PEPTIDI E PROTEINE
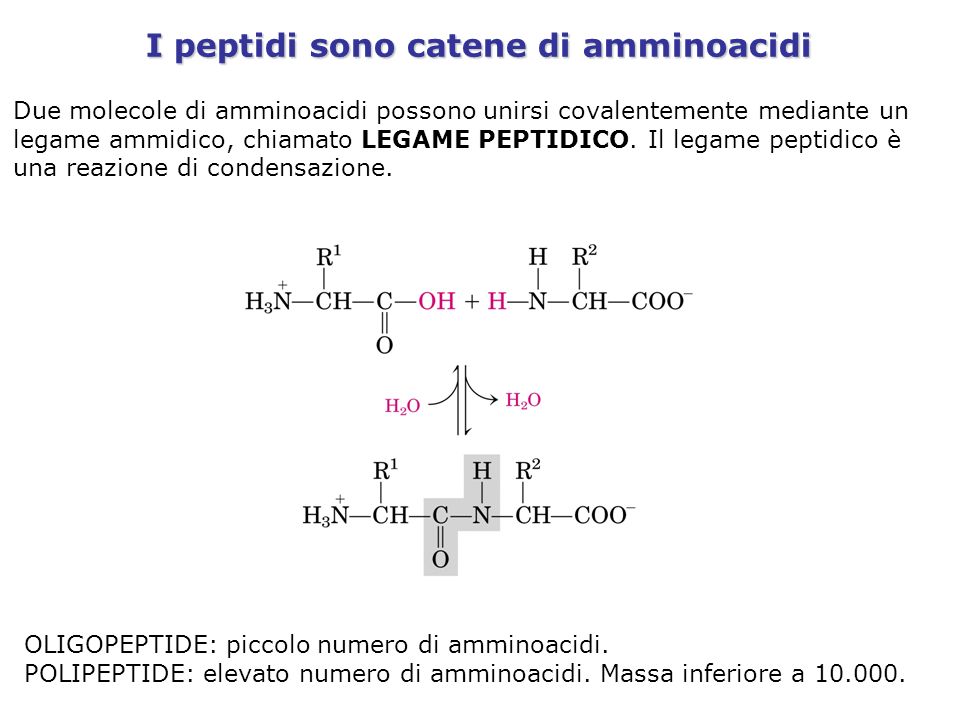
I peptidi sono catene di amminoacidi Due molecole di amminoacidi possono unirsi covalentemente mediante un legame ammidico, chiamato LEGAME PEPTIDICO. Il legame peptidico è una reazione di condensazione. OLIGOPEPTIDE: piccolo numero di amminoacidi. POLIPEPTIDE: elevato numero di amminoacidi. Massa inferiore a 10.000.
Il legame peptidico è rigido e planare Gli atomi di Ca di amminoacidi adiacenti sono separati da tre legami covalenti: Ca – C – N – Ca PROPRIETA’ DEL LEGAME PEPTIDICO I 6 atomi del gruppo peptidico giacciono sullo stesso piano l’ossigeno legato al carbonio del gruppo carbonilico e l’atomo di idrogeno legato all’azoto amminico, si trovano in trans. L’ossigeno carbonilico ha una parziale carica negativa e l’azoto amminico ha una parziale carica positiva ciò genera un parziale dipolo elettrico. I legami ammidici C-N hanno un parziale carattere di doppio legame per effetto della risonanza non possono ruotare liberamente. La rotazione è permessa solo attorno ai legami N-Ca e Ca-C.
Il legame peptidico è rigido e planare e sono di 180° quando il polipeptide è nella conformazione complanare estesa e tutti i gruppi peptidici sono sullo stesso piano. e possono assumere tutti i valori compresi tra -180° e +180°, ma molti valori risultano proibiti per interferenze steriche tra gli atomi dello scheletro del polipeptide e quelli delle catene laterali.
Il legame peptidico è rigido e planare e sono pari a 0 quando i due legami peptidici che fiancheggiano il Ca sono sullo stesso piano CONFORMAZIONE PROIBITA I legami peptidici rigidi limitano il numero di conformazioni che una catena polipeptidica può assumere
Il grafico di Ramachandran BLU: conformazioni in cui non si hanno sovrapposizioni steriche CONFORMAZIONI CONSENTITE. TURCHESI: conformazioni che si trovano al limite della stabilità a causa di contatti non favorevoli tra atomi CONFORMAZIONI CONSENTITE. CELESTI: conformazioni che sono consentite solo se è permessa una certa flessibilità degli angoli di legame CONFORMAZIONI CONSENTITE. L-alanina
I peptidi sono catene di amminoacidi Le unità amminoacidiche sono definite RESIDUI ogni residuo è un amminoacido che ha perso un protone dal suo gruppo amminico e un ossidrile dal suo gruppo carbossilico. Estremità ammino-terminale carbossi-terminale L’idrolisi del legame peptidico, nonostante sia una reazione esoergonica, avviene molto lentamente, a causa dell’elevata energia di attivazione. I legami peptidici delle proteine sono abbastanza stabili (la vita media è di circa 7 anni)
I peptidi e i polipeptidi biologicamente attivi hanno dimensioni molto variabili Non è possibile fare generalizzazioni sulla massa molecolare dei peptidi e delle proteine attive in relazione alla loro funzione. Molti peptidi di piccole dimensioni svolgono la loro funzione a concentrazioni molto basse (ossitocina, bradichidina, ecc.). Peptidi: ormone pancreatico, glucagone.
I peptidi e i polipeptidi biologicamente attivi hanno dimensioni molto variabili Alcune proteine sono costituite da una singola catena polipeptidica, altre da 2 o più polipeptidi associati noncovalentemente (MULTISUBUNITA’). Se almeno 2 catene sono uguali, si parla di PROTEINA OLIGOMERICA, e le 2 unità identiche sono chiamate PROTOMERI. EMOGLOBINA: DIMERO DI PROTOMERI O TETRAMERO CON 4 SUBUNITA’ POLIPEPTIDICHE.
I polipeptidi hanno una caratteristica composizione di amminoacidi Quando una proteina viene completamente idrolizzata, produce una miscela di amminoacidi.
Alcune proteine contengono gruppi chimici diversi dagli amminoacidi Molti enzimi contengono solo amminoacidi e nessun altro gruppo chimico PROTEINE SEMPLICI. Altre proteine contengono, oltre agli amminoacidi, gruppi chimici funzionali permanentemente associati PROTEINE CONIUGATE. La parte non amminoacidica viene definita GRUPPO PROSTETICO.
Le proteine hanno diversi livelli di struttura Insieme dei legami covalenti (legami peptidici e ponti disolfuro) che uniscono le subunità monomeriche di una proteina. Si riferisce a disposizioni particolarmente stabili dei residui amminoacidici che danno origine a organizzazioni strutturali ricorrenti, Descrive tutti gli aspetti del ripiegamento tridimensionale di un polipeptide. Descrive la disposizione nello spazio di due o più subunità polipeptidiche.
ENERGIA LIBERA: G, è la quantità di lavoro macroscopico che il sistema può compiere sull'ambiente ENTALPIA: H, è una funzione di stato che esprime la quantità di energia che un sistema termodinamico può scambiare con l'ambiente. ENTROPIA: l'entropia è una grandezza che viene interpretata come una misura del disordine di un sistema fisico.
PROCESSO SPONTANEO (G<0) G = H - TS dove G: forza trainante H: variazione di entalpia creata dalla scissione o dalla formazione di legami S: variazione del disordine Poiché H è positivo in un processo di fusione o di evaporazione, e poiché in questi due processi la S è elevata e positiva (aumenta il disordine), ne consegue che il G è negativo PROCESSO SPONTANEO (G<0)
La struttura tridimensionale delle proteine Lo scheletro covalente delle proteine è costituito da numerosi legami. Se la rotazione attorno a questi legami fosse libera, le proteine potrebbero assumere un numero infinito di strutture. Ogni proteina ha una specifica funzione chimica e strutturale Ogni proteina assume un’unica struttura tridimensionale
La struttura tridimensionale delle proteine Cinque principi: La struttura tridimensionale di una proteina è determinata dalla sua sequenza amminoacidica. La funzione di una proteina dipende dalla sua struttura. Una determinata proteina assume un’unica o poche conformazioni stabili. Le forze più importanti che stabilizzano la struttura di una determinata proteina sono interazioni non covalenti. Tra le numerose strutture delle diverse proteine, si possono individuare alcuni modelli strutturali comuni. LE PROTEINE PERO‘ NON HANNO STRUTTURE STATICHE LA LORO FUNZIONE IMPLICA IL PASSAGGIO DA UNA CONFORMAZIONE ALL’ALTRA.
La struttura tridimensionale delle proteine L’organizzazione spaziale degli atomi di una proteina viene detta CONFORMAZIONE. Le conformazioni possibili di una proteina comprendono tutti gli stati strutturali che è possibile ottenere senza la rottura dei legami covalenti. Un cambiamento conformazionale può avvenire, ad esempio, a causa della rotazione attorno a legami singoli. Soltanto una, o poche, delle innumerevoli conformazioni teoricamente possibili di una proteina contenente centinaia di legami singoli è quella che tende a predominare nelle condizioni biologiche. La necessità dell’esistenza di numerose conformazioni stabili è legata alle modificazioni che avvengono nella maggior parte delle proteine in seguito al legame con altre molecole o mentre catalizzano reazioni chimiche. Le conformazioni possibili nelle specifiche condizioni sono quelle termodinamicamente più stabili, che hanno la minore energia libera G. Le proteine che si trovano nella loro conformazione funzionale vengono dette PROTEINE NATIVE.
La conformazione di una proteina è stabilizzata da interazioni deboli STABILITÀ: tendenza a mantenere la conformazione nativa. Le proteine native sono solo marginalmente stabili la differenza di energia libera (G) tra stati avvolti e non avvolti è solo tra i 20 e i 65 jK/mole. Una data catena polipeptidica può teoricamente assumere innumerevoli conformazioni poiché lo stato non avvolto possiede un’elevata entropia conformazionale. L’entropia conformazionale e le interazioni generate dai legami idrogeno tra la proteina e le molecole di solvente tendono a mantenere lo stato non avvolto.
La conformazione di una proteina è stabilizzata da interazioni deboli La stabilità di una proteina non è data semplicemente dalla somma delle energie libere di formazione delle numerose interazioni deboli presenti. Ogni gruppo capace di formare un legame idrogeno era impegnato con una molecola di acqua, prima che la proteina si ripiegasse. Per ogni legame idrogeno che si genera all’interno di una proteina, bisogna che si rompa un legame idrogeno (della stessa forza) tra lo stesso gruppo e l’acqua. LA STABILITA’ NETTA DETERMINATA DA UN’INTERAZIONE DEBOLE (differenza di energia libera tra stato avvolto e non avvolto) E’ PROSSIMA ALLO ZERO.
Perché la conformazione nativa di una proteina è favorita rispetto alla altre? 1. I gruppi idrofobici tendono a strutturarsi in organizzazioni compatte diminuzione dello strato di solvatazione dell’acqua AUMENTO DI ENTROPIA. 2. Quando un composto polare viene posto in acqua, si hanno due effetti: una diminuzione netta dei legami idrogeno per unità di massa; si forma uno strato di solvatazione delle molecole di acqua strutturate attorno alla molecola polare. 3. Quando una proteina assume la sua conformazione nativa, la formazione di legami intramolecolari porta alla riduzione dello strato di solvatazione AUMENTO DI ENTROPIA. La maggior parte della variazione di energia libera che si ha quando in una proteina si formano interazioni deboli deriva dall’aumento di entropia della soluzione acquosa circostante, per effetto dell’ammassamento delle superfici idrofobiche circostanti.
Perché la conformazione nativa di una proteina è favorita rispetto alla altre? 4. All’interno di una proteina nella sua conformazione nativa, la presenza di gruppi polari o carichi che non abbiano una controparte idonea con cui formare legami idrogeno o interazioni ioniche, determina una CONDIZIONE FORTEMENTE DESTABILIZZANTE E TERMODINAMICAMENTE INSOSTENIBILE LO STATO RIPIEGATO NON E’ PIU’ FAVORITO. 5. I legami idrogeno tra gruppi presenti nelle proteine si generano attraverso un meccanismo cooperativo. Inoltre, le interazioni tra gruppi con cariche opposte (ponti salini), possono avere effetti stabilizzanti su uno o più stati conformazionali nativi di alcune proteine.