Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La batteriorodopsina: una pompa protonica

2
Halobacterium sp. ceppo NRC-1
5 mm

3
La Storia della batteriorodopsina
1975: Studi pioneristici di Unwin & Henderson. Cristalli bidimensionali con struttura ricostruita a 7 Å di risoluzione mediante microscopia elettronica Grande impatto sugli esperimenti successivi e le teorie per la predizione delle eliche transmembrana. 1980: Michel cristallizza la proteina : Struttura EM a ~3 Å di risoluzione, si osserva il retinale legato. 1998: Struttura ai raggi X a 2 Å di risoluzione, e successivamente si ottengono molte strutture in stati meccanicistici differenti 2000: Si comprende il meccanismo della traslocazione protonica in grande dettaglio (grazie alla spettroscopia FT-IR e alla mutagenesi sito-specifica)
")
4
Premio Nobel per la chimica (1988)
Hartmut Michel primo a cristallizzare la Br nel 1980

5
Le proprieta’ della batteriorodopsina
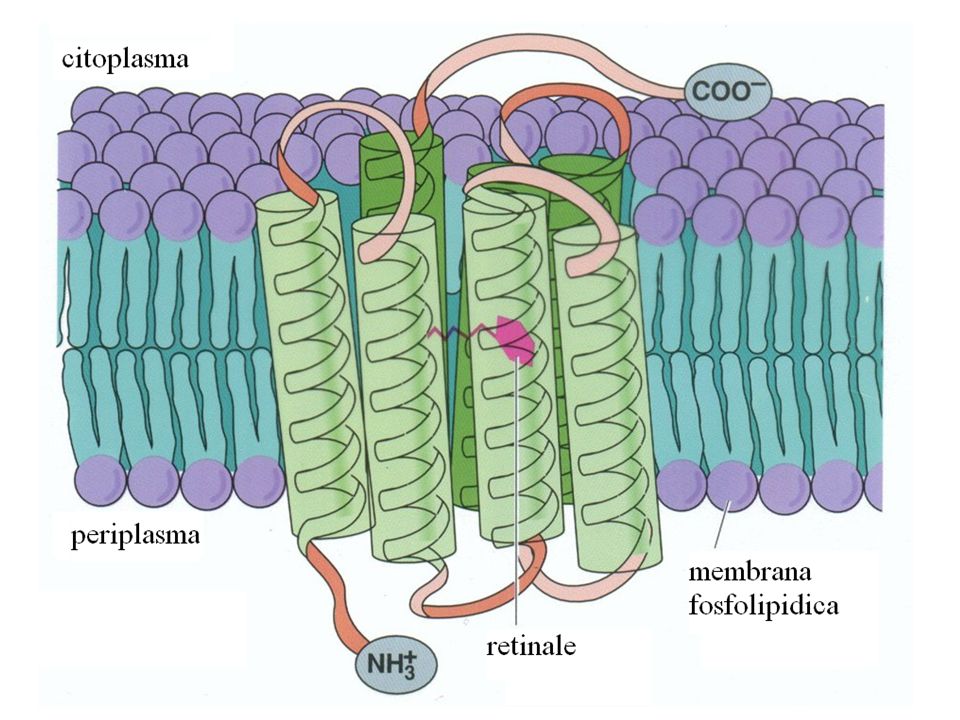
Catena polipeptidica unica di 248 amminoacidi (determinata dal sequenziamento sia della proteina che del DNA) La proteina è sintetizzata nella cellula nella forma di un precursore con una sequenza N-terminale segnale di 13 amminoacidi e di un residuo di Asp C-terminale che vengono rimossi. La proteina matura pesa 26 kD. L’analisi di sequenza mostra la presenza di 7 a-eliche. Il C-terminale è rivolto verso il citoplasma mentre l’N-terminale è esposto allo spazio extracellulare. Il gruppo prostetico è costituito dal trans-retinale legato alla K216 mediante una base di Schiff.
 La proteina è sintetizzata nella cellula nella forma di un precursore con una sequenza N-terminale segnale di 13 amminoacidi e di un residuo di Asp C-terminale che vengono rimossi. La proteina matura pesa 26 kD. L’analisi di sequenza mostra la presenza di 7 a-eliche. Il C-terminale è rivolto verso il citoplasma mentre l’N-terminale è esposto allo spazio extracellulare. Il gruppo prostetico è costituito dal trans-retinale legato alla K216 mediante una base di Schiff.")
7
La struttura primaria e la topologia
Khorana, JBC, 1988, 263, 7439

8
Kyte J, Doolittle RF (May 1982).
"A simple method for displaying the hydropathic character of a protein". J. Mol. Biol. 157 (1): 105–32.
: 105–32.")
9
Le scale di idropatia scala Kyte & Doolittle costante di ripartizione
Arg -4.5 Asn -3.5 Asp -3.5 Cys 2.5 Gln -3.5 Glu -3.5 Gly -0.4 His -3.2 Ile 4.5 Leu 3.8 Lys -3.9 Met 1.9 Phe 2.8 Pro -1.6 Ser -0.8 Thr -0.7 Trp -0.9 Tyr -1.3 Val 4.2 costante di ripartizione L’energia libera per il trasferimento dell’amminoacido dall’acqua al benzene è una misura della idrofobicità benzene H2O

10
Gli algoritmi moderni permettono di stimare la topologia della proteina ( TGRPEWIWLALGTALMGLGTLYFLVKGMGVSDPDAKKFYAITTLVPAIAFTMYLSMLL GYGLTMVPFGGEQNPIYWARYADWLFTTPLLLLNLALLVDADQGTILALVGADGIMIG TGLVGALTKVYSYRFVWWAISTAAMLYILYVLFFGFSMRPEVASTFKVLRNVTVVLWS AYPVVWLIGSEGAGIVPLNIETLLFMVLDVSAKVGFGLILLRSRAIFG out /in membrana topologia=o6-25i38-60o75-97i o i o i

12
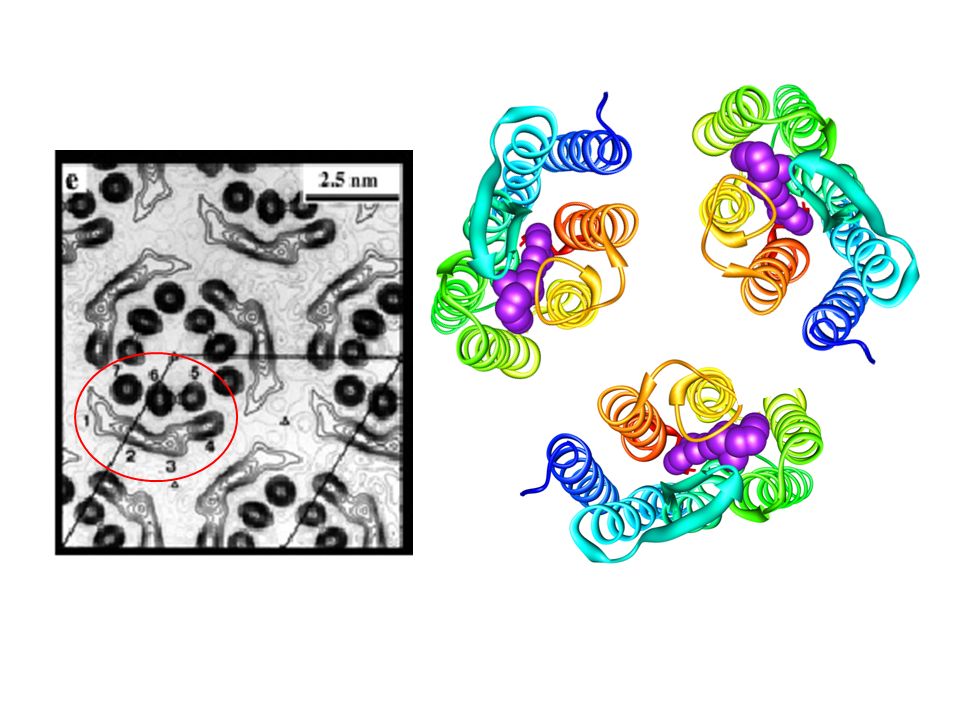
I cristalli bidimensionali

13
Analisi della struttura
I monomeri della batteriorodopsina formano trimeri molto stabili come suggerito dalle immagini al ME. I contatti intermolecolari elica-elica corrispondono a circa Å2 (circa il 27% della superficie del monomero). E’ presente un ponte salino intermolecolare fra K40 e D104
. E’ presente un ponte salino intermolecolare fra K40 e D104.")
14
Le 7 a-eliche ottenute per microscopia elettronica

16
I cristalli

17
Strutture della batteriorodopsina pubblicate
1975 microscopia elettronica (Henderson & Unwin, Nature) 7 Å 1990 microscopia elettronica (Henderson et al. J. Mol. Biol.) 3.5 Å 1996 microscopia elettronica (Grigorieff et al.J. Mol. Biol.) 3.5 Å 1997 microscopia elettronica (Kimura et al. Nature) 3.0 Å 1997 diffrazione raggi X (Pebay-Peyroula et al. Science) 2.5 Å 1998 diffrazione raggi X (Luecke et al. Science) 2.3 Å 1998 diffrazione raggi X (Essen et al. PNAS) 2.9 Å 1998 diffrazione raggi X (Takeda et al. J. Mol. Biol.) 3.5 Å 1999 microscopia elettronica (Mitsuoka et al. J. Mol. Biol.) 3.0 Å 1999 diffrazione raggi X (Belrhali et al. Structure) 1.9 Å 1999 diffrazione raggi X (Luecke et al. J. Mol. Biol.) 1.55 Å le strutture sono simili nelle a eliche differenze nelle anse interelica molecole d’acqua e lipidi visibili alle risoluzioni più grandi
 7 Å microscopia elettronica (Henderson et al. J. Mol. Biol.) 3.5 Å microscopia elettronica (Grigorieff et al.J. Mol. Biol.) 3.5 Å microscopia elettronica (Kimura et al. Nature) 3.0 Å diffrazione raggi X (Pebay-Peyroula et al. Science) 2.5 Å diffrazione raggi X (Luecke et al. Science) 2.3 Å diffrazione raggi X (Essen et al. PNAS) 2.9 Å diffrazione raggi X (Takeda et al. J. Mol. Biol.) 3.5 Å microscopia elettronica (Mitsuoka et al. J. Mol. Biol.) 3.0 Å diffrazione raggi X (Belrhali et al. Structure) 1.9 Å diffrazione raggi X (Luecke et al. J. Mol. Biol.) 1.55 Å. le strutture sono simili nelle a eliche. differenze nelle anse interelica. molecole d’acqua e lipidi visibili alle risoluzioni più grandi.")
18
Banca dati delle proteine di membrana

19
La struttura ad alta risoluzione
C-ter il retinale è inserito in una tasca costituita dalle eliche B-G F G B D C G F A E ~ 40Å A E B C D N-ter ~ 30Å spazio extracellulare

20
La Br è estremamente idrofobica lato citosolico
membrana lato extracellulare pdb:1c3w

21
Analisi della struttura
elica B elica C Le anse interelica hanno struttura irregolare. La connessione fra le eliche B e C forma un un foglietto b a 2 filamenti antiparalleli stabilizzato da 6 legami idrogeno

22
Analisi della struttura
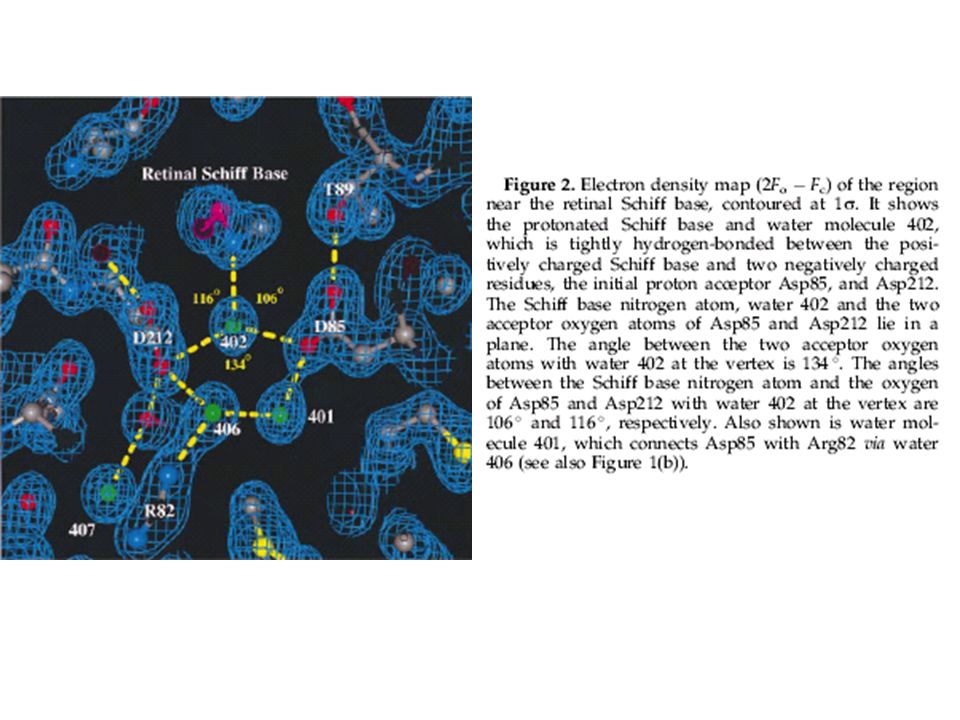
Vi è una rete estesa di legami idrogeno interelica (31) che stabilizzano la struttura terziaria del monomero che non risulta essere influenzato dall’arrangiamento trimerico nella membrana Il retinale è posto in una tasca delimitata da molti residui: W86, T89, L93, T90, M118, W138, S141, T142, M145, W182, Y185, W189, D212 E’ presente una rete estesa di legami idrogeno vicino alla base di Schiff fra residui amminoacidici e 7 molecole di acqua, di importanza meccanicistica
 che stabilizzano la struttura terziaria del monomero che non risulta essere influenzato dall’arrangiamento trimerico nella membrana. Il retinale è posto in una tasca delimitata da molti residui: W86, T89, L93, T90, M118, W138, S141, T142, M145, W182, Y185, W189, D212. E’ presente una rete estesa di legami idrogeno vicino alla base di Schiff fra residui amminoacidici e 7 molecole di acqua, di importanza meccanicistica.")
23
Vi è una rete estesa di legami idrogeno interelica (31) che stabilizzano la struttura terziaria del monomero K30-L224 T170-S226 A G F G Y185-D212

24
Il retinale è posto in una tasca delimitata da molti residui: W86, T89, L93, T90, M118, W138, S141, T142, M145, W182, Y185, W189, D212

25
(a) Region of the active site that contains an all trans retinal
(a) Region of the active site that contains an all trans retinal. The plane of Trp86 and Trp182 flank the polyene chain between the 9-methyl and 13-methyl groups, and contribute to its immobilization in the retinal-binding pocket. Part of the hydrogen-bonded chain that leads from the Schiff base to the extracellular surface via water 402, Asp85, and water 401, is visible in this view. Water 501, which connects the peptide CO of Ala215 with the indole NH of Trp182, is also visible. The corresponding omit map with all retinal atoms omitted from the model still features a hole in the b-ionone ring, and can be viewed at hudel/br/1.55/ret omitmap.gif.
 Region of the active site that contains an all trans retinal. The plane of Trp86 and Trp182 flank the polyene chain between the 9-methyl and 13-methyl groups, and contribute to its immobilization in the retinal-binding pocket. Part of the hydrogen-bonded chain that leads from the Schiff base to the extracellular surface via water 402, Asp85, and water 401, is visible in this view. Water 501, which connects the peptide CO of Ala215 with the indole NH of Trp182, is also visible. The corresponding omit map with all retinal atoms omitted from the model still features a hole in the b-ionone ring, and can be viewed at hudel/br/1.55/ret omitmap.gif.")
26
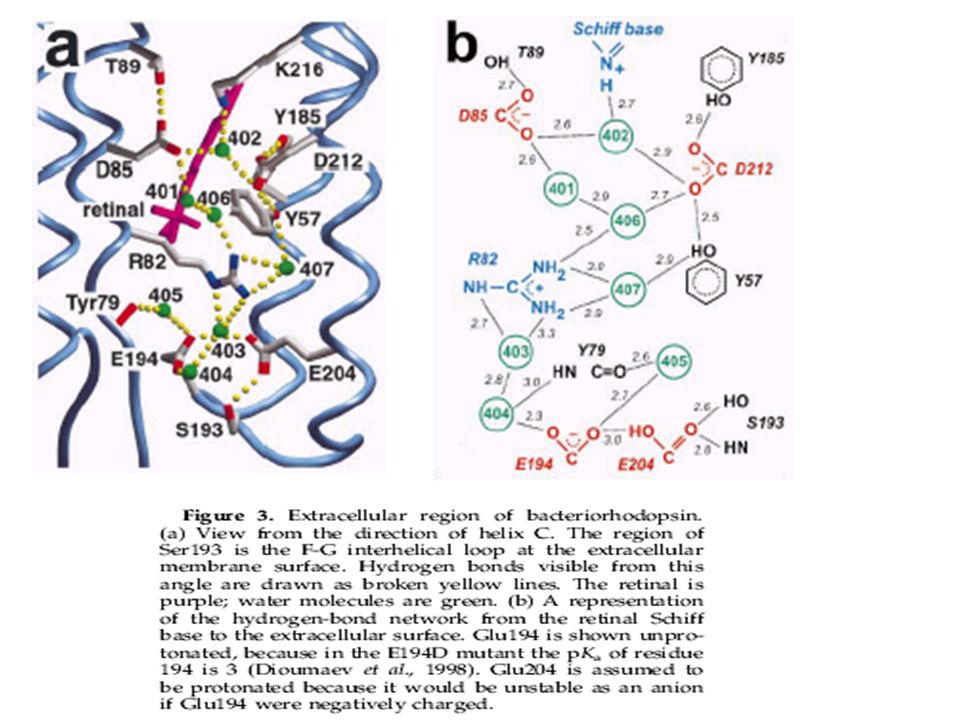
(b) Region of Arg82 in the extracellular half channel
(b) Region of Arg82 in the extracellular half channel. In the top portion, the guanidinium moiety is connected to the Schiff base via water 406, water 401, and Asp85 (see also Figures 2 and 3(b)). Water 407 further stabilizes the positive charge by accepting hydrogen bonds from both guanidinium nitrogen atoms. In the bottom portion, the connection to the extracellular surface continues via water 403, water 404 (not visible from this angle), Glu194 and Glu204. An omit map of this region, with all Arg82, Glu194 and Glu204 side-chain atoms and surrounding water molecules omitted from the model, can be viewed at /r82 omitmap.gif.
 Region of Arg82 in the extracellular half channel. In the top portion, the guanidinium moiety is connected to the Schiff base via water 406, water 401, and Asp85 (see also Figures 2 and 3(b)). Water 407 further stabilizes the positive charge by accepting hydrogen bonds from both guanidinium nitrogen atoms. In the bottom portion, the connection to the extracellular surface continues via water 403, water 404 (not visible from this angle), Glu194 and Glu204. An omit map of this region, with all Arg82, Glu194 and Glu204 side-chain atoms and surrounding water molecules omitted from the model, can be viewed at /r82 omitmap.gif.")
29
Il retinale è sintetizzato a partire dalla vitamina A

31
Il gruppo prostetico della batteriorodopsina
il trans-retinale nella BR forma una base di Schiff con K216 L’assorbimento di luce induce una isomerizzazione trans-cis 1c8r 1c8s Luecke et al.Science 1999, 286,

32
La fotoisomerizzazione del retinale eliche G
trans-retinale K216 11 13 9 7 hn K216 13 13-cis-retinale pdb: 1c8r e 1c8s

33
Ogni pompa ionica deve fornire un accesso alternato allo ione da trasportare sui 2 lati della membrana

34
Nella batteriorodopsina il retinale interrompe la continuita’ fra 2 emi-canali che connettono lo spazio citoplasmatico (alto) allo spazio extracellulare (basso) luce

35
Il fotociclo

36
Lo spettro di assorbimento dipende dall’intorno del cromoforo
Lo spettro di assorbimento dipende dall’intorno del cromoforo. Un esempio il triptofano

37
lato citoplasmatico H+ H+ lato extracellulare K216 D96 K216 D85 ms fs
retinale tutto trans H+ K216 D96 retinale 13-cis K216 D85 ms fs retinale R82 ms ps E194 E204 ms μs H+ μs lato extracellulare

38
luce

Presentazioni simili














>")
>")




>")

