Scaricare la presentazione
2
I gas respiratori O 2 e CO 2 sono scambiati per diffusione semplice sia a livello dell’interfaccia alveoli-capillari polmonari sia a livello dell’interfaccia capillari sistemici-cellule. La diffusione obbedisce alla legge di Fick: Flusso Gradiente di concentrazione Permeabilità della membrana Spessore Area di scambio

3
Negli organismi biologici in genere il fattore determinante è il gradiente di concentrazione perché A, d e P sono massimizzate per favorire il processo. In fisiologia respiratoria ci si riferisce alle pressioni parziali dei gas in soluzione. Un gas si sposta da regioni a maggior pressione parziale verso regioni a minor pressione parziale

4
Prima di descrivere lo scambio dei gas vale la pena ricordare che il passaggio di gas da ambiente gassoso ad ambiente liquido è condizionato da alcuni fattori: Temperatura Temperatura Pressione parziale del gas all’interfaccia aria-liquido Pressione parziale del gas all’interfaccia aria-liquido Proprietà chimiche del gas e del liquido Proprietà chimiche del gas e del liquido La concentrazione di un gas in un liquido a temperatura costante è regolata dalla legge di Henry: C gas in soluzione = k · P

5
N.B.: All’equilibrio la pressione parziale di ossigeno è uguale nella fase gassosa e in soluzione. Questo però NON SIGNIFICA CHE LE CONCENTRAZIONI DI OSSIGENO SIANO UGUALI!!! Infatti la concentrazione di ossigeno disciolta dipende anche dalla SOLUBILITÁ dell’ossigeno (che in acqua è poco solubile) L’anidride carbonica è circa 20 volte più solubile dell’ossigeno in acqua. Quindi anche a uguali pressioni parziali la quantità di gas disciolta in acqua è maggiore per la CO 2 rispetto a O 2
 L’anidride carbonica è circa 20 volte più solubile dell’ossigeno in acqua. Quindi anche a uguali pressioni parziali la quantità di gas disciolta in acqua è maggiore per la CO 2 rispetto a O 2.")
6
Come avviene lo scambio dei gas respiratori nei polmoni e nei tessuti?

8
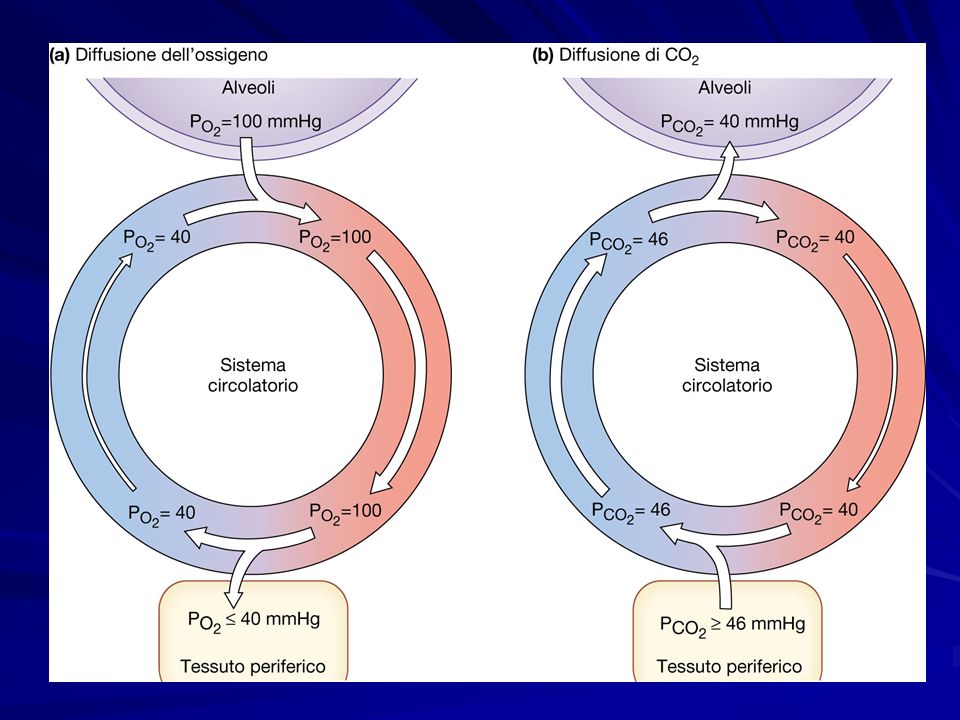
Sangue arterioso Sangue venoso PO2PO2PO2PO2100mmHg40mmHg P CO 2 40mmHg46mmHg pH7.407.37 Le pressioni parziali dei gas alveolari differiscono da quelle atmosferiche per tre motivi: Fra aria e sangue avvengono continuamente scambi L’aria alveolare è satura di vapor acqueo In seguito ad un’inspirazione aria atmosferica fresca si mescola con quella ricca di CO 2 dello spazio morto di conduzione

9
A cosa può essere dovuto un basso contenuto arterioso di ossigeno?

10
1.La riduzione della P O 2 alveolare diminuisce l’assunzione di ossigeno dai polmoni Esistono due cause possibili di una ridotta P O 2 alveolare: i.L’aria inspirata ha un basso contenuto di ossigeno (vedi altitudine) ii.La ventilazione alveolare è inadeguata (ipoventilazione)
 ii.La ventilazione alveolare è inadeguata (ipoventilazione)")
12
2. Le alterazioni della membrana alveolare modificano lo scambio di gas fra alveoli e capillari polmonari Lo scambio di ossigeno dagli alveoli al sangue richiede il passaggio attraverso la barriera formata da cellule alveolari e endotelio capillare. Questa diffusione può essere alterata per: i.Diminuzione dell’area disponibile per lo scambio gassoso ii.Aumento dello spessore della membrana alveolare iii.Aumento della distanza di diffusione

15
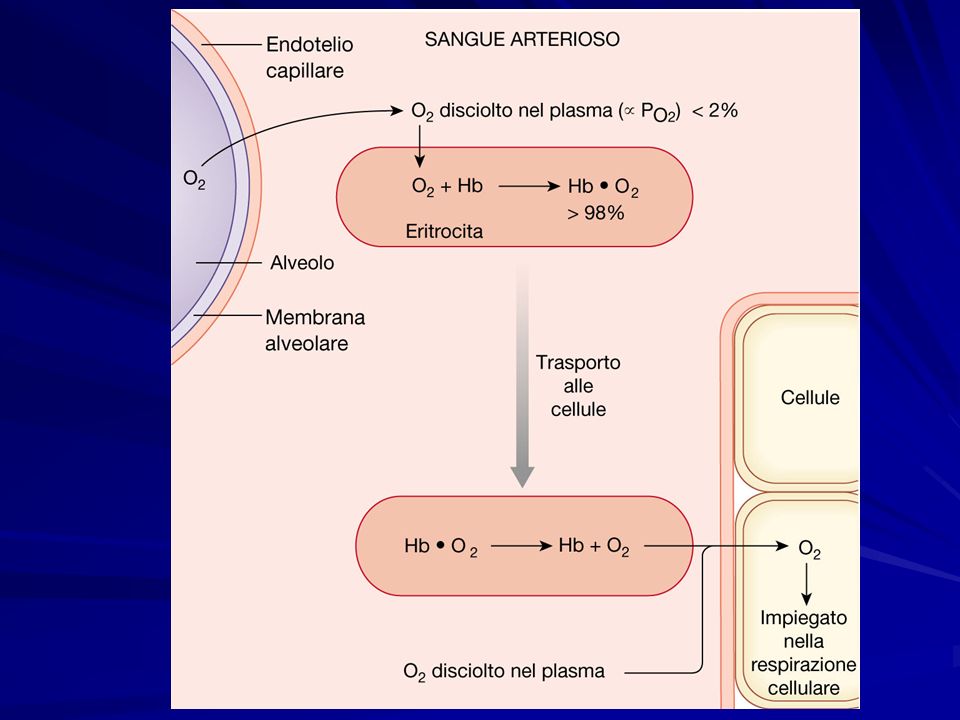
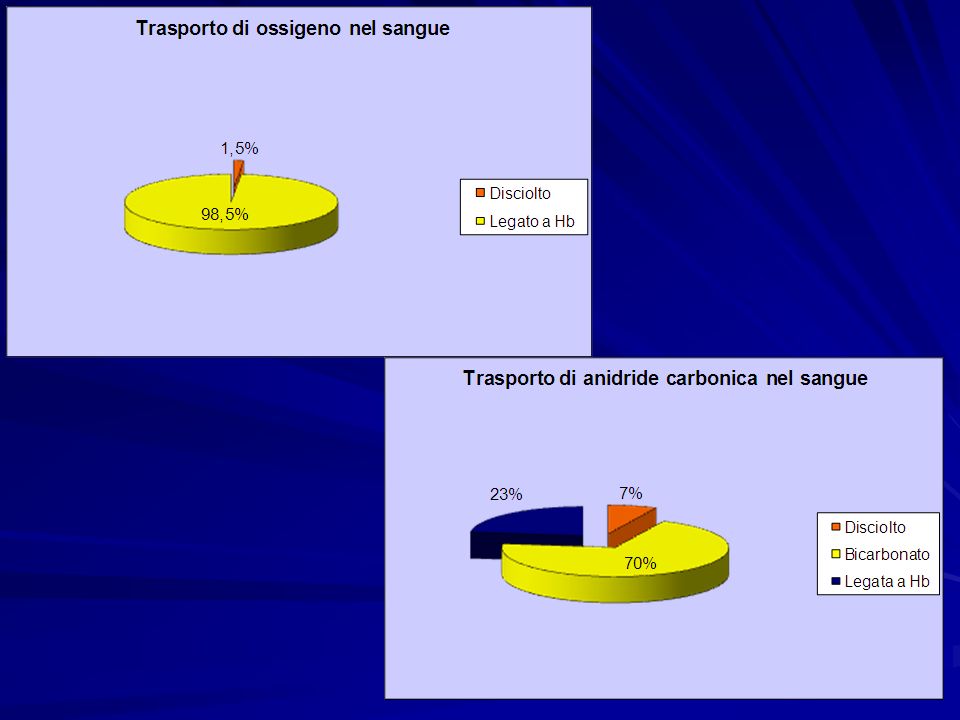
L’ossigeno è trasportato nel sangue con due meccanismi: in soluzione nel sangue (meno del 10% dell’ossigeno consumato che è ca 250ml/min a riposo) in soluzione nel sangue (meno del 10% dell’ossigeno consumato che è ca 250ml/min a riposo) legato all’emoglobina a cui si lega secondo la reazione reversibile legato all’emoglobina a cui si lega secondo la reazione reversibile Hb + O 2 ↔ HbO 2
 in soluzione nel sangue (meno del 10% dell’ossigeno consumato che è ca 250ml/min a riposo) legato all’emoglobina a cui si lega secondo la reazione reversibile legato all’emoglobina a cui si lega secondo la reazione reversibile Hb + O 2 ↔ HbO 2")
16
E’ possibile respirare senza emoglobina? La richiesta di O 2 in condizioni normali è di: 250 ml/min Con una G.C. di 5 l/min ed una quantità di O 2 sciolto sotto forma di gas di 3 ml/l, avremmo una disponibilità di O 2 di: 3 ml/l x 5 l/min = 15 ml/min Per far fronte a una richiesta di 250 ml/min la G.C. dovrebbe aumentare 16 volte mentre può al massimo crescere 9 volte

17
Respirazione con emoglobina L’emoglobina lega 20 ml di O 2 ogni 100 ml di sangue (cioè per esattezza 19.7 ml/100ml che equivalgono a 197 ml/l di sangue). Di questi ne scambia 5 ml/100 ml con i tessuti. 5 ml/100 ml scambiato equivalgono a 50 ml/l che moltiplicati per la G.C. (5 l/min) danno: 50 ml/l x 5 l/min = 250 ml/min che rappresenta la quantità di O 2 al minuto di cui abbiamo bisogno in condizioni normali 1gr Hb trasporta ca 1.35ml O 2 (numero di Hüfner) ca 146gr Hb / l di sangue ca 197ml O 2 / l di sangue
 danno: 50 ml/l x 5 l/min = 250 ml/min che rappresenta la quantità di O 2 al minuto di cui abbiamo bisogno in condizioni normali 1gr Hb trasporta ca 1.35ml O 2 (numero di Hüfner) ca 146gr Hb / l di sangue ca 197ml O 2 / l di sangue.")
18
A M

20
Emoglobina = proteina con elevatissima capacità di trasporto per O 2 Proteina con struttura quaternaria (M.W.= 68kDa) formata da 4 sub-unità ciascuna avvolta attorno ad un gruppo eme (fra loro identici) contenente un atomo di Fe che lega una molecola di ossigeno: pertanto la molecola di emoglobina può trasportare fino a 4 molecole di ossigeno Ossigenazione dell’emoglobina e non ossidazione!!!
 formata da 4 sub-unità ciascuna avvolta attorno ad un gruppo eme (fra loro identici) contenente un atomo di Fe che lega una molecola di ossigeno: pertanto la molecola di emoglobina può trasportare fino a 4 molecole di ossigeno Ossigenazione dell’emoglobina e non ossidazione!!!")
21
Adulto (HbA) α 2 β 2 Adulto (2%) (HbA 2 ) α 2 δ 2 Feto (HbF) α 2 γ 2
 α 2 β 2 Adulto (2%) (HbA 2 ) α 2 δ 2 Feto (HbF) α 2 γ 2")
22
Il legame fra ossigeno ed emoglobina segue la legge dell’azione di massa Hb + O 2 ↔ HbO 2 Verso dx se aumenta la concentrazione di ossigeno Verso sx se la concentrazione di ossigeno diminuisce

23
cambia colore se legata a O 2 HbO 2 (rosso vivo; ossiemoglobina) Hb (rosso bruno; deossiemoglobina) l’O 2 si lega al Fe 2+ del gruppo eme ma non lo ossida: forma un legame reversibile ad alta affinità. se il Fe 2+ diventa Fe 3+, l’Hb diventa metaemoglobina (composto stabile con l’O 2, cessa di trasportare O 2 ). la metaemoglobina-reduttasi ripristina i livelli di Hb.
. la metaemoglobina-reduttasi ripristina i livelli di Hb..")
24
Diffusione favorita Ingresso dell’ossigeno disciolto nel plasma Rilascio di ossigeno da Hb Diffusione all’interno delle cellule dell’ossigeno disciolto nel plasma; questo riduce la P o 2 nel plasma e tende a far spostare la reazione verso dx con liberazione di O 2 dall’Hb

25
Il legame di ossigeno all’emoglobina si esprime in percentuale. Per un certo valore di P O 2 vale: Quantità di O 2 legata Quantità max che può essere legata X 100 = Saturazione % dell’emoglobina

26
Che relazione esiste fra P O 2 e quantità di ossigeno legata all’emoglobina? In un muscolo in forte esercizio P O 2 scende a 20mmHg e la saturazione di Hb al 35% ca 98% In questo range la saturazione di Hb non scende troppo 75% 26.8 A livello tissutale P O 2 =40mmHg Hb cede il 25% dell’ossigeno trasportato, restando satura al 75% L’andamento sigmoide della curva sta ad indicare una cooperazione positiva fra le quattro subunità della molecola di Hb

27
la quantità di O 2 legata all’Hb è proporzionale alla P O 2 legame reversibile 100% di saturazione (raggiungibile solo per P O 2 pari a 600mmHg!!) equivale alla capacità massima del sangue di legare l’O 2 la [Hb] nel sangue è 15gr/100ml 1 gr di Hb lega 1.35 ml di O 2 l’Hb lega: 15gr/100 ml x 1.35 ml O 2 = 20 ml O 2 /100 ml sangue
![ la quantità di O 2 legata all’Hb è proporzionale alla P O 2 legame reversibile 100% di saturazione (raggiungibile solo per P O 2 pari a 600mmHg!!) equivale alla capacità massima del sangue di legare l’O 2 la [Hb] nel sangue è 15gr/100ml 1 gr di Hb lega 1.35 ml di O 2 l’Hb lega: 15gr/100 ml x 1.35 ml O 2 = 20 ml O 2 /100 ml sangue](http://images.slideplayer.it/33/10307636/slides/slide_27.jpg " la quantità di O 2 legata all’Hb è proporzionale alla P O 2 legame reversibile 100% di saturazione (raggiungibile solo per P O 2 pari a 600mmHg!!) equivale alla capacità massima del sangue di legare l’O 2 la [Hb] nel sangue è 15gr/100ml 1 gr di Hb lega 1.35 ml di O 2 l’Hb lega: 15gr/100 ml x 1.35 ml O 2 = 20 ml O 2 /100 ml sangue")
28
la mioglobina è formata da una sola subunità di globina possiede alta affinità per l’O 2. E’ contenuta nel m. scheletrico ed è saturata a P O 2 = 40 mmHg (sangue venoso) è una riserva di O 2 muscolare, che viene ceduto quando l’O 2 diminuisce
 è una riserva di O 2 muscolare, che viene ceduto quando l’O 2 diminuisce.")
29
l’Hb ha una affinità 250 volte maggiore per il CO il CO sposta la curva di saturazione verso sinistra (maggiore affinità) P CO molto basse saturano l’Hb lo scambio di O 2 in presenza di 50% HbCO è fortemente sfavorito
 P CO molto basse saturano l’Hb lo scambio di O 2 in presenza di 50% HbCO è fortemente sfavorito")
30
Effetti di pH, T e P CO 2 N.B.: Variazioni molto pronunciate nella parte ripida della curva. Questo significa che le variazioni hanno molto effetto a livello tissutale (P O 2 fra 20 e 40mmHg) più che a livello polmonare (P O 2 fra 90 e 100mmHg)
 più che a livello polmonare (P O 2 fra 90 e 100mmHg).")
31
Variazione di pH = effetto Bohr 75% 57% Forte esercizio fisico Anaerobiosi Produzione di ac. lattico Rilascio di H + nel LEC Questo significa che in condizioni di acidosi il sangue rilascia più O 2 ai tessuti che in quel momento ne hanno bisogno

32
Effetto del 2,3-DPG DPG deriva da un intermedio della glicolisi. Una ipossia ne aumenta i livelli riducendo l’affinità di legame di Hb e ossigeno. Altitudine e anemia sono le cause più frequenti

33
HbA e HbF Migliore capacità di legare O 2 in ambiente placentare dove il tenore di ossigeno è basso

34
Riassumendo

35
Trasporto di CO 2

36
Prodotto di scarto della respirazione cellulare che determina acidosi, alterazioni del SNC fino alla morte Più solubile dell’ossigeno nei liquidi corporei Prodotta in quantità notevoli rispetto a quella trasportata nel plasma

37
Il trasporto di CO 2 nel sangue segue tre vie - disciolta nel sangue 7% - diffusione nei globuli rossi 93% di cui 70% convertita in bicarbonato* 23% legata a Hb *Questa quota ha due scopi: Fornire un meccanismo per il trasporto di CO 2Fornire un meccanismo per il trasporto di CO 2 Rendere il bicarbonato disponibile come tamponeRendere il bicarbonato disponibile come tampone

38
diffusione Scambio dei cloruri (elettroneutro) Effetto tampone di Hb *Si può scrivere semplicemente come: CO 2 + H 2 O ↔ H + + HCO 3 - Reazione reversibile che obbedisce alla legge dell’azione di massa * Per mantenere la reazione spostata verso dx e neutralizzare così la CO 2 che si forma in continuazione, vengono sottratti i prodotti della reazione. HCO 3 - scambiato con Cl - in antiporto Gli ioni H + sono bufferati da Hb. Quando la CO 2 aumenta troppo la Hb non riesce a neutralizzare tutti gli idrogenioni che si accumulano determinando acidosi respiratoria Formazione di carbaminoemoglobina. La sua formazione è facilitata anche perché H + e CO 2 riducono l’affinità di Hb per O 2

39
Al diminuire della P CO 2 plasmatica la CO 2 disciolta nell’eritrocita diminuisce spostando la reazione verso dx. Lo scambio dei cloruri si inverte e gli idrogenioni lasciano l’Hb

40
Scambio dei cloruri e legame di H + con Hb forniscono due sistemi volti a sottrarre CO 2 e garantire così il gradiente necessario al suo ingresso nell’eritrocita. Nei casi di eccessiva produzione di CO 2 la Hb non riesce a tamponare tutti i protoni prodotti che passano quindi nel plasma causando ACIDOSI RESPIRATORIA

41
Abbiamo visto che circa il 23% di CO 2 si lega a Hb e in particolare sui gruppi NH 2 esposti formando così carbaminoemoglobina. Tale legame è facilitato dalla presenza di CO 2 e H + i quali riducono entrambi l’affinità di Hb per l’ossigeno. CO 2 + Hb HbCO 2

42
TRASPORTO AI POLMONI DI CO 2 P CO 2 alv < P CO 2 venoso

43
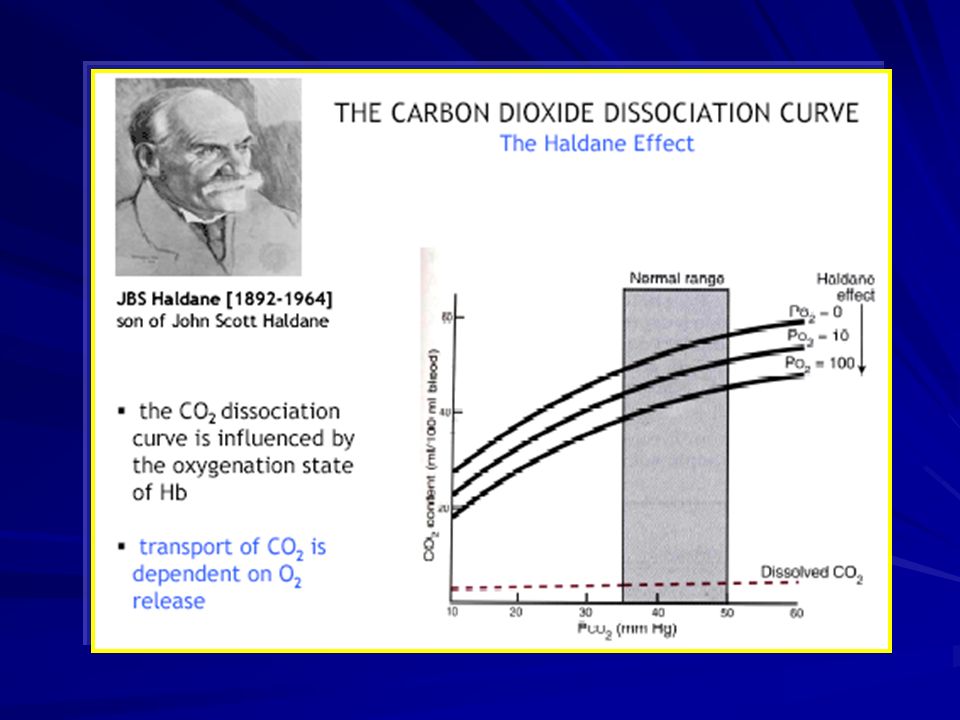
Effetto Haldane La rimozione di ossigeno dall’emoglobina (a livello tessutale) aumenta l’affinità dell’emoglobina per l’anidride carbonica. Questo permette alla CO 2 di essere “caricata” sull’Hb vuota

45
A livello dei tessuti: l’Hb deossigenata lega meglio la CO 2 formando composti carbammino- emoglobinici (effetto Haldane). l’effetto Haldane è responsabile di circa la metà della CO 2 che viene scambiata e trasportata a livello dei tessuti.

46
A livello alveolare: la formazione di HbO 2 riduce il legame di CO 2 e H + con l’Hb la maggiore CO 2 disponibile viene scambiata con l’alveolo ed espulsa

47
C

50
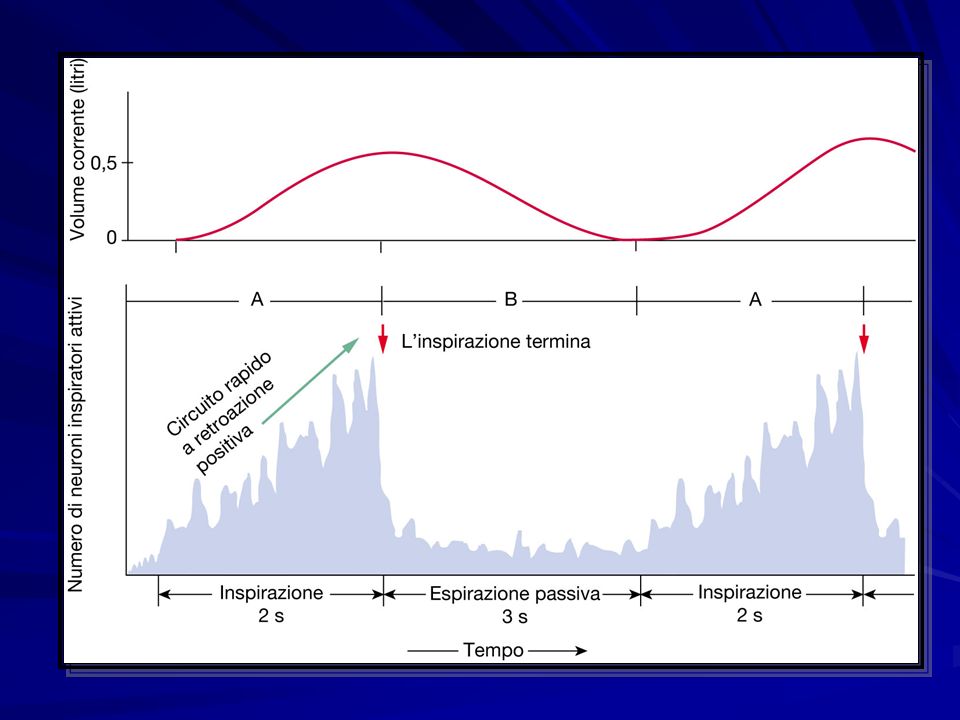
Processo ritmico, non cosciente A differenza del battito cardiaco però il muscolo scheletrico non è dotato di automaticità ma riceve innervazione da motoneuroni somatici che a loro volta prendono ordini dal SNC Meccanismo altamente regolato

51
G V D G V V

52
I centri di controllo respiratori sono localizzati a livello del ponte e del bulbo. I neuroni formano un generatore centrale di rimo respiratorio. Tale rete è difficile da studiare per la complessità e la localizzazione. La maggior parte di informazioni derivano da pazienti con lesioni cerebrali. Esiste un modello atto a spiegare il meccanismo, i cui punti fondamentali sono i seguenti

53
Neuroni del ponte modulano la ventilazione La ritmicità deriva da un insieme di neuroni che scaricano spontaneamente La ventilazione è soggetta a modulazione da parte di vari fattori chimici e dei centri cerebrali superiori Neuroni respiratori bulbari controllano inspirazione (GVD) ed espirazione (GVV)
 ed espirazione (GVV)")
54
Il centro respiratorio bulbare è controllato continuamente nel suo funzionamento dai livelli plasmatici di CO 2, O 2 e H +. I neuroni bulbari sono raggruppati in due importanti nuclei che sono: gruppo ventilatorio dorsale, INSPIRATORIOgruppo ventilatorio dorsale, INSPIRATORIO gruppo ventilatorio ventrale, ESPIRATORIO e controllo dei muscoli inspiratori implicati nell’inspirazione profondagruppo ventilatorio ventrale, ESPIRATORIO e controllo dei muscoli inspiratori implicati nell’inspirazione profonda

56
CO 2, O 2 e pH La CO 2 è lo stimolo principale per modificare la ventilazione. I chemocettori per ossigeno e anidride carbonica sono associati alla circolazione arteriosa. Quando O 2 diminuisce troppo o la CO 2 aumenta, il centro inspiratorio viene stimolato ad aumentare la ventilazione. I chemocettori comprendono chemocettori periferici (glomi aortici e carotidei) e chemocettori centrali (superficie ventrale del bulbo)
 e chemocettori centrali (superficie ventrale del bulbo).")
57
L’ossigeno non è comunque un fattore importante nel determinare la ventilazione perché P O 2 deve scendere sotto a 60mmHg prima di stimolare la ventilazione*. I chemocettori periferici quindi non svolgono un ruolo quotidiano nel controllo della ventilazione * Condizione compatibile con un’altitudine di 3000m

58
Il fattore fondamentale nel controllo della ventilazione è la CO 2 che agisce sui chemocettori centrali Va notato che gli H + presenti nel plasma passano male attraverso la BEE e hanno quindi scarso effetto sui chemocettori centrali

59
Quando P CO 2 aumenta, la CO 2 passa rapidamente la barriera emato-encefalica andando ad attivare i chemocettori centrali che segnalano al generatore centrale di ritmo respiratorio di aumentare la frequenza e la profondità degli atti respiratori. In realtà il parametro che i chemocettori valutano è il valore di pH in base alla seguente reazione CO 2 + H 2 O ↔ H 2 CO 3 ↔ H + + HCO 3 -

60
Questi chemocettori centrali si adattano all’ipercapnia cronica, mentre i chemocettori periferici non si adattano all’ipossia cronica: in questi casi, come ad esempio nell’enfisema, la bassa P O 2 rimane l’unico stimolo alla ventilazione

61
Accanto a quanto visto esistono anche riflessi protettivi che difendono il polmone (riflesso di distensione di Hering-Breuer)
")
62
La ventilazione è inoltre modulata da centri superiori come ipotalamo e sistema limbico. Va però detto che non esiste modo per bloccare i riflessi mediati da chemocettori

63
Per chi fosse interessato http://www.ismmed.org/np_altitude_tutorial.htm http://www.high-altitude-medicine.com/



















, specializzato per la distribuzione di: gas.>")


 * frequenza respiratoria (500 - 150) * 12 = 4.5 l/min corrente * frequenza respiratoria ventilazione.>")

>")
