Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Trasporto degli elettroni e fosforilazione ossidativa

2
C6H12O6 + 6O2 6CO2 + 6 H2O C6H12O6 + 6O2 6CO2 + 24 H+ + 24 e-
Gli organismi aerobici durante il processo di ossidazione dei carburanti metabolici consumano ossigeno e producono biossido di carbonio: C6H12O6 + 6O2 6CO2 + 6 H2O suddivisa in due semireazioni: C6H12O6 + 6O2 6CO H e- 6O H e- 12 H2O

3
Le 12 coppie di e- rilasciate durante l’ossidazione del glucosio non sono trasferite direttamente all’O2 ma ai coenzimi NAD+ e FAD tramite i coenzimi, gli e- passano alla catena di trasporto degli e-

4
Durante il processo di trasporto degli elettroni accadono i eguenti eventi:
NADH e FADH2 sono riossidati a NAD+ e FAD il trasferimento degli e- partecipa all’ossidazione-riduzione di 10 centri redox durante il trasferimento degli e-, dal mitocondrio vengono espulsi dei protoni con la generazione di un gradiente protonico attraverso la membrana mitocondriale.

6
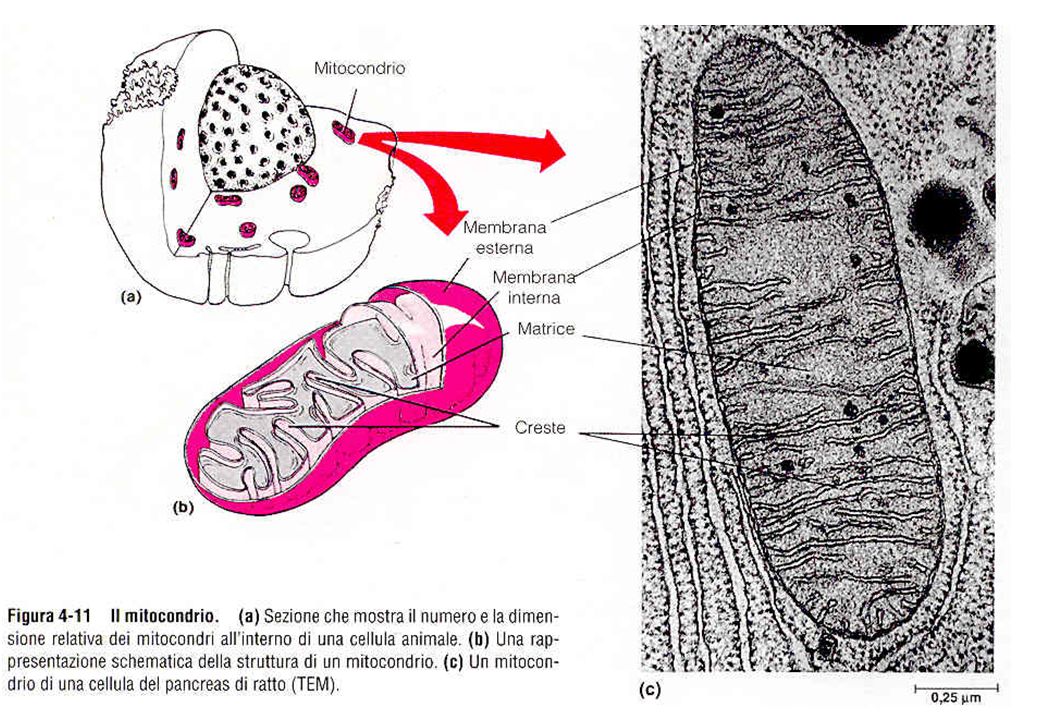
Struttura di un mitocondrio
50% lipidi, 50% proteine 80% proteine

7
NAD+, NADH, NADP+, NADPH, FAD, FADH2
La membrana mitocondriale interna è priva di una proteina trasportatrice del NADH Nel mitocondrio vengono trasportati soltanto gli elettroni del NADH citosolico mediante ingegnosi sistemi “ navetta”

8
Sistemi navetta di substrati:
si servono di isoforme citoplasmatiche e mitocondriali dello stesso enzima

9
Shuttle malato – aspartato (fegato, muscolo cardiaco)
GOT cit reaz. di transamminazione 4 1 mmi 3 2 reaz. di transamminazione GOT mit. malato deidrogenasi (ciclo di Krebs) (reaz. di ossidazione)
 (reaz. di ossidazione)")
10
Glicerofosfato del muscolo alare degli insetti
Il sistema navetta del Glicerofosfato del muscolo alare degli insetti trasferimento di una coppia di e- glicerolo 3-P deidrogenasi FAD dip. isozima

11
Meccanismo conformazionale del traslocatore ADP-ATP o adenina nucleotide traslocasi che trasporta l’ATP fuori dalla matrice scambiandolo con l’ADP (dimero costituito da due subunità identiche di 30 kD) Il Pi che è necessario anche per la sintesi di ATP nella matrice, viene importato dal citosol mediante un sistema di simporto Pi-H+ . Il gradiente protonico transmembrana generato dalla catena di trasporto degli elettroni presente sulla membrana mitocondriale interna non fornisce soltanto la forza termodinamica necessaria per la sintesi di ATP ma promuove anche il trasporto dei suoi materiali di partenza , ADP e Pi.

12
I due sistemi di trasporto mitocondriale per il Ca 2+
I MT funzionano da tamponi per il calcio citosolico. Importanza della distribuzione intracellulare del calcio: contrazione muscolare, trasmissione neuronale, secrezione, azione degli ormoni L’entrata nella matrice è promossa dal potenziale di membrana negativo che attrae gli ioni positivi. La velocità dipende dalla [Ca++] esterna Se la [Ca++]cit antiporto con ioni Na+ = aumenta l’entrata Se la [Ca++] cit = diminuisce l’entrata Quando la [Ca++] citoplasmatica aumenta per un lavoro muscolare sostenuto, [Ca++] aumenta anche nella matrice mitoc., attivando gli enzimi del ciclo di Krebs che porta a un aumento dei livelli di NADH, la cui riossidazione tramite il sistema di trasporto degli elettroni genera l’ATP necessario per la contrazione.

13
I trasportatori degli elettroni che portano gli elettroni dal NADH e dal FADH all’O2 sono associati alla membrana mitocondriale interna. Alcuni di questi centri redox sono mobili, altri sono invece componenti di complessi di proteine integrali di membrana. La sequenza dei trasportatori degli elettroni riflette grossolanamente i loro potenziali di riduzione relativi, cosicchè il processo di trsporto degli elettroni è complessivamente esoergonico.

14
La via più importante per la formazione di ATP, presente sia negli organismi autotrofi che in quelli eterotrofi, è la fosforilazione ossidativa: questo processo è fortemente endoergonico (cioè necessita di una grande quantità di energia, DG > 0) Il meccanismo più rilevante per la sintesi di ATP da ADP e Pi utilizza energia derivante da gradienti elettrochimici. Questa energia viene liberata grazie alla presenza nella cellula (mitocondrio) di catene trasportatrici di elettroni (processo redox fortemente esoergonico, DG < 0)
 di catene trasportatrici di elettroni (processo redox fortemente esoergonico, DG < 0)")
15
La catena respiratoria libera energia grazie alla differenza tra i potenziali di riduzione tra il donatore (NADH + H+), complessi della catena respiratoria e l’accettore (O2) di elettroni. Gran parte di questa energia viene impiegata dall’ATP sintasi per convertire l’ADP in ATP

16
Il flusso di elettroni può produrre un lavoro biologico:
Nel “circuito che potenzia un motore”, la fonte di e- può essere una batteria che contiene due specie chimiche che hanno una diversa affinità per gli e- . Il flusso elettronico procederà spontaneamente nel circuito guidato da una forza proporzionale alla differenza tra le due affinità detta forza elettromotrice (fem). La fem può generare un lavoro se al circuito viene collegato un trasduttore energetico.
. La fem può generare un lavoro se al circuito viene collegato un trasduttore energetico.")
17
In un analogo “circuito biologico” la fonte di elettroni è un composto ridotto come per es. il glucosio. Quando viene ossidato enzimaticamente sono rilasciati e- che fluiscono spontaneamente attraverso una serie di trasportatori intermedi fino a raggiungere una specie chimica con un’elevata affinità per gli e- come l’ossigeno. La forza motrice che si genera fornisce energia ad una varietà di trasduttori molecolari (enzimi e proteine) che compiono un lavoro biologico.
 che compiono un lavoro biologico.")
18
sostanza organica + O2 D CO2 + H2O + energia
L’ossidazione biologica completa di sostanze organiche nutrienti crea un flusso di elettroni che procede verso l’ossigeno mediante una serie di trasportatori di elettroni intermedi che si comportano come un ponte a cestelli: reazione redox esoergonica: gli elettroni passano spontaneamente da un trasportatore a bassa affinità per gli elettroni ad un altro con affinità maggiore e così via. La reazione complessiva è: sostanza organica + O2 D CO2 + H2O + energia

19
reazioni di ossido riduzione
Il sistema mitocondriale di trasporto degli e- non è altro che una sequenza di reazioni di ossido riduzione Durante le reazioni redox, gli elettroni trasferiti possono passare direttamente dai donatori agli accettori: Fe2+ + Cu2+ D Fe3+ + Cu+ In alternativa, gli elettroni possono essere trasferiti da un donatore a un accettore mediante un trasportatore di elettroni: XH2 + Y D X +Y H2 NADH + H+ + FAD NAD+ + FADH2

20
Un atomo di idrogeno neutro è in grado di trasferire un singolo elettrone:
H D e- + H+ Lo ione negativo idruro (H-) è invece in grado di trasferire due elettroni: H- D 2e- + H+
 è invece in grado di trasferire due elettroni: H- D 2e- + H+")
21
Le forme ossidate e ridotte dei composti sono dette coppie redox.
donatore di e- (agente riducente) accettore di e- (agente ossidante) La facilità con la quale gli e- passano dal riducente all’ossidante è espressa quantitativamente dal valore del potenziale di ossido-riduzione del sistema.
 accettore di e- (agente ossidante) La facilità con la quale gli e- passano dal riducente all’ossidante è espressa quantitativamente dal valore del potenziale di ossido-riduzione del sistema.")
22
In una coppia redox il riducente con un potenziale di ox-red molto negativo cede facilmente i suoi e- a coppie con potenziali redox meno negativi o più positivi Viceversa un forte ossidante (caratterizzato da un potenziale redox molto positivo) mostra una grande affinità per gli e-.
 mostra una grande affinità per gli e-.")
23
Il potenziale di riduzione standard (E0) è una misura quantitativa dell'affinità di un accettore per gli elettroni in una semi-reazione. Per convenzione internazionale, alla semi-reazione H+ + e- D 1/2H2 viene assegnato un potenziale di riduzione standard pari a zero, quando la concentrazione di ioni H+ è 1 M e H2 è presente alla pressione parziale di 101,3 kPa. Questa semi-reazione viene usata come riferimento per la determinazione dei potenziali delle altre semi-reazioni.

24
Il più forte agente ossidante cioè ha elevata affinità per gli elettroni

25
complesso I: NADH deidrogenasi
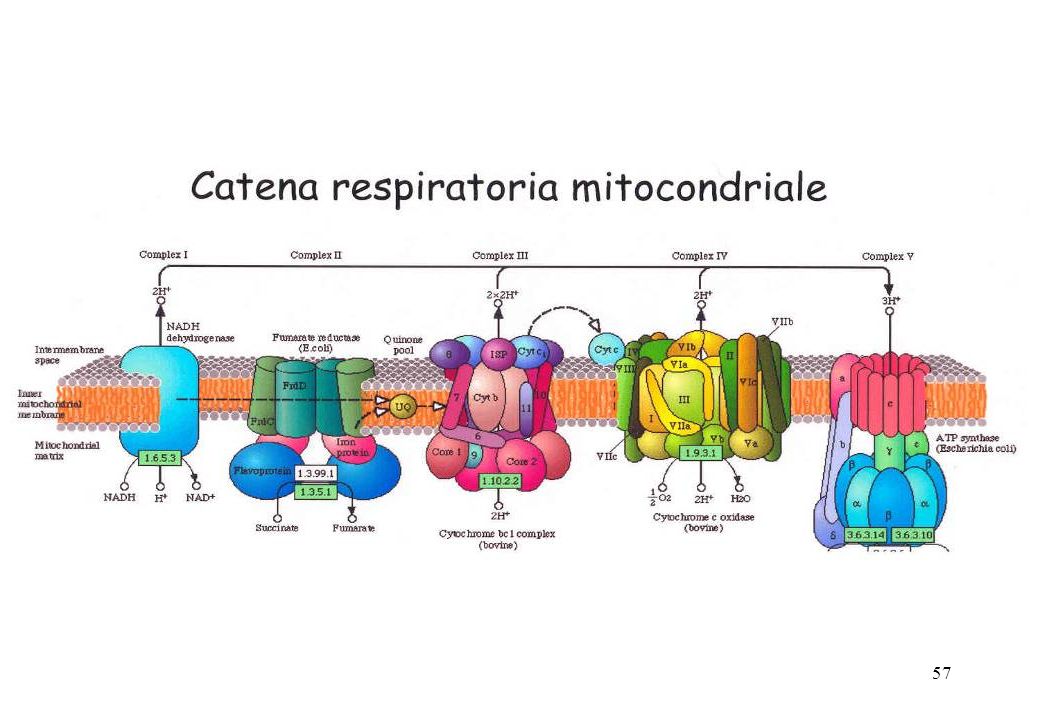
La catena respiratoria comprende 3 complessi proteici inseriti nella membrana mitocondriale interna (complessi I, III e IV) e molecole di trasferimento mobili, quali il coenzima Q e il citocromo-c: complesso I: NADH deidrogenasi complesso III: citocromo-c reduttasi complesso IV: citocromo-c ossidasi II Il complesso II è la succinato deidrogenasi, è l’unico enzima presente del ciclo dell’acido citrico presente nella membrana interna mitocondriale. Il complesso V, l’ATP sintasi, non partecipa al trasferimento di elettroni
 e molecole di trasferimento mobili, quali il coenzima Q e il citocromo-c: complesso I: NADH deidrogenasi. complesso III: citocromo-c reduttasi. complesso IV: citocromo-c ossidasi. II. Il complesso II è la succinato deidrogenasi, è l’unico enzima presente del ciclo dell’acido citrico presente nella membrana interna mitocondriale. Il complesso V, l’ATP sintasi, non partecipa al trasferimento di elettroni.")
26
Tutti i complessi della catena respiratoria sono formati da numerose subunità polipeptidiche e contengono una serie di coenzimi redox legati alle proteine: Flavine: FMN e FAD nei complessi I e II Coenzima Q Centri Ferro-Zolfo: complessi I, II e III Gruppi eme: II, III e IV I trasportatori flavinici e il CoQ sono trasportatori misti: trasportano elettroni e H+ I centri Ferro-Zolfo e i gruppi eme dei citocromi sono trasportatori puri: trasportano elettroni

27
Nicotinamide adenina dinucleotide (NAD+)
e nicotinamide adenina dinucleotide fosfato (NADP+). R= H nel NAD+ R= PO3–- nel NADP
. R= H nel NAD+ R= PO3–- nel NADP.")
28
L’FMN e il FAD subiscono delle reazioni di ossido-riduzione reversibili. L’FMN è saldamente legato al complesso-I della catena respiratoria e non si comporta come un substrato diffusibile. Il FAD è tenacemente legato al complesso II, o succinato deidrogenasi. Le flavine sono dei derivati della vitavina riboflavina

29
Flavina adenina dinucleotide (FAD)
Costituita da FMN + AMP

30
Il CoQ, a differenza dei nucleotidi flavinici è diffusibile e può spostarsi, durante il trasporto elettronico, tra molecole donatrici e accettrici. Il coenzima Q è un benzochinone con una catena isoprenoide insatura. Un valore numerico riportato al pedice indica il numero di unità isoprenoidi (CoQ6, CoQ10)
.")
31
I tre stati di ossidazione dell’FMN e del CoQ
Forma ossidata I tre stati di ossidazione dell’FMN e del CoQ Flavin mononucleotide (FMN) ( forma ossidata o chinonica) CoenzimaQ (CoQ) o ubichinone (forma ossidata o chinonica) Forma radicalica FMNH*(forma radicalica o semichinonica) CoenzimaQH*o ubisemichinone (forma radicalica o semichinonica Forma ridotta Coenzima QH2 o ubichinolo (forma ridotta o idrochinonica) FMNH 2 (forma ridotta o idrochinonica)
 ( forma ossidata o chinonica) CoenzimaQ (CoQ) o ubichinone (forma ossidata o chinonica) Forma radicalica. FMNH*(forma radicalica o semichinonica) CoenzimaQH*o ubisemichinone. (forma radicalica o semichinonica. Forma ridotta. Coenzima QH2 o ubichinolo (forma ridotta o idrochinonica) FMNH 2 (forma ridotta o idrochinonica)")
32
Stati di ossidazione di Flavin mononuleotide (FMN) e Coenzima Q (CoQ) o ubichinone
Il FMN (a) e il coenzimaQ (b) formano radicali liberi semichinonici stabili. IL FMN e il CoQ (che possono trasferire uno o due elettroni per volta) sono quindi il punto di contatto tra il donatore a due elettroni NADH e i citocromi accettori ad un solo elettrone
 e il coenzimaQ (b) formano radicali liberi semichinonici stabili. IL FMN e il CoQ (che possono trasferire uno o due elettroni per volta) sono quindi il punto di contatto tra il donatore a due elettroni NADH e i citocromi accettori ad un solo elettrone.")
33
Esempi di centri ferro-zolfo
I centri Fe-S, come i nucleotidi flavinici, sono strettamente legati alle proteine. Il ferro è legato allo zolfo elementare e ai gruppi tiolici di cisteine messe a disposizione dalla proteina. Benchè in un centro possano essere presenti vari atomi di ferro, i centri ferro-zolfo partecipano a reazioni di trasferimento di un solo elettrone Esempi di centri ferro-zolfo 33

34
I citocromi sono delle proteine contenenti eme.
I citocromi rappresentano l’ultima classe di componenti che partecipano al trasporto degli elettroni. I citocromi sono delle proteine contenenti eme. L’eme è la ferro-porfirina che si trova nelle emoglobine e nella mioglobina. I citocromi del tipo a, b e c sono le principali varianti di queste proteine presenti nelle cellule. Ciascun citocromo è costituito da una catena polipeptidica e da un gruppo eme specifico. Il ferro presente nei citocromi, ma non quello nell’emoglobina, subisce delle ossido-riduzioni fisiologiche passando dallo stato ferroso (2+) a quello ferrico (3+) e viceversa
 a quello ferrico (3+) e viceversa.")
35
I gruppi eme sono composti tetrapirrolici contenenti ferro
Esempi di gruppi eme I gruppi eme sono composti tetrapirrolici contenenti ferro

36
Sequenza del trasporto degli elettroni
L’ossidazione del NADH e del FADH2 viene compiuta dalla catena di trasporto degli elettroni, un insieme di complessi proteici contenenti centri redox caratterizzati da affinità per gli elettroni progressivamente crescenti. Gli elettroni viaggiano lungo questa catena partendo da potenziali di riduzione standard più bassi verso potenziali più alti. Gli elettroni vengono trasportati dai Complessi I e II al Complesso III mediante il CoQ, e dal complesso III al Complesso IV attraverso la proteina periferica di membrana citocromo c 4 H+ Complesso I Complesso II Succinato deidrogenasi-FAD FeS, cit b560

37
NADH + CoQ (ossidato) NAD+ + CoQ (ridotto)
Il complesso I o NADH-coenzima Q ossidoreduttasi (NADH deidrogenasi) catalizza l’ossidazione del NADH da parte del CoQ. E’ il più grosso, complesso contenente 43 catene polipeptidiche. Contiene una molecola di FMN e sei o sette centri ferro-zolfo che partecipano al trasporto degli elettroni. NADH + CoQ (ossidato) NAD+ + CoQ (ridotto) 4 H+ (6-7) Complesso II Succinato deidrogenasi-FAD FeS, cit b560 Complesso I 37
 catalizza l’ossidazione del NADH da parte del CoQ. E’ il più grosso, complesso contenente 43 catene polipeptidiche. Contiene una molecola di FMN e sei o sette centri ferro-zolfo che partecipano al trasporto degli elettroni. NADH + CoQ (ossidato) NAD+ + CoQ (ridotto) 4 H+ (6-7) Complesso II. Succinato deidrogenasi-FAD. FeS, cit b560. Complesso I. 37.")
38
Il NADH può trasferire soltanto due elettroni per volta, mentre i citocromi del Complesso III a cui il CoQ passa i suoi elettroni, sono in grado di accettare solo un elettrone per ogni passaggio. Il FMN e CoQ che possono trasferire uno o due elettroni per volta, sono quindi il punto di contatto tra il donatore a due elettroni NADH e i citocromi accettori a un solo elettrone.

39
Il complesso I potrebbe esistere in due stadi conformazionali:
Modello che rappresenta il trasporto dei protoni (batteriodopsina,una proteina integrale di membrana di Halobacterium halabium) Nel momento in cui gli elettroni vengono trasportati tra i vari centri redox del Complesso I, quattro protoni vengono trasferiti fuori dalla matrice nello spazio inermembrana . Il complesso I potrebbe esistere in due stadi conformazionali: ossidato e ridotto.
 Nel momento in cui gli elettroni vengono trasportati tra i vari centri redox del Complesso I, quattro protoni vengono trasferiti fuori dalla matrice nello spazio inermembrana . Il complesso I potrebbe esistere in due stadi conformazionali: ossidato e ridotto.")
40
FADH2 + CoQ (ossidato) FAD + CoQ (ridotto)
Il complesso II (citocromo c-reduttasi) catalizza l’ossidazione del FADH2 da parte del CoQ FAD + CoQ (ridotto) FADH2 + CoQ (ossidato) I suoi centri redox comprendono il FAD legato covalentemente alla succinato deidrogenasi, sul quale passano inizialmente gli elettroni , un centro 4Fe-4S, due centri 2Fe-2S e un citocromo b560 . 4 H+ Complesso I Complesso II Succinato deidrogenasi-FAD FeS, cit b560
 catalizza l’ossidazione del FADH2 da parte del CoQ. FAD + CoQ (ridotto) FADH2 + CoQ (ossidato) I suoi centri redox comprendono il FAD legato covalentemente alla succinato deidrogenasi, sul quale passano inizialmente gli elettroni , un centro 4Fe-4S, due centri 2Fe-2S e un citocromo b H+ Complesso I. Complesso II. Succinato deidrogenasi-FAD. FeS, cit b560.")
41
Quando il succinato è convertito in fumarato nel ciclo del TCA, nella succinico deidrogenasi avviene una concomitante riduzione del FAD legato a FADH2. Questo FADH2 trasferisce i suoi elettroni immediatamente ai centri Fe-S che li passano al UQ. UQ + 2H+ + 2e = UQH2

42
L’energia libera per il trasferimento degli elettroni dal succinato al CoQ è insufficiente per promuovere la sintesi di ATP . Il complesso II è però ugualmente importante perché consente ad elettroni con un potenziale relativamente alto di entrare nelle catene di trasporto degli elettroni saltando il Complesso I. Il complesso I e II non operano in serie, anche se entrambi trasferiscono elettroni da substrati ridotti, NADH o Succinato al CoQ. Il CoQ che diffonde nel doppio strato lipidico tra i complessi respiratori, serve come una sorta di punto di raccolta per gli elettroni. Dalla prima tappa dell’ossidazione degli acidi grassi si generano gli elettroni che entrano nella catena di trasporto degli elettroni a livello del CoQ Il CoQ raccoglie anche gli elettroni provenienti dal FADH2 prodotto dalla navetta del glicerolfosfato.

43
CoQ (ridotto) + citocromo c (ossidato)
Il complesso III catalizza l’ossidazione del CoQ (ridotto) da parte del citocromo c . La UQ-citocromo c riduttasi (UQ-cit c riduttasi, nome con cui è noto questo complesso) Tale complesso funziona per consentire a una molecola di CoQH2 , un trasportatore a due elettroni , di ridurre due molecole di citocromo c, un trasportatore a un elettrone. Questo complesso contiene due citocromi b,un citocromo c1 e un centro 2Fe-2S (centro di Rieske) al quale è legata la proteina ferro zolfo detta ISP. citocromo c (ridotto) + CoQ (ossidato) CoQ (ridotto) + citocromo c (ossidato) 4 H+ Complesso I Complesso II Succinato deidrogenasi-FAD FeS, cit b560
 da parte del citocromo c . La UQ-citocromo c riduttasi (UQ-cit c riduttasi, nome con cui è noto questo complesso) Tale complesso funziona per consentire a una molecola di CoQH2 , un trasportatore a due elettroni , di ridurre due molecole di citocromo c, un trasportatore a un elettrone. Questo complesso contiene due citocromi b,un citocromo c1 e un centro 2Fe-2S (centro di Rieske) al quale è legata la proteina ferro zolfo detta ISP. citocromo c (ridotto) + CoQ (ossidato) CoQ (ridotto) + citocromo c (ossidato) 4 H+ Complesso I. Complesso II. Succinato deidrogenasi-FAD. FeS, cit b560.")
44
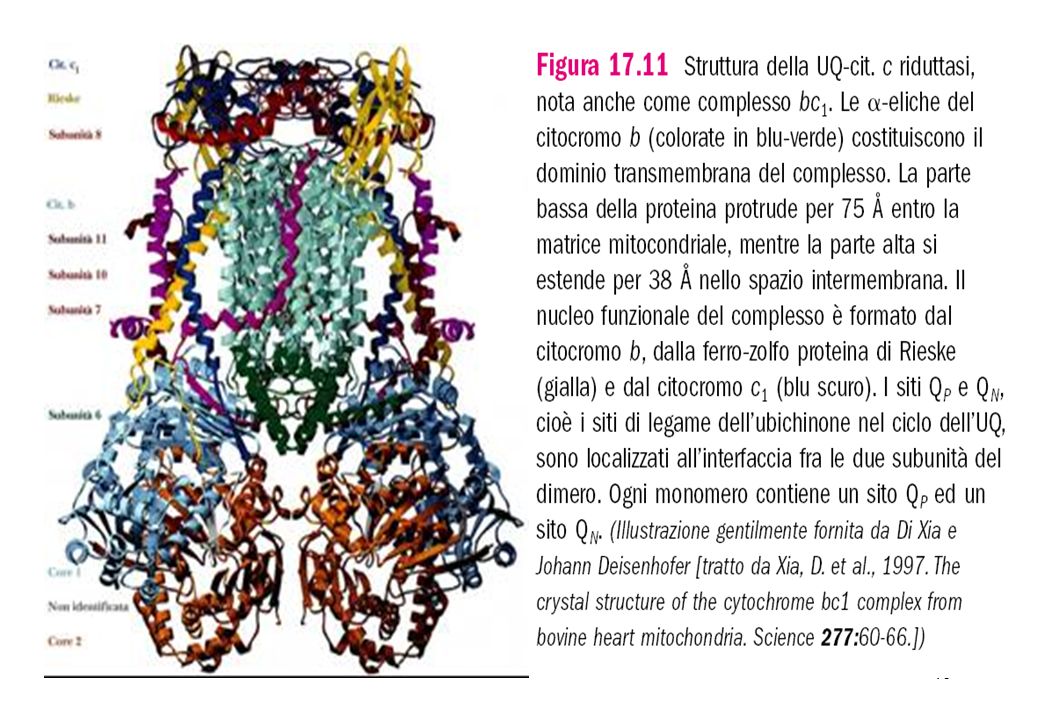
Il Complesso III è un dimero a forma di pera la cui parte più larga è immersa nella matrice mitocondriale. La porzione che attraversa la membrana è costituita da 13 eliche transmembrana per ogni monomero la maggior parte delle quali sono inclinate rispetto al piano della membrana. Otto di queste eliche appartengono alle subunità del citocromo b che lega i gruppi eme di entrambi i citocromi di tipo b, il citocromo b 562 ( detto anche citocromo H a causa del suo alto potenziale e che si trova vicino allo spazio intermembrana ) e b ( chiamato anche b1 per il basso potenziale e che si trova nella matrice ). La proteina ferro-zolfo (ISP) che lega il centro di Rieske, è ancorata nella regione transmembrana mediante due eliche situate alla sua estremità N-terminale e si estende nello spazio intermembrana. Il citicromo c1 è ancorato nella regione transmembrana mediante la sua eleica C-terminale relativamente mobile contenente un gruppo eme di tipo c
 e b 566 ( chiamato anche b1 per il basso potenziale e che si trova nella matrice ). La proteina ferro-zolfo (ISP) che lega il centro di Rieske, è ancorata nella regione transmembrana mediante due eliche situate alla sua estremità N-terminale e si estende nello spazio intermembrana. Il citicromo c1 è ancorato nella regione transmembrana mediante la sua eleica C-terminale relativamente mobile contenente un gruppo eme di tipo c.")
45
Esempi di gruppi eme I gruppi eme sono composti tetrapirrolici contenenti ferro I cit a contengono una catena isoprenoide di 15 carbonii uniti a un gruppo vinilico modificato ed un gruppo formile al posto di uno dei metili. I cit b contengono la ferro-protoporfirina IX lo stesso eme che si trova nell’emoglobina e nella mioglobina . I cit c contengono l’eme c , derivato dalla ferro-protoporfirina IX ma legata covalentemente a residui di cisteina della apoproteina.

47
La membrana mitocondriale interna contiene una grande concentrazione di CoQ e CoQH2
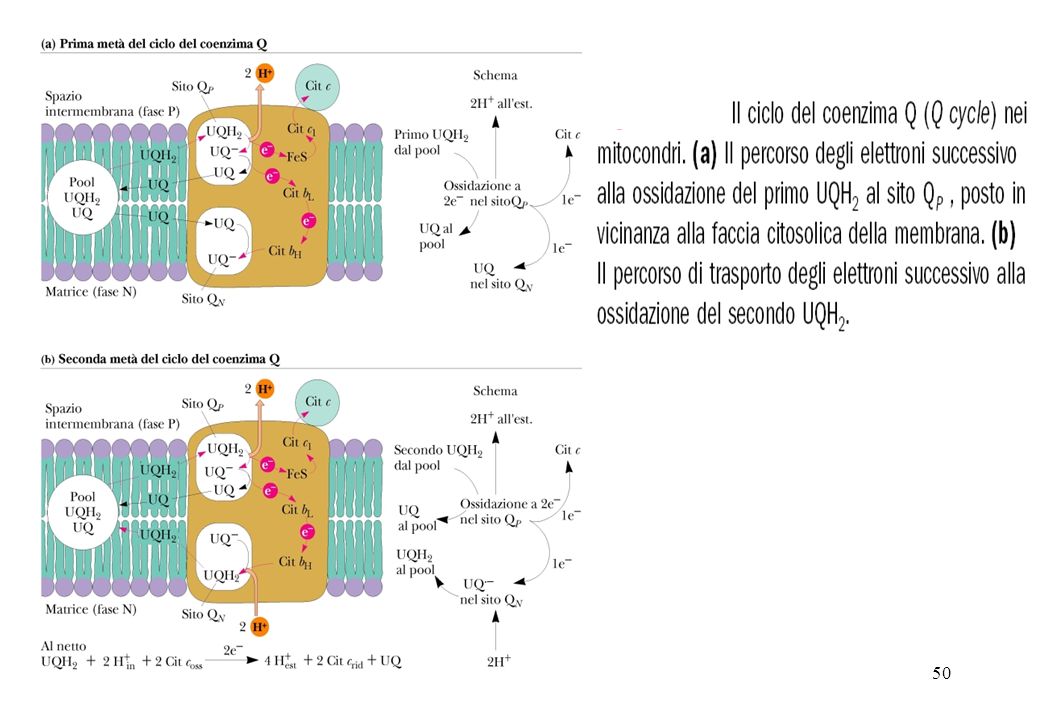
Il ciclo Q inizia quando una molecola di UQH2 diffonde dalla membrana ad un sito catalitico del complesso III il sito QP. L’ossidazione di QUH2 ha luogo in due fasi: Un primo elettrone proveniente dal UQH2 è trasferito alla proteina di Rieske e successivamente al cit c1. Questa reazione rilascia due H+ nel citosol e produce UQ.-, un anione semichinonico del UQ nel sito QP. Un secondo elettrone è successivamente trasferito a l’eme bL convertendo l’ UQ.- a UQ. L’elettrone sull’eme bL vicino al lato citosolico della membrana è quindi trasferito ad una molecola di UQ su un secondo legante chinoni QN convertendo questo UQ a UQ.-. UQ.- rimane saldamente legato al sito QN . Si completa la prima metà del ciclo del CoQ

48
La seconda metà del ciclo è simile alla prima.
Una seconda molecola di UQH2 viene ossidata al sito QP ed en secondo elettrone è trasferito al cit c1 L‘ altro elettrone passa all’eme bL e successivamente all’eme bH. L’elettrone su bH è trasferito sul anione semichinonico UQ.-, al sito QN . Con l’apporto di due H+ provenienti dalla matrice mitocondriale questa reazione produce UQH2, che viene rilasciato dal sito QN e torna nella membrana completando il ciclo del CoQ

49
L’essenza del ciclo Q è che il CoQH2 subisce una riossidazione che avviene in due cicli, in cui il semichinone, CoQ.-, è un intermedio stabile. Questo richiede che vi siano per il coenzima Q due siti di legame indipendenti: QP che si lega a CoQH2 ed è localizzato tra il centro di Rieske [2Fe-2S] e il gruppo eme bL in prossimità dello spazio intermembrana. QN che lega sia CoQ.- sia CoQ ed è localizzato vicino al gruppo eme bH in prossimità della matrice.

51
Il citocromo c è un trasportatore mobile di elettroni
Gli elettroni che attraversano il Complesso III sono trasferiti al citocromo C dal cit c1. Il citocromo C è il solo citocromo nella catena di trasporto degli elettroni che sia idrosolubile. Il citocromo C come l’UQ è un trasportatore mobile , è associato debolmente alla membrana interna mitocondriale ( nello spazio intermembrana dalla parte citosolica della membrana interna) . In questa posizione è in grado di acquisire elettroni dall’insieme Fe-S-cit1 del Complesso III e successivamente migrare lungo la superfice della membrana allo stato ridotto, per trasferire gli elettroni alla citocromo c ossidasi , il IV complesso della catena di trasporto degli elettroni.
 . In questa posizione è in grado di acquisire elettroni dall’insieme Fe-S-cit1 del Complesso III e successivamente migrare lungo la superfice della membrana allo stato ridotto, per trasferire gli elettroni alla citocromo c ossidasi , il IV complesso della catena di trasporto degli elettroni.")
52
IL CITOCROMO C E’ UN MEDIATORE DELLA APOPTOSI morte cellulare programmata
Reagendo a segnali molecolari nel citosol, nella membrana mitocondriale si aprono i canali di trasporto che rilasciano il citocromo c. il citocromo c a sua volta, attiva le caspasi, una famiglia di proteasi contenenti una cisteina nel loro sito catalitico. L’attivazione delle caspasi innesca una serie di reazioni proteolitiche che causano in ultimo la morte della cellula

53
citocromo c (ridotto) + ½ O2
citocromo c (ossidato) + H2O citocromo c (ridotto) + ½ O2 Il complesso IV (citocromo c-ossidasi) catalizza l’ossidazione del citocromo c ridotto da parte dell’O2, l’accettore terminale degli elettroni nel processo di trasporto degli elettroni 4 H+ Complesso I Complesso II Succinato deidrogenasi-FAD FeS, cit b560 citocromo c ossidasi
 + H2O. citocromo c (ridotto) + ½ O2. Il complesso IV (citocromo c-ossidasi) catalizza l’ossidazione del citocromo c ridotto da parte dell’O2, l’accettore terminale degli elettroni nel processo di trasporto degli elettroni. 4 H+ Complesso I. Complesso II. Succinato deidrogenasi-FAD. FeS, cit b560. citocromo c. ossidasi.")
54
Il complesso IV dei mammiferi è un dimero i cui monomeri hanno un peso molecolare di 200 KD e sono costituiti da 13 subunità. La parte centrale del ComplessoIV è costituita dalle sue tre subunità I,II,III, più grandi e più idrfobiche, che sono codificate dal DNA mitocondriale, le altre sono codificate dal DNA nucleare e devono essere trasportate dentro il mit. Struttura del dimero delle citocromo c ossidasi, l’enzima che catalizza le ossidazioni, con l’acquisto di un elettrone, di quattro molecole consecutive di citocromo c ridotto e la contemporanea riduzione di una molecola di O2 utilizzando quattro elettroni. 4 citocromoc (Fe2+) + 4H+ + O citocromo c(Fe3+) + 2H2O
 + 4H+ + O2 4 citocromo c(Fe3+) + 2H2O.")
55
Contiene quattro centri redox: il citocromo a, il citocromo a3, un atomo di rame chiamato CuB e un paio di atomi di rame chiamato centro CuA, Sono anche presenti uno ione Mg2+ e uno ione Zn2+ . Il Fe dell’eme a3 e il CuB formano un unico complesso binucleare

56
RIASSUMENDO Reazione della citocromo c ossidasi e riduzione di O2 ad H2O. Per ridurre una molecola di O2 a 2 di H2O a livello del complesso binucleare citocromo a3-CuB è necessario un totale di 4 elettroni, donati dal citocromo c, e di 4 protoni che si originano nella matrice mitocondriale. Oltre ai 4 protoni utilizzati per ridurre l’O2, 4 protoni provenienti dalla matrice vengono traslocati nello spazio intermembrana (ogni 2 e- che attraversano il complesso IV, vengono traslocati 2 protoni).
.")
58
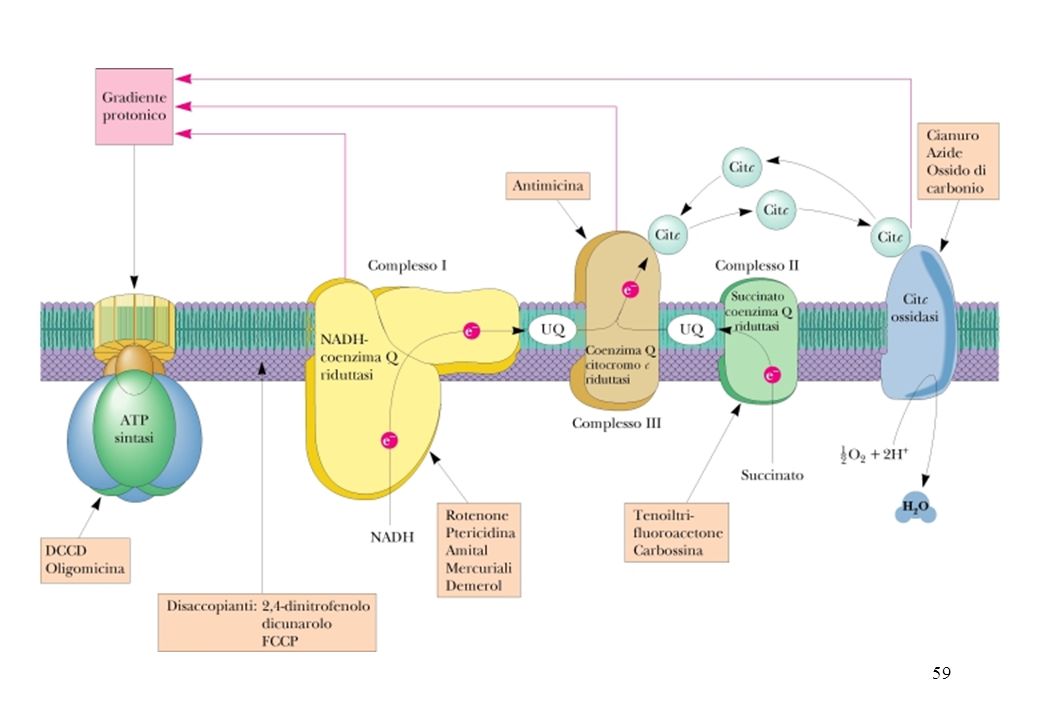
Inibitori della catena respiratoria
Inibitore sito/tipo di azione Rotenone Si lega al complesso I e blocca il trasferimento di elettroni dai gruppi Fe-S all’ubichinone (Q) Carbossina Si lega al complesso II e blocca il trasferimento di elettroni dal FADH2 all’ubichinone Antimicina A Si lega al complesso III e blocca il trasferimento dall’ubichinolo ai gruppi Fe-S Cianuro Blocca il flusso elettronico legandosi al Fe3+ dei citocromi del complesso IV Monossido di carbonio Oligomicina Blocca il flusso di H+ attraverso l’ATP sintasi e Amital
 Carbossina. Si lega al complesso II e blocca il trasferimento di elettroni dal FADH2 all’ubichinone. Antimicina A. Si lega al complesso III e blocca il trasferimento dall’ubichinolo ai gruppi Fe-S. Cianuro. Blocca il flusso elettronico legandosi al Fe3+ dei citocromi del complesso IV. Monossido di carbonio. Oligomicina. Blocca il flusso di H+ attraverso l’ATP sintasi. e Amital.")
60
Inibitori sito-specifici del trasporto di elettroni

61
dalla matrice mitocondriale allo spazio intermembrana
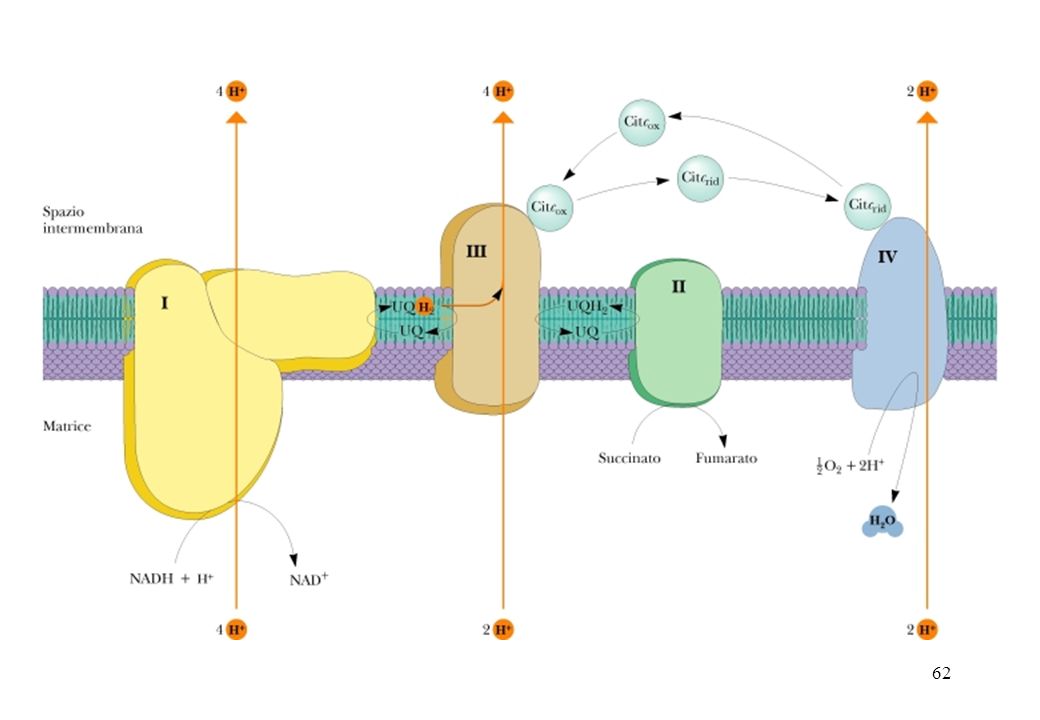
Il trasporto degli elettroni lungo la catena respiratoria determina un passaggio di protoni dalla matrice mitocondriale allo spazio intermembrana Teoria Chemiosmotica di Peter Mitchell L’energia libera prodotta durante il trasporto degli elettroni viene conservata pompando ioni H+ dalla matrice mitocondriale nello spazio intermembrana, per creare attraverso la membrana mitocondriale interna un gradiente elettrochimico di H+. Il potenziale elettrochimico di questo gradiente viene sfruttato per sintetizzare ATP. Il trasporto degli elettroni , promuove il trasferimento dei protoni da parte dei Complessi I, III, e IV dalla matrice una regione contenente una bassa [H+], attraverso la membrana mitocondriale interna, allo spazio intermembrana ( che è in contatto con il citosol), caratterizzato da un’alta [H+].
, caratterizzato da un’alta [H+].")
63
Nel modello proposto da Mitchell, il pH della matrice si innalza, e la matrice stessa acquista un potenziale elettrico negativo rispetto al citosol come conseguenza dell’uscita dei protoni, Il pompaggio dei protoni cioè produce un gradiente di pH ed una differenza di potenziale elettrico attraverso la membrana mitocondriale interna, condizioni che contribuiscono entrambe ad attrarre i protoni di nuovo nella matrice del citoplasma. Il flusso di protoni lungo questo gradiente elettrochimico è un processo favorito energeticamente, e può quindi promuovere la sintesi di ATP.

64
Fosforilazione ossidativa
l’energia libera prodotta durante il trasporto degli e- deve essere conservata in una forma utilizzabile dall’ATP sintasi.

65
Il trasporto degli elettroni , promuove il trasferimento dei protoni da parte del complesso I, III, e IV dalla matrice una regione contenente una bassa concent. di H+, attraverso la membrana mitocondriale interna, caratterizzata da un’alta concent. di H+ L’energia libera sequestrata dal risultante gradiente elettrochimico (forza motrice protonica ) alimenta la sintesi di ATP.
 alimenta la sintesi di ATP.")
66
L’ATP sintasi (F1Fo-ATPasi), è una proteina transmembrana costituita da più subunità con un peso complessivo di 450 KD. Fo ( lega l’antibiotico oligomicina B) è costituita nei mammiferi da 6 copie di una proteina che attraversa la membrana che formano un canale polare per il passaggio di H+. F1 è costituita da subunità a3b3gde. Le subunità a e b sono disposte in modo alternato. Lo stelo è costituito dalla subunità g associato alle subunità d ed e
 è costituita nei mammiferi da 6 copie di una proteina che attraversa la membrana che formano un canale polare per il passaggio di H+. F1 è costituita da subunità a3b3gde. Le subunità a e b sono disposte in modo alternato. Lo stelo è costituito dalla subunità g associato alle subunità d ed e.")
67
Il meccanismo per la sintesi di ATP può essere suddiviso in tre tappe:
Traslocazione di protoni promossa da F0 Formazione catalitica del legame fosfoanidrinico dell’ATP promossa da F1 Accoppiamento della dissipazione del gradiente protonico con la sintesi di ATP, che necessitano dell’interazione di F1 con F0

68
Interconversione dei tre stati conformazionali
1 - L'ADP e il Pi si legano debolmente al sito di legame nella conformazione L. 2. Un cambiamento conformazionale promosso dal rilascio di energia libera converte il sito L in sito di legame T (chiamato bTP) che catalizza la formazione di ATP. Questa tappa implica anche cambiamenti conformazionali negli altri due protomeri, che convertono il sito T a cui è legato l'ATP in sito aperto (O) (detto bE), e che convertono il sito O in sito L. 3. L'ATP viene sintetizzato a livello del sito T di una subunità mentre si dissocia dal sito O di un'altra subunità. L'energia libera fornita dal flusso dei protoni facilita principalmente il rilascio dall'enzima dell'ATP appena sintetizzato; cioè, promuove la transizione T O, distruggendo così le interazioni ATP-enzima che in precedenza avevano promosso la formazione spontanea di ATP a partire da ADP e Pi, quando la subunità era nella conformazione T.
 che catalizza la formazione di ATP. Questa tappa implica anche cambiamenti conformazionali negli altri due protomeri, che convertono il sito T a cui è legato l ATP in sito aperto (O) (detto bE), e che convertono il sito O in sito L. 3. L ATP viene sintetizzato a livello del sito T di una subunità mentre si dissocia dal sito O di un altra subunità. L energia libera fornita dal flusso dei protoni facilita principalmente il rilascio dall enzima dell ATP appena sintetizzato; cioè, promuove la transizione T O, distruggendo così le interazioni ATP-enzima che in precedenza avevano promosso la formazione spontanea di ATP a partire da ADP e Pi, quando la subunità era nella conformazione T.")
69
IL RAPPORTO P/O E possibile esprimere la quantità di ATP sintetizzato in termini di molecole di substrato ossidate. Le ossidazioni di NADH e FADH2 sono associati rispettivamente con la sintesi di 3, 2 molecole di ATP. Questa stechiometria chiamata rapporto P/O mette in relazione la quantità di ATP sintetizzata con la quantità di ossigeno che viene ridotto.

70
Disaccoppiamento della fosforilazione ossidativa
La presenza nella membrana interna di un agente che ne aumenta la permeabilità agli ioni H+ disaccoppia la fosforilazione ossidativa dal trasporto degli elettroni in quanto fornisce una strada per la dissipazione del gradiente protonico elettrochimico che non necessita della sintesi di ATP. La dissipazione di un gradiente elettrochimico di H+, che viene generato dal trasporto degli elettroni ed è disaccoppiato dalla sintesi di ATP, produce calore .

71
Meccanismo d’azione del 2,4-dinitrofenolo
Uno ionoforo trasportatore di protoni, come il DNP, disaccoppia la fosforilazione ossidativa dalla catena di trasporto degli elettroni dissipando il gradiente elettrochimico di protoni generato dal trasporto degli elettroni.

72
Nel tessuto adiposo bruno ( grasso bruno ) il disaccoppiamento della fosforilazione ossidativa genera calore. I mitocondri del grasso bruno contengono un canale per i protoni conosciuto con il nome di proteina disaccippiante ( UCP, chiamata Termogenina ). Il flusso di protoni porta alla dissipazione del gradiente protonico presente attraverso la membrana mitocondriale interna. Questo processo consente all’ossidazione del substrato di procedere e di generare calore senza sintesi diATP:
. Il flusso di protoni porta alla dissipazione del gradiente protonico presente attraverso la membrana mitocondriale interna. Questo processo consente all’ossidazione del substrato di procedere e di generare calore senza sintesi diATP:")
73
Controllo coordinato della glicolisi e del ciclo dell’acido citrico
Le fonti principali degli elettroni che entrano nella catena di trasporto degli elettroni sono: Glicolisi Degradazioni degli ac. grassi Ciclo dell’ac. citrico Disegno schematico ,la sfera verde indica attivazione, la sfera rossa indica inibizione

74
Specie reattive dell’ossigeno
Nella riduzione di O2 si cela un pericolo: il trasferimento di 4 elettroni dà origine a prodotti innocui il trasferimento di un singolo elettrone forma un anione superossido il trasferimento di 2 elettroni genera un perossido O2 e- O2 - . O2 2-

75
I derivati tossici dell’ossigeno molecolare (ROS) vengono rimossi da enzimi protettivi
. 2 + 2H+ O2 + H2O2 superossido dismutasi Catalasi H2O + O2 . 2 2H2 O2 2 GSH+H2O GSSG+ 2 H2O glutatione perossidasi Della superossido dismutasi (SOD) esistono due forma: La SOD mitocondriale un tetramero contenente Mn La SOD citosolica un dimero contenente Cu e Zn
 esistono due forma: La SOD mitocondriale un tetramero contenente Mn. La SOD citosolica un dimero contenente Cu e Zn.")
76
La malattia di Alzheimer
Alcune malattie degenerative sono associate a danni ossidativi del mitocondrio: Morbo di Parkinson La malattia di Alzheimer La Corea di Huntington

77
Alcuni individui affetti dalla malattia ereditaria sclerosi laterale amiotrofica (ALS; malattia di Lou Gehrig) sono caratterizzati da una Cu,Zn-SOD mutata Negli enzimi la magior parte delle mutazioni porta a una perdita della funzione. Nella ALS il difetto viene ereditato con un tratto dominante, fattore che è compatibile con l’acquisizione di una attività tossica. Infatti la SOD si comporta come una perossidasi e si pensa che ossidi i lipidi, causando la degenerazione dei motoneuroni, caratteristica della malattia.

78
Anemia da carenza di Ferro
Fabbisogno di ferro in un uomo adulto: 1mg/die in una donna in età fertile: 2mg/die in una donna in gravidanza: 3mg/die Il ferro è necessario per il mantenimento di una quantità normale di Hb, di citocromi di centri ferro-zolfo.

79
Miopatie mitocondriali
il lattato si accumula nel liquido cerebrospinale crampi, debolezza muscolare, encefalopatia trattamento con CoQ10 Carenza di CoQ10: forma rara che colpisce i bambini.

80
Malattie mitocondriali
Neuropatia ottica ereditaria di Leber (mutazione del Complesso I) Miopatie mitocondriali I mitocondri svolgono un ruolo centrale nell’apoptosi
 Miopatie mitocondriali. I mitocondri svolgono un ruolo centrale nell’apoptosi.")
81
CITOCROMO P450 Sono enzimi ossidativi detossificanti
Idrolizzano le molecole idrofobiche in modo da ottenere prodotti solubili Molti dei substrati sono composti tossici come alcuni idrocarburi aromatici policiclici alcuni dei quali cancerogeni: bifenili policlorurati(PCB), fenobarbital e steroidi L’idrossilazione catalizzata dal cit. 450 converte queste molecole in sostanze più solubili per la successiva escrezione
, fenobarbital e steroidi. L’idrossilazione catalizzata dal cit. 450 converte queste molecole in sostanze più solubili per la successiva escrezione.")
Presentazioni simili