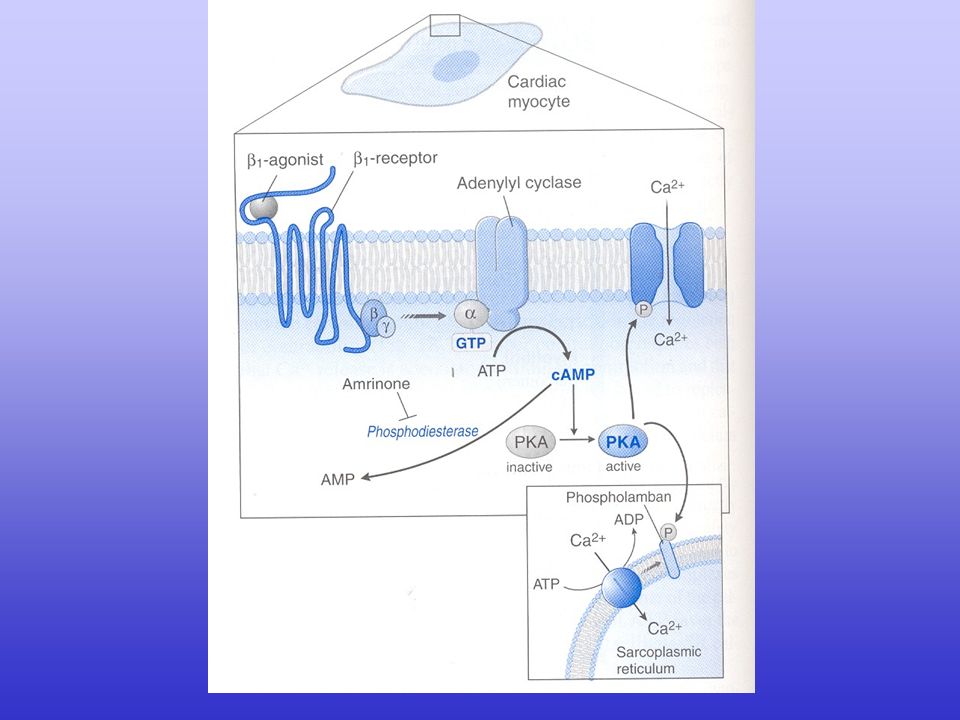
Regulation of intracellular Ca2+ signalling in the heart. Several intracellular signalling pathways can increase the gain of the excitation–contraction coupling system. Agonist-activation of the -adrenoceptor allows for activation of adenylate cyclase (AC) via specific G proteins. The subsequent generation of cAMP activates protein kinase A (PKA), which can be targeted to L-type Ca2+ channel (LTCC) via A-kinase anchoring protein (AKAP), and ryanodine receptor-2 (RyR2) and Na+/Ca2+ exchanger (NCX) via muscle AKAP (mAKAP), respectively. Faster heart rates increase the average cytosolic Ca2+ concentration, which activates Ca2+/calmodulin-dependent protein kinase (CaMKII). CaMKII can phosphorylate LTCC, RyR2 (to which CaMKII is directly targeted) and phospholamban (PLB). Activation of the angiotensin-II receptor (AT-IIR), -adrenoceptor or endothelin-1 receptor (ET-1R) activates phospholipase C (PLC) via specific G proteins, which in turn activate protein kinase C (PKC- ). PKC- can directly phosphorylate protein phosphatase inhibitor-1 (I-1), augmenting the activity of the protein phosphatase-1 (PP1) and causing hypophosphorylation of PLB. SERCA2a, sarcoplasmic reticulum ATP-ase.
 via specific G proteins. The subsequent generation of cAMP activates protein kinase A (PKA), which can be targeted to L-type Ca2+ channel (LTCC) via A-kinase anchoring protein (AKAP), and ryanodine receptor-2 (RyR2) and Na+/Ca2+ exchanger (NCX) via muscle AKAP (mAKAP), respectively. Faster heart rates increase the average cytosolic Ca2+ concentration, which activates Ca2+/calmodulin-dependent protein kinase (CaMKII). CaMKII can phosphorylate LTCC, RyR2 (to which CaMKII is directly targeted) and phospholamban (PLB). Activation of the angiotensin-II receptor (AT-IIR), -adrenoceptor or endothelin-1 receptor (ET-1R) activates phospholipase C (PLC) via specific G proteins, which in turn activate protein kinase C (PKC- ). PKC- can directly phosphorylate protein phosphatase inhibitor-1 (I-1), augmenting the activity of the protein phosphatase-1 (PP1) and causing hypophosphorylation of PLB. SERCA2a, sarcoplasmic reticulum ATP-ase.")
24
Each molecule of this family consists of three distinct structural motifs: a steroid nucleus, a sugar moiety and a lactone moiety. The lactone moiety defines the functional class of each compound. Cardenolides contain a five-membered unsaturated butyrolactone ring, whereas bufadienolides contain a six-membered unsaturated pyrone ring. Novel therapeutic applications of cardiac glycosides Ioannis Prassas & Eleftherios P. Diamandis Nature Reviews Drug Discovery 7, (November 2008)
")
25
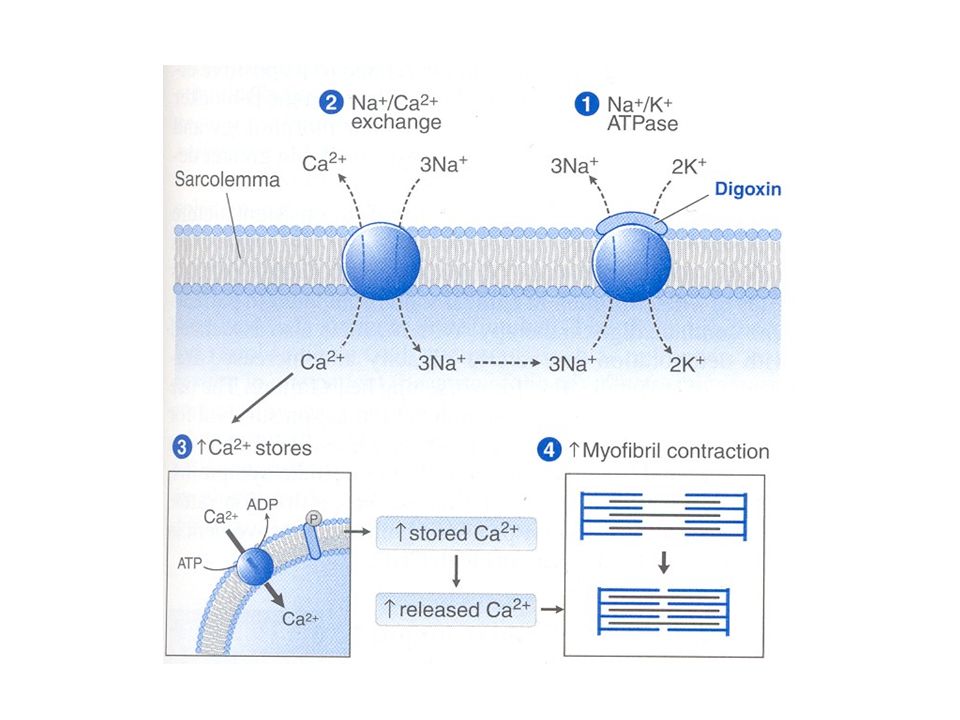
FUNZIONAMENTO DELLA POMPA Na+/K+

26
Fig. 1 NKA consists of a catalytic alpha subunit and a beta subunit that is required for the proper insertion of NKA into the plasma membrane. NKA uses the energy from ATP hydrolysis to transport Na+ out of the cell and K+ into the cell. The gradients created by this active transport will allow Na+ to enter the cell via sodium channels (1), co-transporters, such as the Na-glucose c-otransporter or the sodium amino acid co-transporter (2) and counter transporters such as the sodium hydrogen exchanger or the Na-calcium exchanger (3) without extra costs of energy. Journal of Internal Medicine Vol. 261, 1 Pages: Copyright New roles for an old enzyme: Na,K-ATPase emerges as an interesting drug target A. Aperia

27
Fig. 3. Schematic representation of the sodium pump
Fig. 3. Schematic representation of the sodium pump. The sodium pump is composed of two subunits in equimolar ratios: catalytic α (10-pass transmembrane proteins; presented in yellow) and the regulatory β (transmembrane proteins containing several glycosylation sites; presented in green). In some cells and tissues, an additional third subunit (presented in red) has been found to be associated with functional Na+/K+-ATPase. This third type of subunit is regulatory and belongs to transmembrane proteins from the FXYD family. There are four different α and three β subunit isoforms cloned to date. Sodium pump subunits are selectively expressed in various tissues and in a species-dependent manner. This figure summarizes α and β isoform expression patterns for normal human cells and tissues, as well as human FXYD distribution. The first extra-cellular loop of the α subunit (amino acids 111–122) is the most important component of the CS binding site. The basis for the relative sensitivity/insensitivity of the different α isoforms to ouabain resides mostly in two amino acids in this first extra-cellular region of the α subunit. The mouse α2 isoform, which is sensitive to ouabain, has a leucine at position 111 and an asparagine at position 122, in contrast to the ouabain-insensitive α1 isoform where arginine and aspartic acid appear at these sites. The wild-type human α1 subunit is highly sensitive to ouabain inhibition while the rat α1 subunit is essentially ouabain insensitive (1000-fold less sensitivity than the human subunit). Two key residues, glutamine Q111 and asparagine N122 are invariant in the human subunit but are respectively, mutated to arginine (R) and aspartic acid (D) in rat α1. The two key residues (Q111 and N122) are invariant in all four human α subunits, characterized by the same affinity (nM range) towards CSs. This figure is based on data reviewed in [49], [56], [57] and [61].
 and the regulatory β (transmembrane proteins containing several glycosylation sites; presented in green). In some cells and tissues, an additional third subunit (presented in red) has been found to be associated with functional Na+/K+-ATPase. This third type of subunit is regulatory and belongs to transmembrane proteins from the FXYD family. There are four different α and three β subunit isoforms cloned to date. Sodium pump subunits are selectively expressed in various tissues and in a species-dependent manner. This figure summarizes α and β isoform expression patterns for normal human cells and tissues, as well as human FXYD distribution. The first extra-cellular loop of the α subunit (amino acids 111–122) is the most important component of the CS binding site. The basis for the relative sensitivity/insensitivity of the different α isoforms to ouabain resides mostly in two amino acids in this first extra-cellular region of the α subunit. The mouse α2 isoform, which is sensitive to ouabain, has a leucine at position 111 and an asparagine at position 122, in contrast to the ouabain-insensitive α1 isoform where arginine and aspartic acid appear at these sites. The wild-type human α1 subunit is highly sensitive to ouabain inhibition while the rat α1 subunit is essentially ouabain insensitive (1000-fold less sensitivity than the human subunit). Two key residues, glutamine Q111 and asparagine N122 are invariant in the human subunit but are respectively, mutated to arginine (R) and aspartic acid (D) in rat α1. The two key residues (Q111 and N122) are invariant in all four human α subunits, characterized by the same affinity (nM range) towards CSs. This figure is based on data reviewed in [49], [56], [57] and [61].")
28
Azioni e effetti indesiderati dei glicosidi cardioattivi
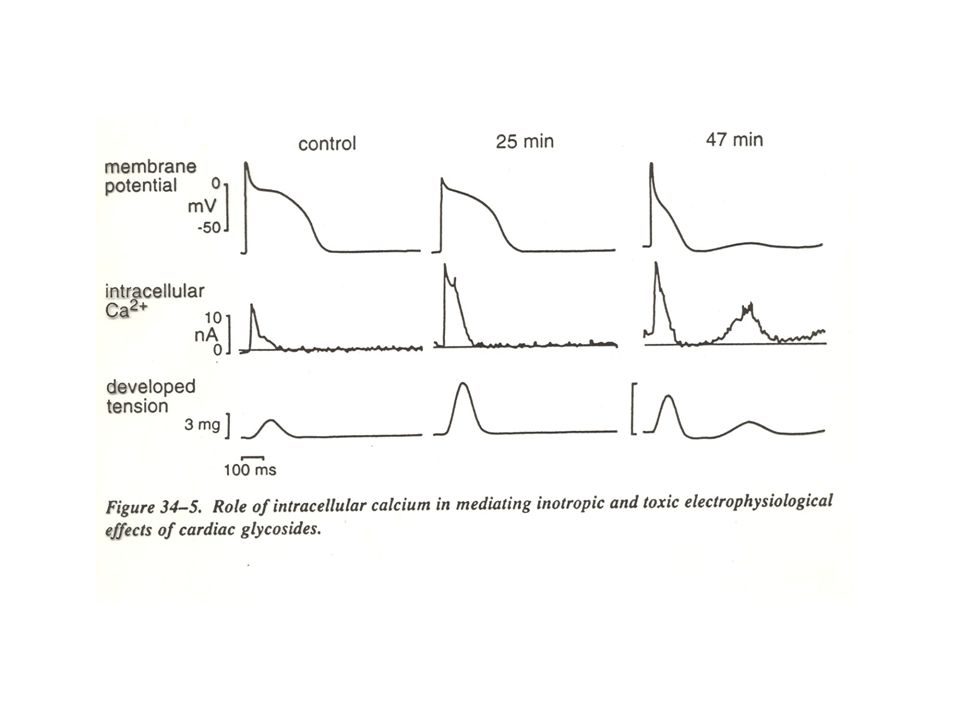
Aumento della forza contrattile cardiaca

30
Azioni e effetti indesiderati dei glicosidi cardioattivi
Aumento della forza contrattile cardiaca Rallentamento del ritmo cardiaco e riduzione della velocità di conduzione AV

31
Azioni e effetti indesiderati dei glicosidi cardioattivi
Aumento della forza contrattile cardiaca Rallentamento del ritmo cardiaco e riduzione della velocità di conduzione AV Disturbi del ritmo cardiaco: blocco della conduzione AV aumento di attività pacemaker ectopica

Presentazioni simili




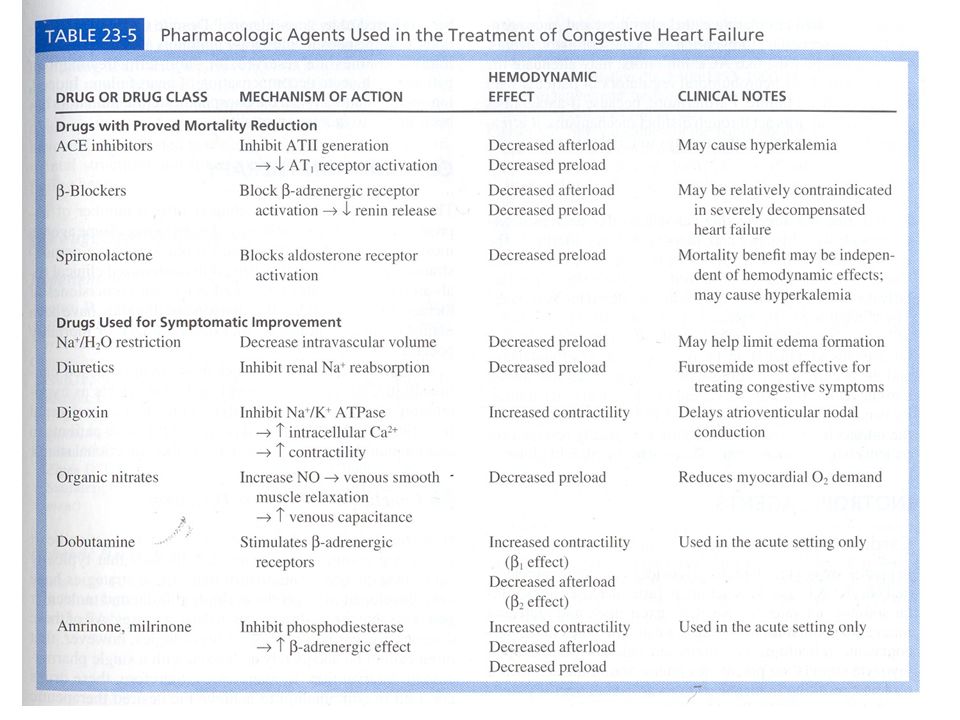











>")

>")




, specializzato per la distribuzione di:>")
