Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
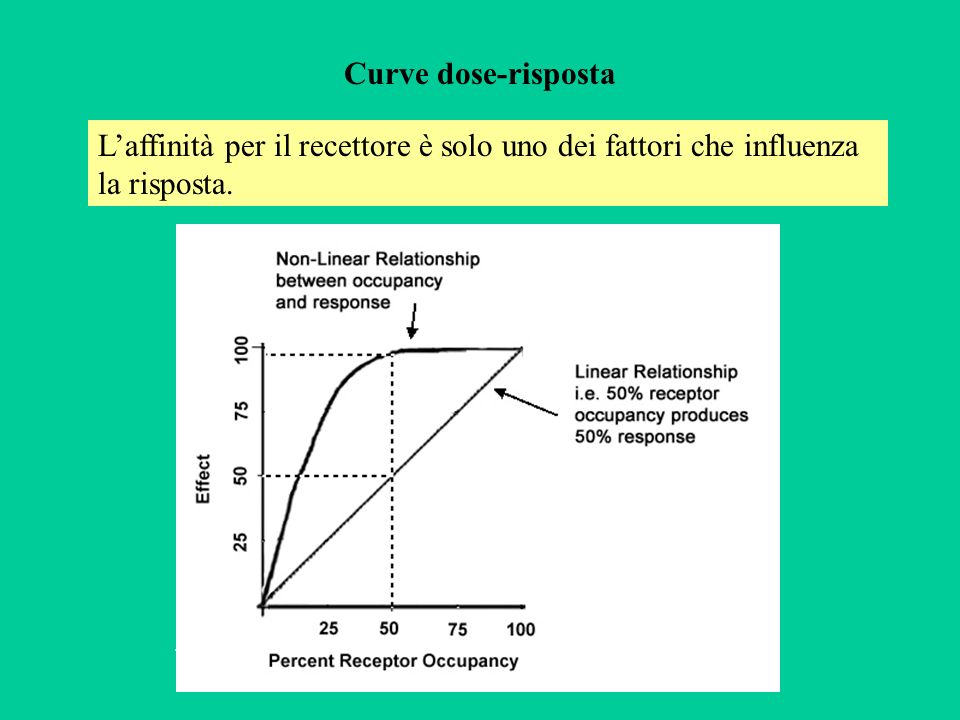
Curve dose-risposta L’affinità per il recettore è solo uno dei fattori che influenza la risposta. .
2
Per caratterizzare l’attività, la potenza e la specificità di un farmaco, si studia la risposta a diversi livelli: Studi in vitro: Livello molecolare soprattutto per inibitori enzimatici Livello cellulare per farmaci che agiscono su recettori di membrana e nucleari, canali ionici, trasportatori Livello di tessuto od organo isolati Studi in vivo: organismo in toto

3
Curve dose-risposta: agonisti
Gli studi in vivo sono poco utilizzati per la caratterizzazione di base dei farmaci per l’interferenza di fattori farmacocinetici e perché la risposta viene influenzata dai meccanismi omeostatici dell’organismo. Si utilizzano quindi in genere sistemi in vitro. Farmaci in grado di evocare essi stessi una risposta (agonisti). Il campione biologico (cellule, tessuto isolato ecc.) viene esposto a concentrazioni crescenti di farmaco. Si misura la risposta con metodi idonei.
. Il campione biologico (cellule, tessuto isolato ecc.) viene esposto a concentrazioni crescenti di farmaco. Si misura la risposta con metodi idonei.")
4
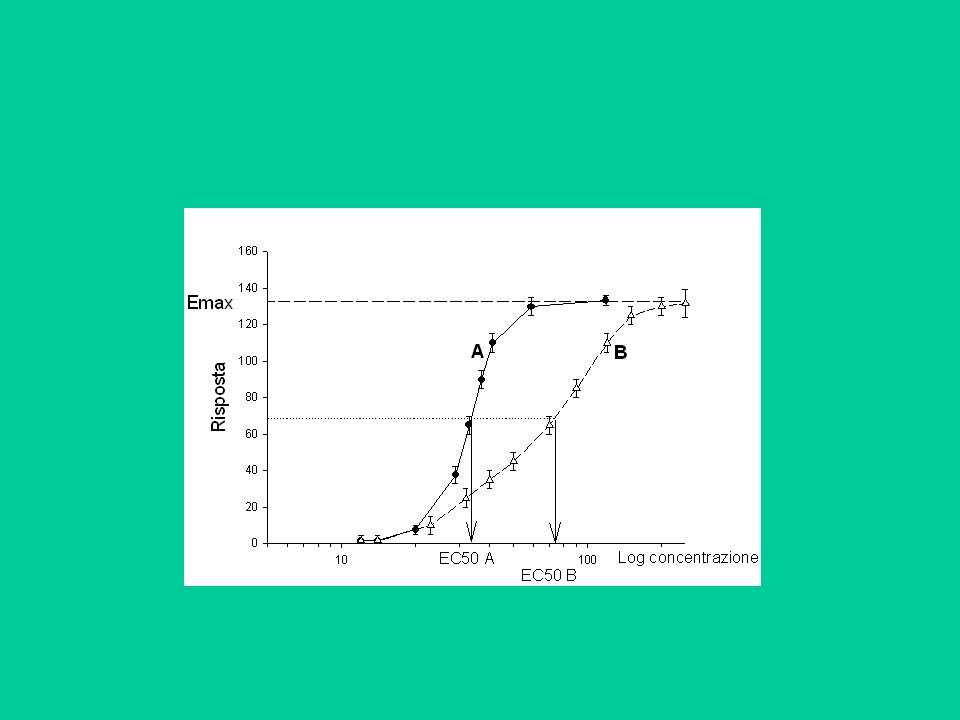
Riportando in un grafico cartesiano le risposte (in ascissa) in funzione del logaritmo della concentrazione (in ordinata), si ottiene in genere una curva sigmoide. I parametri caratterizzanti sono: l’effetto massimo (Emax); la concentrazione (o dose) che produce un effetto pari al 50% di Emax (EC50) e la minima concentrazione alla quale si ha un effetto, che sono una misura della potenza del farmaco. Emax EC50 Log [F] Concentrazione di soglia
; la concentrazione (o dose) che produce un effetto pari al 50% di Emax (EC50) e la minima concentrazione alla quale si ha un effetto, che sono una misura della potenza del farmaco. Emax. EC50. Log [F] Concentrazione di soglia.")
5
L’Emax dipende dal sistema biologico (numero di recettori, amplificazione del segnale ecc.); è quindi uguale per farmaci di una stessa classe che siano agonisti puri ed è una misura dell’efficacia. L’EC50 dipende dal farmaco ed è una misura della potenza.

7
Risposta % EC50 Concentrazione Curve dose-risposta di farmaci con uguale EC50 ma diversa concentrazione-soglia.

8
Curve dose-risposta e farmaci antagonisti
Gli antagonisti non sono in grado di evocare una risposta in seguito alla loro interazione con il recettore. In presenza di un antagonista, la curva dose-risposta di un agonista si sposta ‘a destra’; cioè, per ottenere la stessa risposta sono necessarie concentrazioni più alte. Log [agonista]

9
La comunicazione intercellulare: trasmettitori e recettori
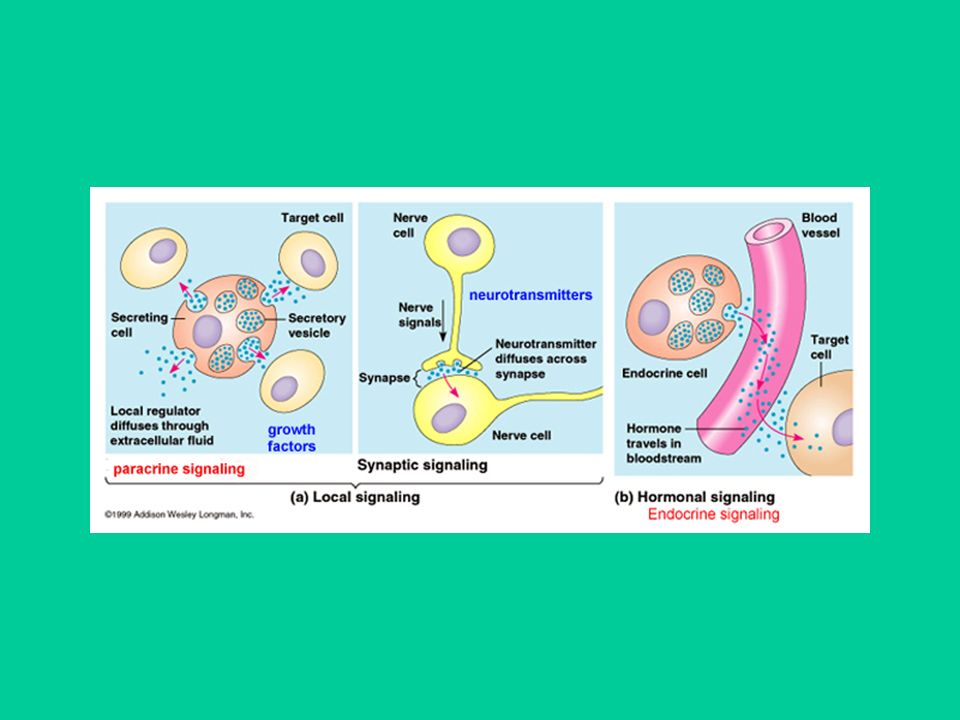
La comunicazione tra cellule permette agli organismi di adattarsi in modo coordinato e continuo alle variazioni ‘ambientali’. Le cellule comunicano mediante mediatori chimici di natura molto varia Il mediatore viene prodotto e secreto da un tipo cellulare La cellula cui è destinata la comunicazione, cellula ‘bersaglio’, ha recettori in grado di riconoscere il mediatore

10
I recettori sono sempre proteine che legano i trasmettitori (ligandi) ad un sito specifico (sito di legame). In conseguenza all’interazione recettore-ligando il recettore modifica la sua conformazione; questa modifica di conformazione da’ origine ad una risposta da parte della cellula bersaglio (trasduzione). Vi sono numerosi meccanismi che provvedono allo ‘spegnimento’ del segnale (interruzione della trasmissione)
. Vi sono numerosi meccanismi che provvedono allo ‘spegnimento’ del segnale (interruzione della trasmissione)")
12
Caratteristiche della trasmissione
Ogni recettore riconosce un solo trasmettitore ma può legare anche dei ‘modulatori’ della trasmissione Un trasmettitore può essere riconosciuto da un solo recettore (es., ormoni) o da più sottotipi recettoriali (es., neurotrasmettitori). La correttezza (specificità) del flusso di comunicazioni è assicurata principalmente da tre fattori: Presenza dei recettori solo sulle cellule bersaglio Nel caso di trasmettitori con recettori multipli, la specificità è assicurata dalla vicinanza della cellula bersaglio al sito di rilascio del trasmettitore Meccanismi di rimozione del trasmettitore
 o da più sottotipi recettoriali (es., neurotrasmettitori). La correttezza (specificità) del flusso di comunicazioni è assicurata principalmente da tre fattori: Presenza dei recettori solo sulle cellule bersaglio. Nel caso di trasmettitori con recettori multipli, la specificità è assicurata dalla vicinanza della cellula bersaglio al sito di rilascio del trasmettitore. Meccanismi di rimozione del trasmettitore.")
13
Tipi di recettore Esistono due grandi classi di recettori
Recettori intracellulari. Recettori di membrana Le due classi differiscono per la natura dei ligandi e per il tipo di risposta cellulare evocato

14
Recettori intracellulari
Sono i recettori per gli ormoni steroidei, tiroidei, per le vitamine A e D. Sono grandi proteine monomeriche che in assenza di ligando si trovano in forma inattiva nel citosol o nel nucleo, legate ad altre proteine ‘inibitorie’ (Heat Shock Proteins, HSP). Il legame con il ligando endogeno (o con un farmaco) provoca la dissociazione dalle HSP e la dimerizzazione delle proteine; il dimero si lega a regioni specifiche del DNA (HRE, Hormone Responsive Elements) e stimola o inibisce la trascrizione di specifici geni.
. Il legame con il ligando endogeno (o con un farmaco) provoca la dissociazione dalle HSP e la dimerizzazione delle proteine; il dimero si lega a regioni specifiche del DNA (HRE, Hormone Responsive Elements) e stimola o inibisce la trascrizione di specifici geni.")
15
Struttura dei recettori intracellulari.
I diversi recettori intracellulari compongono una ‘superfamiglia’; hanno cioè una struttura molecolare molto simile. Inoltre, alcune regioni sono altamente conservate. Dominio di legame al DNA Dominio di legame al ligando Dominio per l’attivazione trascrizionale

17
Caratteristiche della trasmissione mediata dai recettori intracellulari
Sono modulatori della sintesi proteica. L’effetto si instaura quindi lentamente (alcune ore) e perdura a lungo. Specificità: nonostante il dominio di legame con il DNA sia molto simile, ligandi (ormoni) diversi provocano effetti diversi. I meccanismi che assicurano la specificità sono diversi: localizzazione cellulare selettiva; metabolismo tessuto-specifico.
 e perdura a lungo. Specificità: nonostante il dominio di legame con il DNA sia molto simile, ligandi (ormoni) diversi provocano effetti diversi. I meccanismi che assicurano la specificità sono diversi: localizzazione cellulare selettiva; metabolismo tessuto-specifico.")
18
Recettori di membrana Esistono 3 famiglie principali di recettori di membrana: I recettori-canale I recettori accoppiati a proteine G Recettori con attività tirosin-chinasica Le 3 famigli differiscono per struttura, cinetiche e modalità di attivazione. I neurotrasmettitori riconoscono in genere sia recettori-canale che recettori accoppiati a proteine G (es. recettori nicotinici e muscarinici).
.")
19
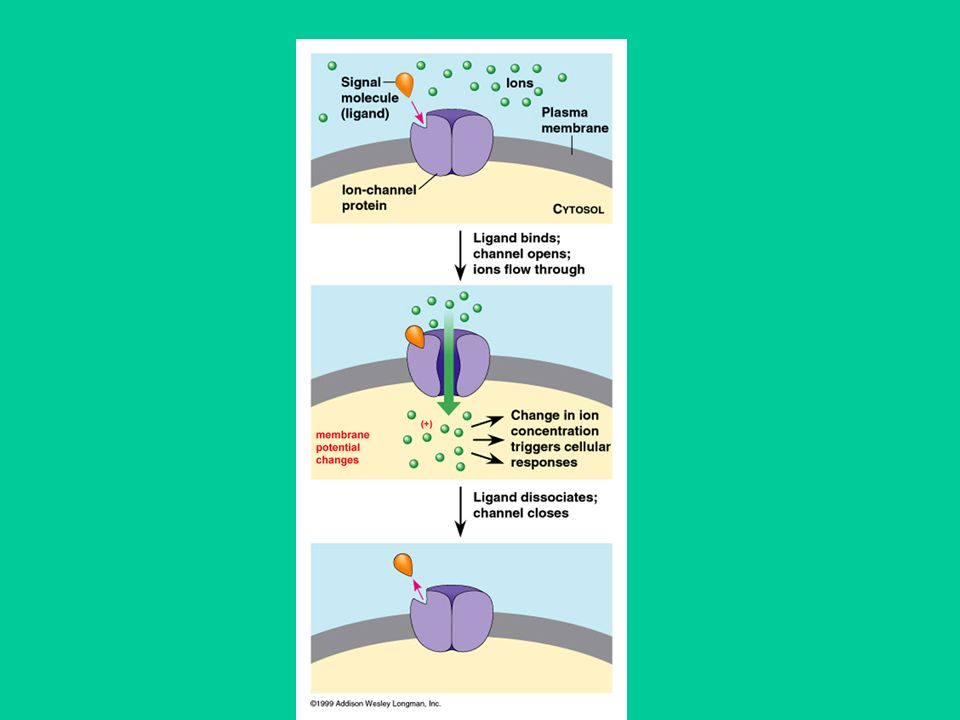
Recettori-canale (recettori ionotropici)
Sono proteine che formano un canale ionico che attraversa la membrana cellulare. Attraverso questi canali possono passare ioni (Na+, K+, Ca++, Cl-) che sono impermeabili alla membrana cellulare. L’attivazione di questi recettori da parte di un agonista endogeno o esogeno determina l’apertura del canale e un conseguente flusso di ioni.
 che sono impermeabili alla membrana cellulare. L’attivazione di questi recettori da parte di un agonista endogeno o esogeno determina l’apertura del canale e un conseguente flusso di ioni.")
21
Il flusso di ioni causa variazioni del potenziale elettrico transmembrana e quindi una variazione dell’eccitabilità cellulare. I recettori-canale sono costituiti da 5 subunità. Il sito di legame per l’agonista si trova in genere all’esterno della membrana cellulare. Sono presenti siti allosterici cui si legano sostanze in grado di modulare la risposta all’agonista. Nella regione intracellulare vi sono siti di fosforilazione che regolano l’attività di questi recettori.

22
L’attivazione continua dei recettori-canale provoca desensitizzazione
Alcune proprietà quali la conduttanza, tempo di apertura del canale, sensibilità alla desensitizzazione, e capacità di legare agonisti e antagonisti varia notevolmente in funzione del tipo di subunità che compone il recettore-canale. Ad esempio, i recettori nicotinici gangliari hanno diversa sensibilità farmacologiche dei recettori nicotinici muscolari.

23
Recettori accoppiati a Proteine G
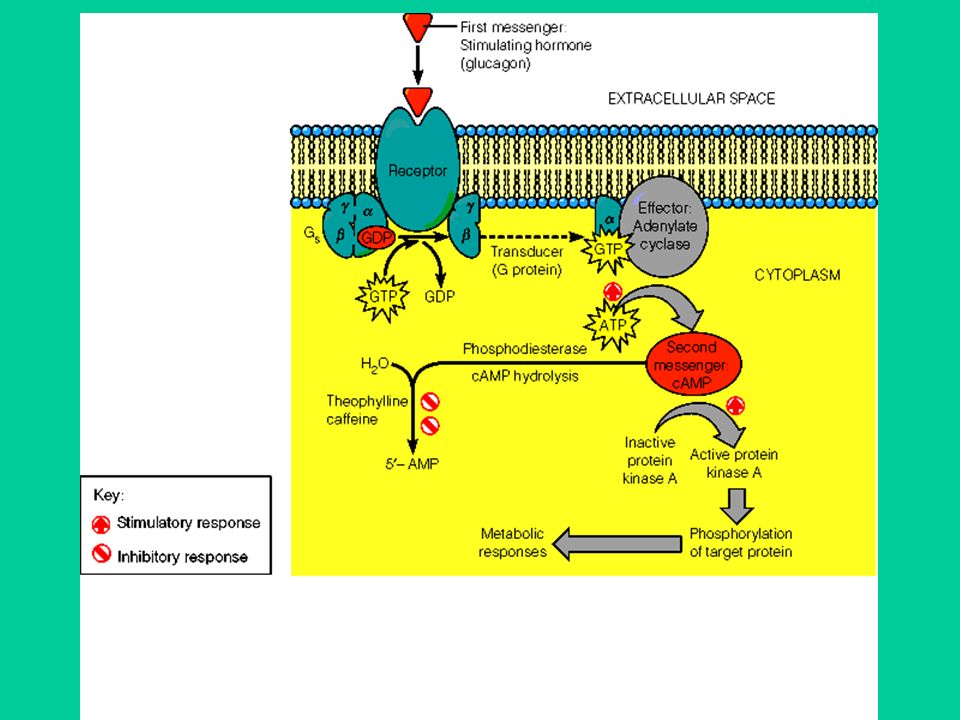
Il legame dell’agonista al recettore provoca una modificazione conformazionale del recettore attivazione della proteina G accoppiata al recettore generazione di un secondo messaggero

24
Struttura dei recettori accoppiati a Proteine G
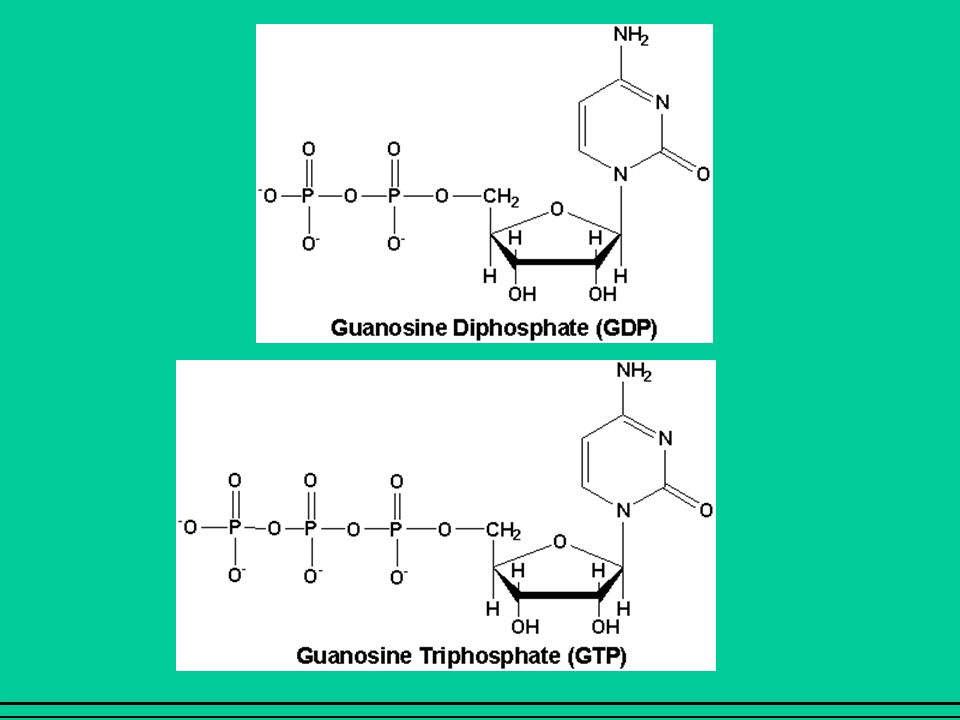
I recettori sono proteine monomeriche il cui filamento proteico attraversa 7 volte la membrana cellulare, formando 3 anse intracellulari e 3 extracellulari. La III ansa intracellulare lega la proteina G Le proteine G sono formate da 3 subunità: , ß, In assenza di agonista la subunità lega il GDP (guanosin difosfato); questo legame conferisce alta affinità per le subunità ß e
; questo legame conferisce alta affinità per le subunità ß e ")
25
Il legame dell’agonista al recettore provoca una modificazione conformazionale del recettore e, di conseguenza, della subunità GDP viene sostituito da GTP (guanosin trifosfato) dissociazione della subunità (con GTP legato) dalle subunità ß. La subunità regola l’attività degli effettori: enzimi (adenilato ciclasi, fosfolipasi C) che catalizzano la formazione di secondi messaggeri; canali ionici (Ca++, K+). Anche la subunità ß può regolare l’attività di alcuni effettori (canali ionici). La subunità è in grado di idrolizzare il GTP ad essa legato (attività GTP-asica); quando il GTP viene idrolizzato a GDP, la subunità si riassocia alla subunità ß spegnimento del segnale.
 che catalizzano la formazione di secondi messaggeri; canali ionici (Ca++, K+). Anche la subunità ß può regolare l’attività di alcuni effettori (canali ionici). La subunità è in grado di idrolizzare il GTP ad essa legato (attività GTP-asica); quando il GTP viene idrolizzato a GDP, la subunità si riassocia alla subunità ß spegnimento del segnale.")
27
Il ciclo delle proteine G

29
L’effetto dell’attivazione delle proteine G dipende dal tipo di subunità
La stimolazione di recettori diversi ma comunque accoppiati a proteine G provoca risposte cellulari diverse. Ad esempio, i recettori ß-adrenergici e quelli colinergici muscarinici sono entrambi accoppiati a proteine G ma producono effetti opposti. La diversità di effetti è dovuta alla diversità delle subunità delle proteine G. Esistono tre tipi di subunità : s, i, q; le proteine G contenenti queste subunità si chiamano Gs, Gi, Gq.

30
I tre tipi di subunità hanno azioni diverse:
s stimola l’adenilato ciclasi i inibisce l’adenilato ciclasi e regola canali ionici q stimola la fosfolipasi C

31
Gli effettori del sistema di trasduzione delle proteine G. 1
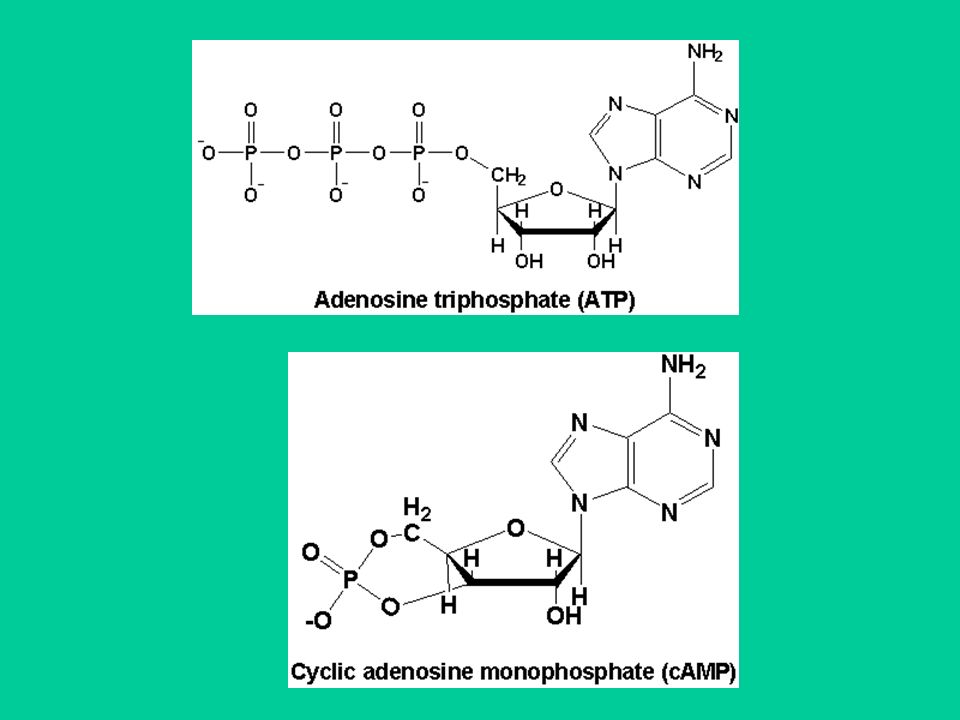
Gli effettori del sistema di trasduzione delle proteine G. 1. Il sistema adenilato ciclasi/cAMP L’adenilato ciclasi è l’enzima che sintetizza il cAMP (3’,5’,-adenosina monofosfato ciclico; AMP ciclico) dall’ATP. Il cAMP regola molte funzioni cellulari, emdiante un unico meccanismo che è l’attivazione di protein chinasi. Le protein chinasi catalizzano la fosforilazione di proteine cellulari, utilizzando ATP come donatore di gruppi fosfato. La fosforilazione delle proteine (enzimi, canali ionici ecc.) è una modalità molto utilizzata per regolare la funzionalità delle proteine. cAMP viene idrolizzato da enzimi detti fosfodiesterasi.
 dall’ATP. Il cAMP regola molte funzioni cellulari, emdiante un unico meccanismo che è l’attivazione di protein chinasi. Le protein chinasi catalizzano la fosforilazione di proteine cellulari, utilizzando ATP come donatore di gruppi fosfato. La fosforilazione delle proteine (enzimi, canali ionici ecc.) è una modalità molto utilizzata per regolare la funzionalità delle proteine. cAMP viene idrolizzato da enzimi detti fosfodiesterasi.")
34
Subunità catalitica Subunità regolatoria

35
Gli effettori del sistema di trasduzione delle proteine G. 2. Il sistema Fosfolipasi C/diacilglicerolo-inositolo fosfato

36
Gli effettori del sistema di trasduzione delle proteine G. 2
Gli effettori del sistema di trasduzione delle proteine G. 2. Il sistema Fosfolipasi C/diacilglicerolo-inositolo fosfato La subunità q stimola la fosfolipasi C La fosfolipasi C (PLC) è un enzima che scinde un fosfolipide di membrana, il fosfatidilinositolo bifosfato (PIP2) in diacilglicerolo (DAG) e inositolo trifosfato (InsP3). DAG e InsP3 sono entrambi secondi messaggeri. DAG attiva la protein chinasi C (PKC); la PKC attivata fosforila diverse proteine intracellulari InsP3 si lega ad un recettore sulla membrana del reticolo endoplasmatico attivazione di un canale Ca++ rilascio di Ca++ dai siti di deposito
 è un enzima che scinde un fosfolipide di membrana, il fosfatidilinositolo bifosfato (PIP2) in diacilglicerolo (DAG) e inositolo trifosfato (InsP3). DAG e InsP3 sono entrambi secondi messaggeri. DAG attiva la protein chinasi C (PKC); la PKC attivata fosforila diverse proteine intracellulari. InsP3 si lega ad un recettore sulla membrana del reticolo endoplasmatico attivazione di un canale Ca++ rilascio di Ca++ dai siti di deposito.")
37
La fosforilazione di proteine (enzimi, canali ionici) è il modo più frequente di regolazione rapida dell’attività delle proteine stesse. Le variazioni della concentrazione intracellulare di Ca++ regola molte risposte cellulari (attivazione di enzimi, di canali ionici, contrazione muscolare, secrezione di ormoni e neurotrasmettitori ecc.) Spegnimento del segnale: DAG viene fosforilato ad acido fosfatidico. InsP3 viene defosforilato a Ins. Ins ed acido fosfatidico formano il fosfatidilinositolo (PI) che poi viene fosforilato a riformare PIP2.
 Spegnimento del segnale: DAG viene fosforilato ad acido fosfatidico. InsP3 viene defosforilato a Ins. Ins ed acido fosfatidico formano il fosfatidilinositolo (PI) che poi viene fosforilato a riformare PIP2.")
Presentazioni simili