Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
CINETICHE ENZIMATICHE
METODI DI MISURA DELLE CINETICHE ENZIMATICHE

2
LO STUDIO DELLA CINETICA ALLO STATO STAZIONARIO
NON DA MOLTE INFORMAZIONI SUL MECCANISMO DI REAZIONE Allo stato stazionario possiamo solo misurare il flusso in uscita o in entrata, i quali sono uguali tra loro. Possiamo comunque ottenere alcune informazioni sul meccanismo di reazione variando le condizioni sperimentali (dipendenza dal pH)
")
3
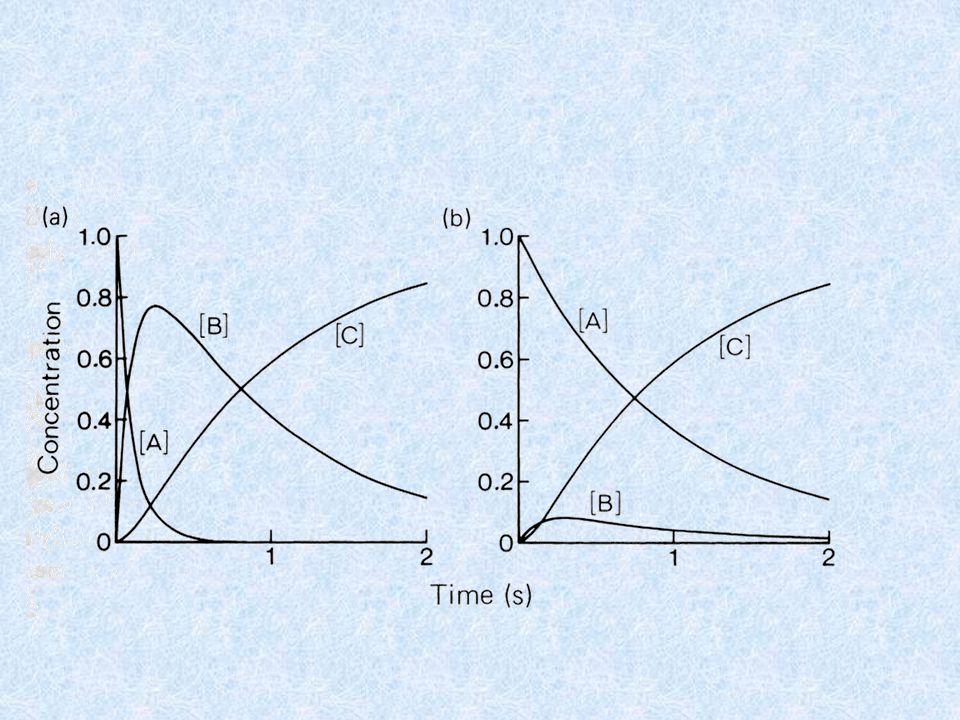
LO STUDIO DELLA CINETICA RAPIDA DELLE REAZIONI
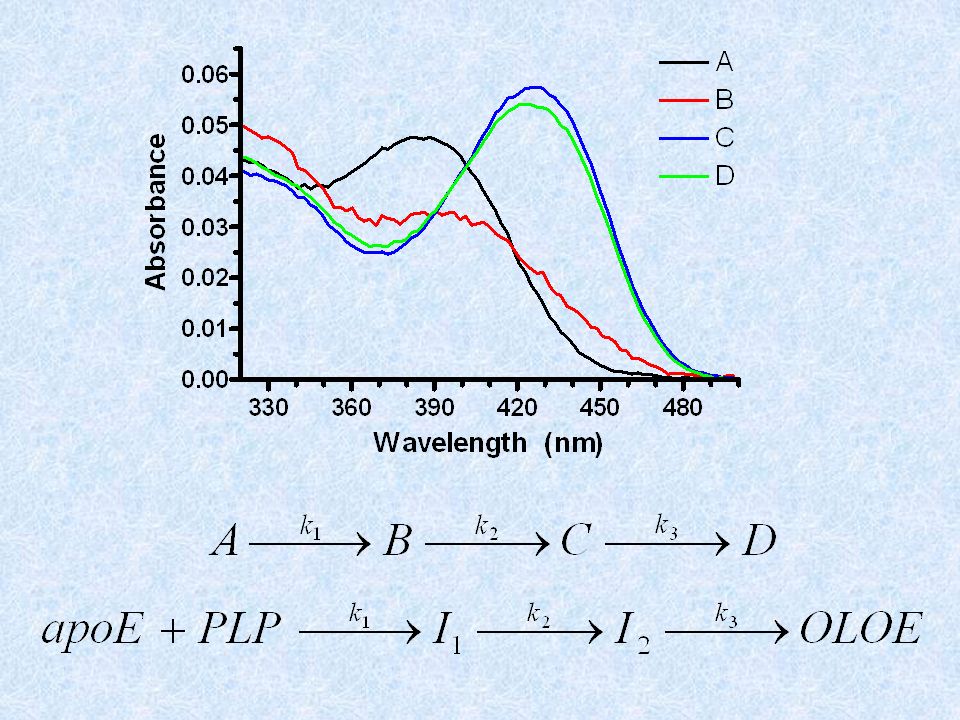
PUO’ DARE IMPORTANTI INFORMAZIONI SUL MECCANISMO DI REAZIONE E + S EX EY EZ E + P

4
(RAPID MIXING)
")
5
Tecniche di mescolamento rapido:
Il metodo del flusso continuo (1923 – H. Hartridge and F.J.W. Roughton) 1 ms 10 m/s 1 cm Tempo morto (dead time) Tempo massimo (upper time limit)
 1 ms. 10 m/s. 1 cm. Tempo morto (dead time) Tempo massimo (upper time limit)")
6
Spettrofotometro a flusso continuo
Tempo morto = 45 s Tempo massimo = 1 ms)
")
7
Spettrofotometro a flusso interrotto
Il metodo del flusso interrotto 1934 – F.J.W. Roughton 1940 B. Chance L Spettrofotometro a flusso interrotto Tempo morto = 2 ms Tempo massimo = NO

8
Il metodo del flusso “smorzato” chimicamente

9
Flash photolysis 1959 Q.H. Gibson
Flash at 347 nm ATP “ingabbiato”

10
Tecniche di rilassamento:
Salto di temperatura o di pressione Rate constant = 1/relaxation time (0.721)
")
11
Tecniche di rilassamento:
Perturbazioni sinusoidali dell’equilibrio tramite ultrasuoni

13
= kon koff [S]>>[E]
![ = kon koff [S]>>[E]](http://slideplayer.it/slide/593626/2/images/13/%EF%81%A1+%3D+kon+koff+%5BS%5D%3E%3E%5BE%5D.jpg " = kon koff [S]>>[E]")
16
E + S EX EZ E + P

19
STOPPED-FLOW DIODE ARRAY

21
SAGGI DI ATTIVITA’ ENZIMATICA
Gli enzimi possono essere usati come strumenti analitici per identificare o quantificare sostanze chimiche specifiche in soluzione. E’ possibile determinare la presenza e la quantità di un determinato enzima in un fluido biologico misurandone l’attività con substrati specifici. Possiamo determinare i parametri cinetici (KM , Kcat o attività specifica) di un particolare enzima. CONDIZIONI SPERIMENTALI DEL SAGGIO Controllo del pH con una soluzione tampone Controllo della temperatura Assenza di inibitori (agenti chelanti, metalli) { CONTINUI SAGGI DI ATTIVITA’ DISCONTINUI o A PUNTI FISSI In un saggio di attività enzimatica noi misuriamo il prodotto della reazione, più di rado misuriamo il consumo di substrato.
 di un. particolare enzima. CONDIZIONI SPERIMENTALI DEL SAGGIO. Controllo del pH con una soluzione tampone. Controllo della temperatura. Assenza di inibitori (agenti chelanti, metalli) { CONTINUI. SAGGI DI ATTIVITA’ DISCONTINUI o A PUNTI FISSI. In un saggio di attività enzimatica noi misuriamo il prodotto della reazione, più di. rado misuriamo il consumo di substrato.")
22
E + S E + P (prodotto colorato - cromoforo
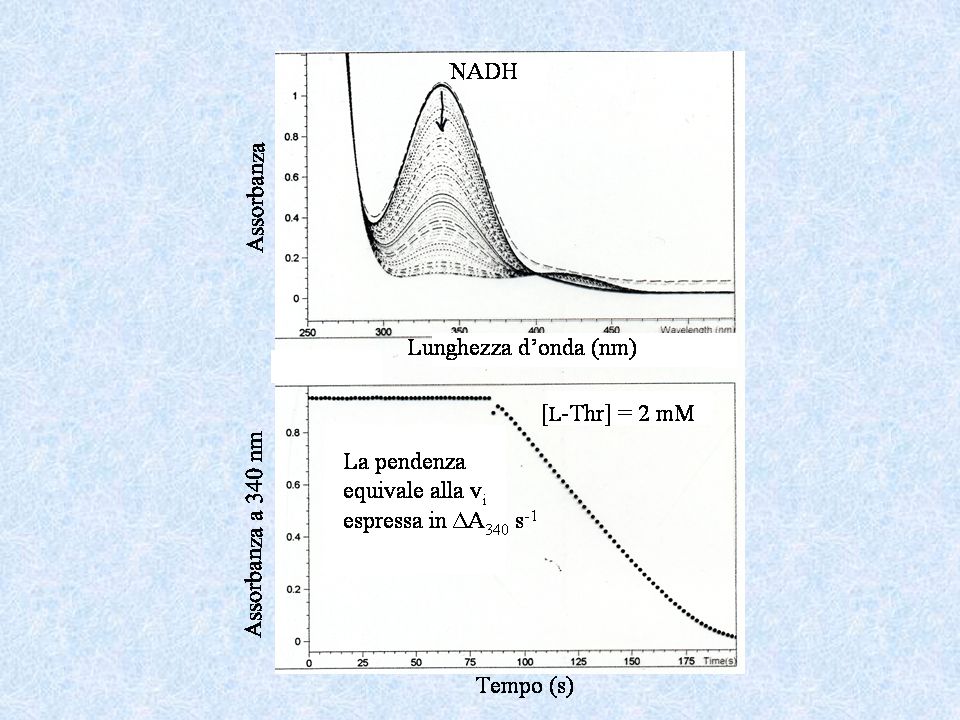
SAGGI CONTINUI Sono più accurati e meno laboriosi di quelli discontinui Per lo più usano metodi spettrofotometrici SAGGI DIRETTI E + S E + P (prodotto colorato - cromoforo o fluorescente – fluoroforo) Esempio: + NADH Piruvato Lattato Deidrogenasi (LDH) + NAD+ Lattato
 Esempio: + NADH. Piruvato. Lattato. Deidrogenasi. (LDH) + NAD+ Lattato.")
23
E1 + S E1 + P (prodotto non colorato o fluorescente)
SAGGI ACCOPPIATI E1 + S E1 + P (prodotto non colorato o fluorescente) E2 + P E2 + Q (secondo prodotto, Q, colorato) L-treonina aldolasi Esempio: + L-treonina glicina acetaldeide alcol deidrogenasi + NADH + NAD+ acetaldeide alcol etilico
 E2 + P E2 + Q (secondo prodotto, Q, colorato) L-treonina. aldolasi. Esempio: + L-treonina. glicina. acetaldeide. alcol. deidrogenasi. + NADH. + NAD+ acetaldeide. alcol etilico.")
25
[Etot] = 0,13 mM Vmax = 26 0,3 mM min-1 KM = 0,19 0,01 mM kcat = Vmax / [Etot] = 26 / 0,13 = 206 min-1
![[Etot] = 0,13 mM Vmax = 26 0,3 mM min-1. KM = 0,19 0,01 mM.](http://slideplayer.it/slide/593626/2/images/25/%5BEtot%5D+%3D+0%2C13+mM+Vmax+%3D+26+%EF%82%B1+0%2C3+mM+min-1.+KM+%3D+0%2C19+%EF%82%B1+0%2C01+mM..jpg "kcat = Vmax / [Etot] = 26 / 0,13 = 206 min-1.")
26
SAGGI DISCONTINUI Alcuni sistemi non possono essere saggiati in modo continuo Perché durante la reazione non si formano o consumano cromofori o fluorofori e le condizioni per far sviluppare colore o fluorescenza sono diverse da quelle del saggio. Non è possibile far sviluppare colore o fluorescenza. La misura del prodotto (o del substrato consumato) devono essere fatte con altri sistemi, ad esempio HPLC, saggi biologici o immunologici. Presenza di un fondo ( background ) elevato; ad esempio di assorbanza dovuta alle proteine in un estratto crudo o di altre molecole che assorbono alla lunghezza d’onda che a noi interessa. In questo caso dobbiamo condurre il saggio e poi eliminare la fonte di disturbo.
 devono essere fatte con altri sistemi, ad esempio HPLC, saggi biologici o immunologici. Presenza di un fondo ( background ) elevato; ad esempio di assorbanza dovuta. alle proteine in un estratto crudo o di altre molecole che assorbono alla lunghezza. d’onda che a noi interessa. In questo caso dobbiamo condurre il saggio e poi. eliminare la fonte di disturbo.")
27
ESEMPI DI SAGGI DISCONTINUI
PRELIEVO DI ALIQUOTE AD INTERVALLI DI TEMPO ENZIMA + SUBSTRATO REAZIONE IN CORSO La reazione viene bloccata con acido, denaturanti delle proteine, calore, inibitori dell’enzima Il prodotto di reazione viene quantificato con vari metodi E1 + S E1 + P P-X metodi colorimetrici Quantificazione con HPLC Trasformazione in un composto che può essere quantificato colorimetricamente o in altro modo 1a FASE 2a FASE X E2 Metodi radioisotopici

28
Esempio: 1a FASE Glutamato 1-semialdeide (GSA) aminolevulinato (ALA) 2a FASE GSA + DMBA COMPOSTO COLORATO
 5-aminolevulinato (ALA) 2a FASE GSA + DMBA COMPOSTO COLORATO.")
29
Metodi radioisotopici
Si basano sull’utilizzo di forme radiomarcate di un substrato Sono metodi molto sensibili ma limitati alle applicazioni in cui è possibile separare facilmente i substrati dai prodotti Esempio: misura dell’attività di un’aminotrasferasi + 1a FASE Miscela di reazione lavaggio eluizione Conteggio radiattività 2a FASE (separazione cromatografica e conteggio della radiattività) colonna cromatografica a scambio anionico aminoacidi chetoacidi
 colonna. cromatografica. a scambio anionico. aminoacidi. chetoacidi.")
30
{ { INATTIVAZIONE DEGLI ENZIMI REVERSIBILE IRREVERSIBILE E + P1
COMPETITIVA INCOMPETITIVA MISTA REVERSIBILE { ASPECIFICA PER IL TIPO DI ENZIMA BASATA SUL MECCANISMO DI REAZIONE DELL’ENZIMA (inibitori suicidi) IRREVERSIBILE E + P1 E + I EI E + P2 EX EY
 IRREVERSIBILE. E + P1. E + I EI. E + P2. EX. EY.")
34
La parete batterica contiene D-aminoacidi (D-alanina ad es
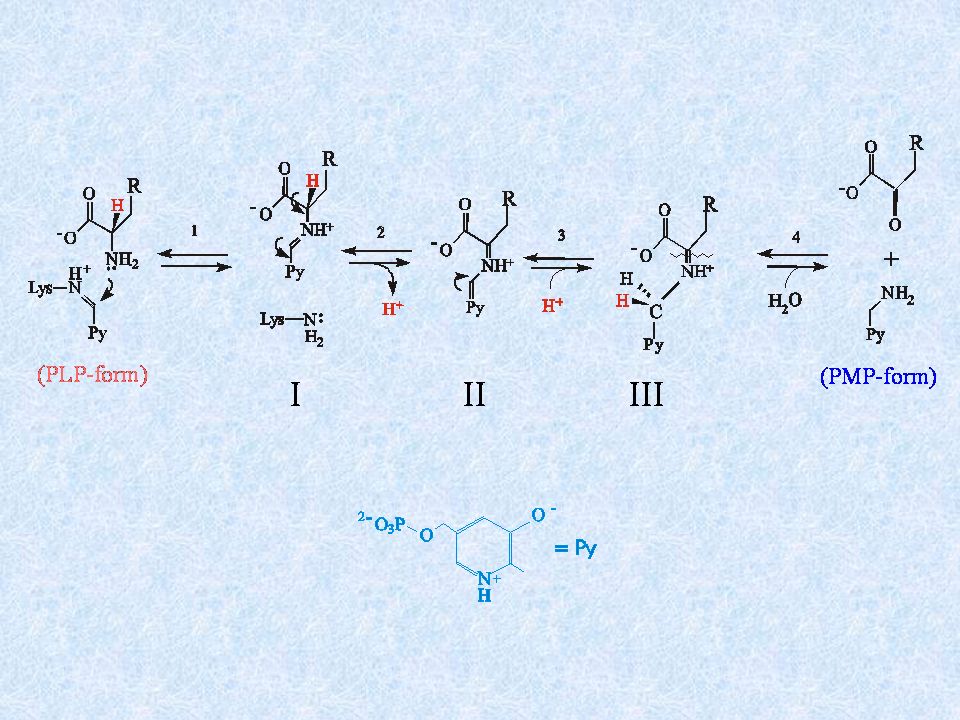
La parete batterica contiene D-aminoacidi (D-alanina ad es.) come costituenti dei ponti polipeptidici che connettono le catene di carboidrati. Questi vengono ottenuti dagli L-aminoacidi tramite reazioni di racemizzazione (catalizzate da racemasi). In alcuni batteri, il D-glutammato che viene incorporato nella parete batterica non è sintetizzato per racemizzazione ma tramite una reazione di transaminazione: D-alanina + a-chetoglutarato piruvato + D-glutammato Questa reazione è catalizzata da un enzima dipendente dal piridossal fosfato: la D-aminoacido aminotrasferasi
 come costituenti. dei ponti polipeptidici che connettono le catene di carboidrati. Questi vengono ottenuti dagli L-aminoacidi tramite reazioni di racemizzazione. (catalizzate da racemasi). In alcuni batteri, il D-glutammato che viene incorporato nella parete batterica. non è sintetizzato per racemizzazione ma tramite una reazione di transaminazione: D-alanina + a-chetoglutarato piruvato + D-glutammato. Questa reazione è catalizzata da un enzima dipendente dal piridossal fosfato: la D-aminoacido aminotrasferasi.")
35
+ E-PLP + INTERMEDIO AMINOACRILATO D-b-cloroalanina (inibitore)
Il Cloro è un buon gruppo uscente nelle reazioni di sostituzione nucleofila + E-PLP D-alanina (substrato naturale) INTERMEDIO AMINOACRILATO Verso l’inibizione + Normale reazione di transaminazione
 INTERMEDIO. AMINOACRILATO. Verso l’inibizione. + Normale reazione. di transaminazione.")
36
MECCANISMO DI INATTIVAZIONE
INTERMEDIO AMINOACRILATO ENZIMA INATTIVATO

37
La VIGABATRINA: un farmaco contro l’epilessia.
L’acido g-aminobutirrico (GABA) è uno dei principali neurotrasmettitori inibitori del sistema nervoso centrale glutammato decarbossilasi L-glutammato GABA + + GABA aminotrasferasi GABA a-chetoglutarato Succinico semialdeide L-glutammato
 è uno dei principali neurotrasmettitori inibitori del. sistema nervoso centrale. glutammato. decarbossilasi. L-glutammato. GABA. + + GABA. aminotrasferasi. GABA. a-chetoglutarato. Succinico semialdeide. L-glutammato.")
38
L’epilessia può essere trattata aumentando la concentrazione di GABA
nel sistema nervoso centrale. Un metodo per ottenere questo risultato è quello di inibire in modo specifico la GABA aminotrasferasi GABA 4-amino-5-esanoato VIGABATRINA La VIGABATRINA, un analogo del GABA, è un efficiente inibitore della GABA aminotrasferasi ed è attualmente utilizzata con successo nel trattamento dell’epilessia

39
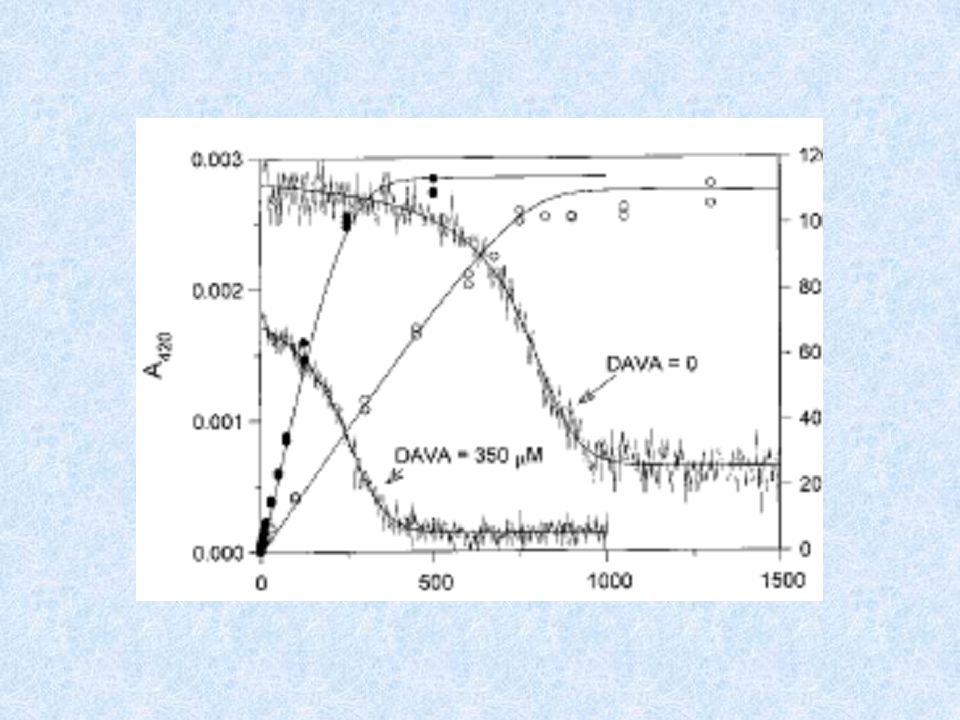
MECCANISMO DI INIBIZIONE DELLA VIGABATRINA
75% ENZIMA INATTIVATO 25%

40
a-difluorometilornitina
L’EFLORNITINA: un farmaco contro la malattia del sonno L’ornitina decarbossilasi è un enzima essenziale per la produzione della putrescina, un precursore delle poliamine + E-PLP ornitina a-difluorometilornitina o eflornitina + E-PLP putrescina

41
MECCANISMO DI INIBIZIONE DELL’EFLORNITINA
ENZIMA INATTIVATO

Presentazioni simili

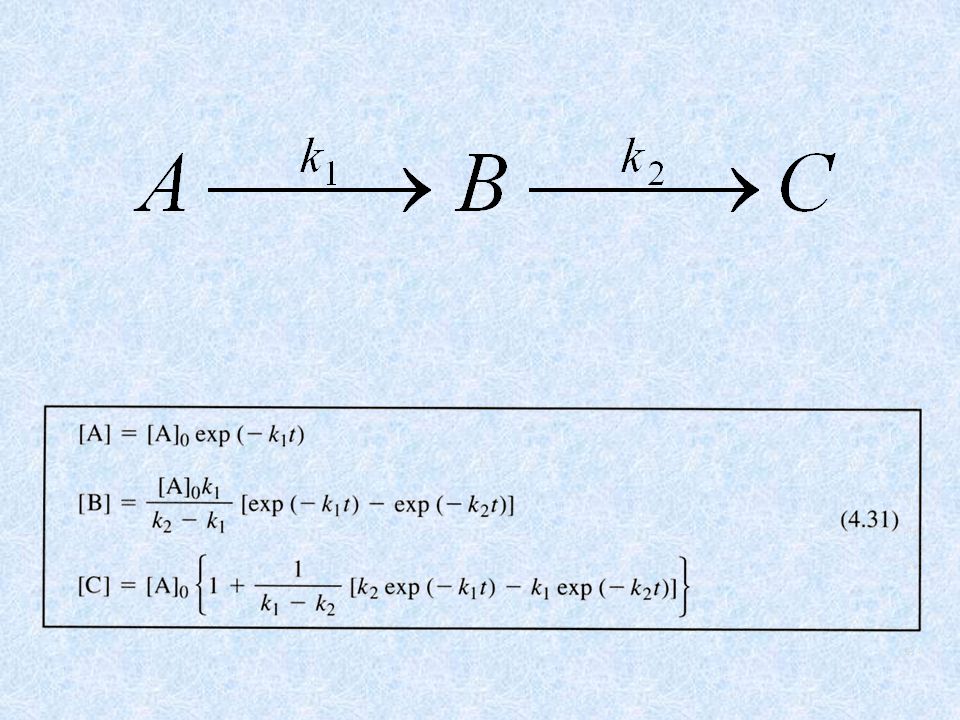
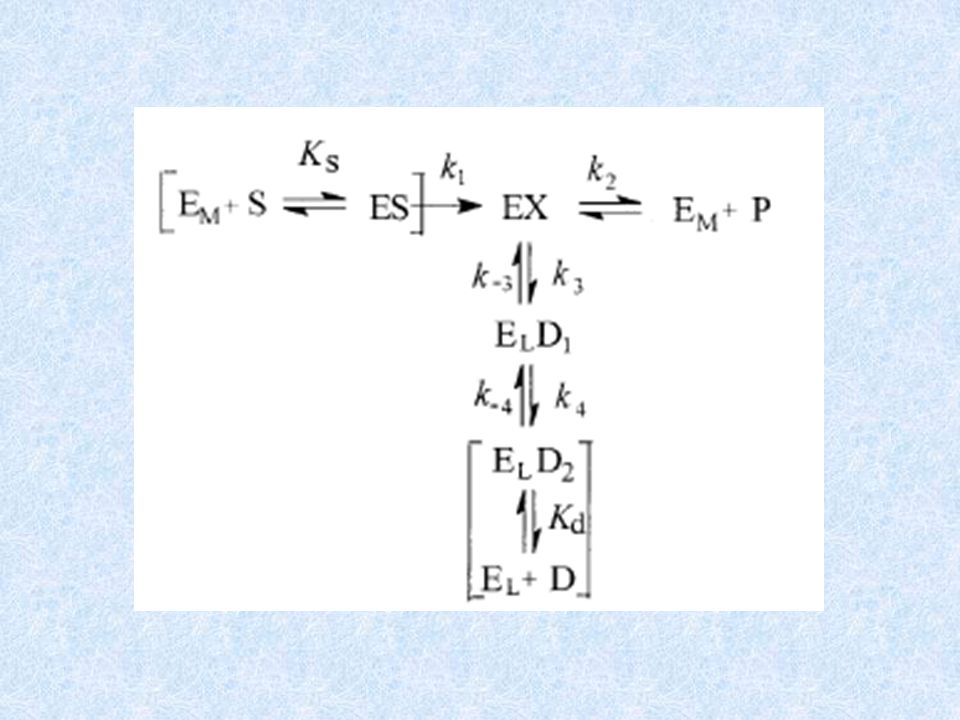







>")







 è un enzima citoplasmatico con un'ampia distribuzione nei tessuti, dove catalizza.>")


>")


