Scaricare la presentazione
1
LA MATRICE EXTRACELLULARE

2
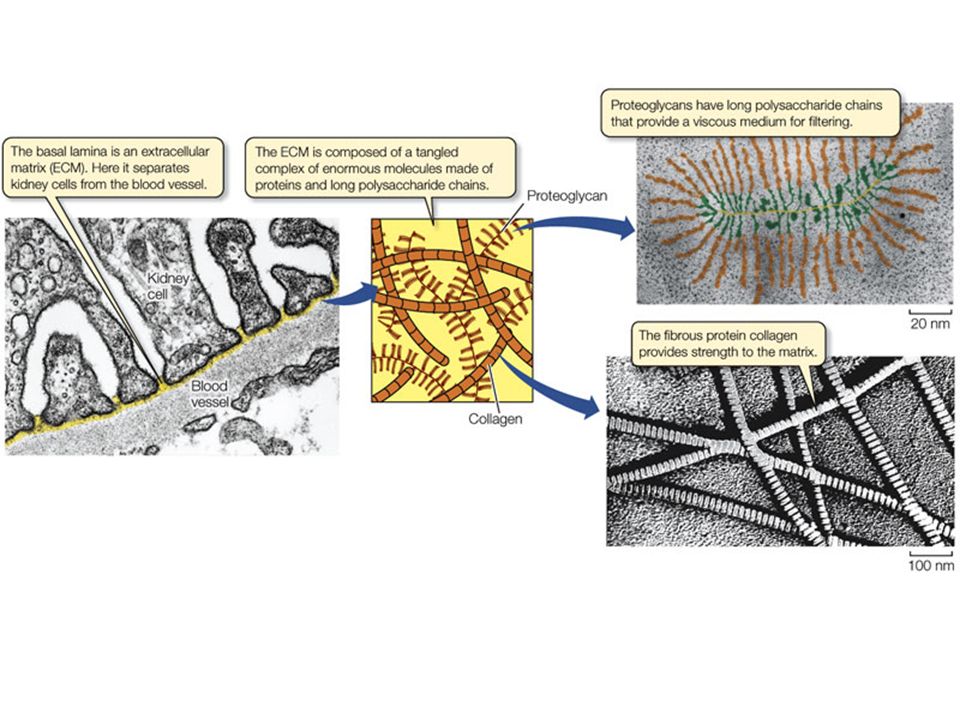
La matrice extracellulare (MEC) è una rete formata da molecole secrete dalle cellule animali
Glicosaminoglicani e Proteoglicani Proteine fibrose (Collagene, Elastina) Proteine “di collegamento” multiadesive (Fibronectina, Laminina, ecc.) Nel corpo umano alcuni tessuti hanno poca MEC (es. cervello), altri ne possiedono in grande quantità (es. osso, cartilagine).
 Proteine di collegamento multiadesive (Fibronectina, Laminina, ecc.) Nel corpo umano alcuni tessuti hanno poca MEC (es. cervello), altri ne possiedono in grande quantità (es. osso, cartilagine).")
3
Diffusione controllata di soluti, comprese molecole di segnalazione
Grazie alle proprietà chimico-fisiche delle molecole che costituiscono la matrice extracellulare, in particolare i glicosaminoglicani e i proteoglicani, si forma una sorta di gel che richiama acqua, contribuendo ad alcune proprietà importanti della MEC: Resistenza dei tessuti alla compressione Diffusione controllata di soluti, comprese molecole di segnalazione Migrazione cellulare (durante lo sviluppo embrionale e durante i processi di riparazione nei tessuti adulti)
")
5
I proteoglicani sono costituiti da diverse catene di glicosaminoglicani legati a una proteina centrale; i proteoglicani possono a loro volta attaccarsi ad un enorme polisaccaride (acido ialuronico). proteoglicano

6
Strutture dei Glicosaminoglicani
Eteropolisaccaridi lineari formati da ripetizioni di un disaccaride ialuronidasi

7
Proteoglicani della matrice extracellulare
Rispetto alle glicoproteine, nei proteoglicani la porzione glucidica è quella predominante (fino al 95%) Lo ialuronano resiste alla compressione e facilita la migrazione cellulare
 Lo ialuronano resiste alla compressione e facilita la migrazione cellulare.")
8
Proteoglicani della superficie cellulare
Molti fattori di crescita sono sequestrati e presentati alle cellule dai proteoglicani Proteoglicani della superficie cellulare

9
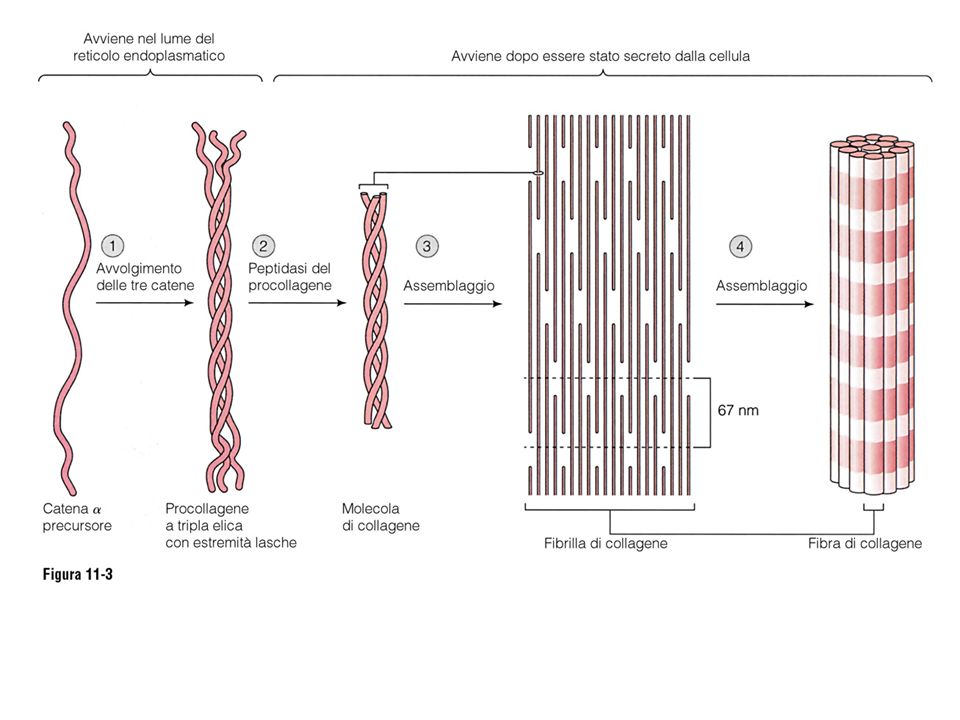
Nella matrice extracellulare, i proteoglicani e i glicosaminoglicani si legano a proteine fibrose (collagene, elastina); ci sono poi alcune proteine di adesione (fibronectina, laminina) che connettono le cellule alla matrice. Collagene/i: le proteine fibrose più abbondanti della matrice extracellulare

10
Le catene polipeptidiche dei collageni sono caratterizzate dalla presenza di una Glicina ogni tre amminoacidi, e da numerose Lisine e Proline, molte delle quali idrossilate (legame agli zuccheri glicoproteina)
")
11
L’unità strutturale di base dei collageni è una tripla elica formata da diversi assortimenti di catene polipeptidiche di tipo 1 e 2

14
Le collagenopatie Esistono numerose patologie da alterazioni del collagene, che possono essere distinte in tre gruppi: metaboliche, come lo scorbuto, causato da avitaminosi C: infatti l’acido ascorbico (o vitamina C) è un cofattore delle idrossilasi che modificano Pro e Lys del collagene, modificazioni estremamente importanti per la corretta formazione delle triple eliche di collagene; b) degenerative, per cause varie (autoimmuni, invecchiamento, ecc) nella MEC di alcuni organi viene prodotta una quantità eccessiva di collagene che sostituisce o comunque modifica le caratteristiche del tessuto funzionale; c) genetiche: mutazioni con conseguenze patologiche (nei casi meno gravi, anomala “elasticità” articolare, o fragilità ossea): alcune comportano alterazioni nella struttura del collagene incompatibili con la sopravvivenza (forme di osteogenesi imperfetta)
 è un cofattore delle idrossilasi che modificano Pro e Lys del collagene, modificazioni estremamente importanti per la corretta formazione delle triple eliche di collagene; b) degenerative, per cause varie (autoimmuni, invecchiamento, ecc) nella MEC di alcuni organi viene prodotta una quantità eccessiva di collagene che sostituisce o comunque modifica le caratteristiche del tessuto funzionale; c) genetiche: mutazioni con conseguenze patologiche (nei casi meno gravi, anomala elasticità articolare, o fragilità ossea): alcune comportano alterazioni nella struttura del collagene incompatibili con la sopravvivenza (forme di osteogenesi imperfetta)")
15
Un altro tipo di proteina fibrosa: l’elastina

16
Le fibronectine legano diverse cellule ai collageni fibrosi e ad altri componenti della matrice extracellulare tramite le integrine (a loro volta collegate al citoscheletro)
")
17
Struttura della laminina

18
La laminina e il collagene di tipo IV formano la lamina basale a sviluppo bidimensionale, situata tra le cellule epiteliali ed il tessuto connettivo sottostante.

19
ADESIONE e CONTATTI CELLULA/CELLULA E CELLULA/MATRICE

20
Le CAM (Cell-Adhesion Molecules) comprendono cinque classi principali di proteine
 comprendono cinque classi principali di proteine")
21
Integrine I recettori presenti sulla membrana cellulare che “contattano” fibronectina, laminina e tenascina (proteine della MEC) appartengono alla famiglia delle integrine. Le integrine sono in grado di interagire anche con proteine di membrana (sempre interazione eterofilica)
 appartengono alla famiglia delle integrine. Le integrine sono in grado di interagire anche con proteine di membrana (sempre interazione eterofilica)")
22
Le integrine sono eterodimeri
Le integrine sono eterodimeri. Ogni eterodimero è composto di una subunità a e una subunità b. Entrambe le catene sono caratterizzate da un importante dominio extracellulare, una zona idrofobica transmembrana e da una corta coda citoplasmatica. Nei vertebrati esistono 16 tipi di catena ed 8 tipi di catena che generano più di 20 integrine distinte.

23
Legame tra Fibronectina ed Integrina
I domini extracellulari dell’integrina legano i componenti della ECM attraverso un legame tra la subunità b e il motivo RGD (Arg-Gly-Asp).
.")
24
Le integrine mediano interazioni deboli cellula-matrice e cellula-cellula
L’adesione cellula-matrice è modulata da variazioni dell’attività e del numero di integrine Fattori de-adesivi promuovono la migrazione cellulare

25
L’aggregazione delle piastrine nel sangue è mediata da particolari tipi di integrine presenti sulla loro membrana plasmatica L’integrina IIb3, normalmente presente sulla membrana plasmatica delle piastrine, non è in grado di legarsi alla proteine ematica fibrinogeno né ad altri ligandi proteici che partecipano tutti alla formazione del coagulo ematico.

26
Dopo che la piastrina viene “attivata” dal legame al collagene della membrana basale del vaso leso o alla trombina nel coagulo in formazione, l’integrina è in grado di legarsi al fibrinogeno; questa interazione accelera la formazione del coagulo. L’attivazione piastrinica è accompagnata da un’alterazione conformazionale nell’integrina IIb3. I pazienti con difetti genetici alla subunità 3 dell’integrina sono predisposti ad eccessivo sanguinamento, comprovando il ruolo dell’integrina nella formazione dei coaguli sanguigni.

27
Selectine Esistono tre tipi di selectine: L-selectina, E-selectina e P-selectina. Strutturalmente la proteina è composta da un dominio N-terminale omologo alle lectine animali Ca2+-dipendenti, da un dominio tipo EGF, da 2 a 9 domini seguiti da un tratto elicoidale transmembrana e da una corta coda citoplasmatica.

28
Ruolo delle lectine nel riconoscimento cellula-cellula
Le lectine sono una famiglia di proteine che sono altamente specifiche per determinati zuccheri. Svolgono un importante ruolo biologico nel processo di riconoscimento delle proteine cellulari.

29
Selectine Le selectine mediano il processo di adesione cellulare tramite il riconoscimento, Ca2+-dipendente, dei glicani sialiati.

30
Superfamiglia delle Immunoglobuline
La superfamiglia delle immunoglobuline comprende gli anticorpi e un’ampia varietà di proteine transmembrana; alcune di queste ultime mediano l’adesione cellula-cellula Ca2+-indipendente (interazione omofilica). Strutturalmente queste proteine sono composte da una serie di domini simili.
. Strutturalmente queste proteine sono composte da una serie di domini simili.")
31
Caderine Famiglia composta da una dozzina di proteine che mediano l’adesione cellulare omofilica Ca2+-dipendente (uniscono cellule dello stesso tipo). Ne esistono più tipi: E-caderina (epiteliale), N-caderina (neurale) e P-caderina (placentale).
. Ne esistono più tipi: E-caderina (epiteliale), N-caderina (neurale) e P-caderina (placentale).")
32
Le caderine presentano:
un segmento extracellulare esteso composto da una serie di subunità poste una dopo l'altra per formare una fibra; vi sono ioni Ca2+ legati ad ogni subunità per dare rigidità alla struttura (se Ca2+ viene rimosso, la catena diventa molle e viene facilmente distrutta dagli enzimi proteolitici) un breve tratto transmembrana un dominio citoplasmatico
 un breve tratto transmembrana. un dominio citoplasmatico.")
33
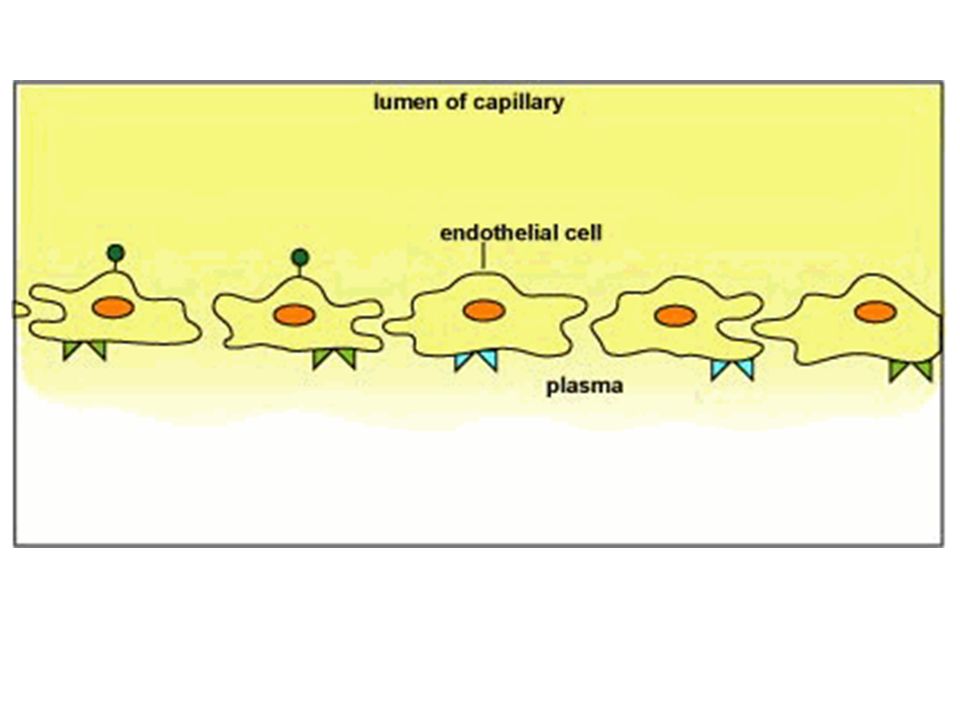
Chemiotassi e molecole di adesione
La chemiotassi è un fenomeno complesso che viene definito come la capacità delle cellule del circolo sanguigno di migrare dal sangue alla sede di un processo infiammatorio in atto o in un sito di infezione. Il reclutamento dei leucociti nel sito di infezione avviene attraverso meccanismi che richiedono l’intervento di numerose molecole. Le molecole di adesione sono presenti sulla superficie delle cellule endoteliali, dei leucociti e nella matrice tissutale. 33

34
4. DIAPEDESI in cui il leucocita attraversa la parete endoteliale
Il processo che porta alla fuoriuscita dei leucociti dai vasi è chiamato EXTRAVASAZIONE e si verifica in 4 fasi. 1. Legame reversibile del leucocita all’epitelio vascolare (ROTOLAMENTO) 2. Stadio di ARRESTO dove il leucocita è legato all’endotelio in modo irreversibile 3. Stadio di ADESIONE STABILE in cui il leucocita è saldamente ancorato e il rotolamento è bloccato 4. DIAPEDESI in cui il leucocita attraversa la parete endoteliale 34
 2. Stadio di ARRESTO dove il leucocita è legato all’endotelio in modo irreversibile. 3. Stadio di ADESIONE STABILE in cui il leucocita è saldamente ancorato e il rotolamento è bloccato. 4. DIAPEDESI in cui il leucocita attraversa la parete endoteliale. 34.")
35
Diverse fasi dell’adesione del leucocita alla parete del vaso
arresto rotolamento adesione stabile diapedesi lume vasale tessuto endotelio 35

37
STRUTTURE COMPLESSE di ADESIONE e COMUNICAZIONE CELLULA-CELLULA e CELLULA-MATRICE

38
Meccanismi di adesione (giunzionale e non) che legano le cellule tra loro (soprattutto negli epiteli) o alla matrice extracellulare
 che legano le cellule tra loro (soprattutto negli epiteli) o alla matrice extracellulare")
39
Le giunzioni: permettono il contatto e la comunicazione cellula-cellula
Giunzioni strette (o occludenti) Desmosomi (o di ancoraggio) Giunzioni comunicanti Plasmodesmi (giunzioni comunicanti nei tessuti vegetali)
 Desmosomi (o di ancoraggio) Giunzioni comunicanti. Plasmodesmi (giunzioni comunicanti nei tessuti vegetali)")
40
Giunzioni strette: creano dei “sigilli” tra cellule animali adiacenti
Giunzioni strette: creano dei “sigilli” tra cellule animali adiacenti. Bloccano il passaggio nello spazio intercellulare. Desmosomi: ancorano tra loro le cellule animali nei tessuti molto resistenti. Giunzioni comunicanti: mettono in connessione cellule animali adiacenti, sono formate da proteine “tunnel”.

41
Giunzioni strette (Tight Junctions)
Presenti apicalmente nel dominio laterale

42
Barriera impermeabile tra le cellule
No sigillo continuo, ma serie di fusioni focali (“trapunta”)
")
43
Le giunzioni strette sigillano alcuni organi cavi (es
Le giunzioni strette sigillano alcuni organi cavi (es. stomaco, intestino) e limitano la diffusione delle proteine e dei fosfolipidi di membrana
 e limitano la diffusione delle proteine e dei fosfolipidi di membrana.")
44
Le giunzioni occludenti garantiscono il movimento direzionale dei materiali

45
Resistenza alle forze di trazione
Desmosomi Resistenza alle forze di trazione

46
Struttura molecolare di un desmosoma
Placche tenute assieme da caderine Ca2+-dipendenti (se si allontana il Ca2+ le placche si separano e le cellule si distaccano)
")
47
Accoppiamento metabolico
Le giunzioni comunicanti (gap junction) permettono a ioni e piccole molecole (<2000 Da) di diffondere tra cellule adiacenti Accoppiamento metabolico
 permettono a ioni e piccole molecole (<2000 Da) di diffondere tra cellule adiacenti. Accoppiamento metabolico.")
48
6 molecole di connessina
Le connessine sono proteine transmembrana che formano “canali” cilindrici (connessoni) raggruppati nelle giunzioni comunicanti 6 molecole di connessina
 raggruppati nelle giunzioni comunicanti. 6 molecole di connessina.")
49
Es. chiusura se Ca2+ citosolico di una cellula aumenta
I 2 connessoni che formano la gap junction, a seconda di come interagiscono, possono formare un canale aperto o chiuso. Es. chiusura se Ca2+ citosolico di una cellula aumenta

50
Le cellule muscolari cardiache sono collegate “elettricamente” da gap junction.
La contrazione avviene in maniera coordinata in seguito alla diffusione del Ca2+ attraverso di esse (contrazione causata da aumento di Ca2+).
.")
51
Adesioni focali ed emidesmosomi (tra cellule e substrato sottostante)
Giunzioni tra cellule e matrice extracellulare: le integrine mediano due tipi di giunzioni stabili in cui il citoscheletro è legato (indirettamente) alla matrice extracellulare.
 alla matrice extracellulare.")
52
(recettori per MEC, collagene IV e laminina)
EMIDESMOSOMA Integrine (recettori per MEC, collagene IV e laminina)
")
53
ADESIONE FOCALE

54
Legame tra citoscheletro e MEC mediato da integrine
Le integrine legano la laminina, proteina che partecipa alla formazione della lamina basale Nelle adesioni focali, fasci di filamenti di actina sono ancorati alle subunità delle integrine, mediante associazione con diverse altre proteine, che includono la -actinina, la talina, e la vinculina. Negli emidesmosomi, l’integrina 64 collega la lamina basale ai filamenti intermedi, mediante la plectina.

55
Riassunto delle strutture di adesione e comunicazione tra cellule adiacenti

56
Un segnale extracellulare (fattore di crescita o altro) può modificare l’adesione di una cellula alle cellule adiacenti e/o alla matrice extracellulare
 può modificare l’adesione di una cellula alle cellule adiacenti e/o alla matrice extracellulare")







>")





 Le cellule distanti tra loro comunicano attraverso molecole (MEDIATORI o NEUROTRASMETTITORI)>")







