Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Ottimizzare l'espressione
Non esiste una singola strategia/procedura che funzioni adeguatamente per tutti i casi Ciascuna proteina è un prodotto in sé unico C

2
CODON USAGE Quando un gene clonato utilizza codoni ai quali la cellula ospite ricorre raramente, si verifica un’incompatibilità cellulare che può interferire con una traduzione efficiente Può accadere che la cellula ospite non produca a sufficienza i tRNA che riconoscono i suddetti codoni di impiego raro e la resa in proteina del gene clonato risulta conseguentemente inferiore alle aspettative La presenza di insufficienti quantità di tRNA può determinare infatti un rallentamento della traduzione, un arresto precoce della traduzione o l’incorporazione di un aminoacido errato con conseguente sintesi di forme varianti della molecola proteica di interesse Di solito la presenza di un piccolo numero di codoni rari non determina un abbassamento significativo dei livelli di espressione Gli effetti sui livelli di espressione possono essere severi se nel gene clonato i codoni rari sono numerosi o presenti in clusters Gli effetti più severi sull’espressione sono stati osservati in presenza di più codoni rari consecutivi localizzati all’estremità N-terminale della sequenza codificante

3
Codoni rari in E.coli Geni di E.coli con elevati livelli di espressione (195 geni) 4290 geni di E. coli analizzati

4
STRATEGIE POSSIBILI 1. SODDISFARE LA RICHIESTA
Problemi di traduzione simili a quelli determinati dal codon usage possono essere provocati da alti livelli di espressione di proteine caratterizzate da un aminoacido abbondante. In questo caso il livello di espressione può essere migliorato fornendo l’aminoacido limitante nel mezzo di coltura. STRATEGIE POSSIBILI 1. SODDISFARE LA RICHIESTA E’ stato dimostrato che il livello di espressione di una proteina il cui gene contiene codoni rari può essere aumentato in maniera significativa rifornendo la cellula ospite dei corrispondenti tRNA Si possono inserire i geni codificanti i tRNA in questione in un plasmide, che può essere lo stesso vettore di espressione o un plasmide compatibile E’ stato creato un vettore, pRARE, contenente i geni codificanti i tRNA per i codoni rari di Arg, Ile, Gly, Leu e Pro

5
argU riconosce le triplette AGG/AGA
Codoni rari in E.coli Geni di E.coli con elevati livelli di espressione (195 geni) 4290 geni di E.coli analizzati tRNAs rari o assenti argU riconosce le triplette AGG/AGA CCC CUA GGA AUA
 4290 geni di E.coli analizzati. tRNAs rari o assenti. argU riconosce le triplette AGG/AGA. CCC. CUA. GGA. AUA.")
6
STRATEGIE POSSIBILI 2. OTTIMIZZAZIONE DEI CODONI
Tali plasmidi sono stati adoperati per trasformare diversi ceppi di E.coli E’ stata così creata la serie RosettaTM di ospiti batterici per l’espressione Tali plasmidi sono compatibili con i vettori pET e con altri vettori di espressione batterici I ceppi Rosetta hanno consentito di ottenere elevati livelli di espressione di proteine i cui geni contenevano codoni rari di E.coli STRATEGIE POSSIBILI 2. OTTIMIZZAZIONE DEI CODONI Produzione di una versione del gene contenente i codoni utilizzati più comunemente dalla cellula ospite SINTESI CHIMICA MUTAGENESI

7
CEPPO ROSETTA Ceppo di E.coli BL21(DE3)pLysS
Presenta mutazioni della proteasi lon ed è privo della proteasi della membrana esterna ompT, ciò che riduce gli eventi proteolitici a carico delle proteine ricombinanti E’ un ospite lisogeno di λDE3 e dunque presenta nel cromosoma una copia del gene codificante la RNA polimerasi T7, sotto il controllo del promotore lacUV5 L’espressione proteica è sotto il controllo del promotore T7 Presenta un plasmide contenente una copia del gene che codifica il lisozima T7, espresso a bassi livelli Il lisozima T7 lega l’RNA polimerasi T7 inibendo la trascrizione guidata da questo enzima In seguito ad induzione con IPTG, la produzione ad elevati livelli dell’RNA polimerasi T7 sopprime ogni inibizione da parte del lisozima T7 Analisi dell’espressione di un mutante di delezione dell’attivatore tessutale del plasminogeno umano BL21 (DE3) pLysS Rosetta (DE3) pLysS BL21 (DE3) Rosetta (DE3) M I U I U M I U I U Il costrutto codifica una proteina di 358 aminoacidi del peso molecolare di 40 kDa e contiene 9 CCC, 8 AGA/AGG, 6 GGA, 6 CGG/CGA, 2 AUA, 1 CUA per un totale di 32 codoni rari (8,9%) E’ riportata un’analisi mediante SDS-PAGE di estratti cellulari totali
 pLysS. Rosetta. (DE3) pLysS. BL21. (DE3) Rosetta. (DE3) M I U I U M I U I U. Il costrutto codifica una proteina di 358 aminoacidi del. peso molecolare di 40 kDa e contiene 9 CCC, 8 AGA/AGG, 6 GGA, 6 CGG/CGA, 2 AUA, 1 CUA per un totale di 32. codoni rari (8,9%) E’ riportata un’analisi mediante SDS-PAGE di estratti. cellulari totali.")
8
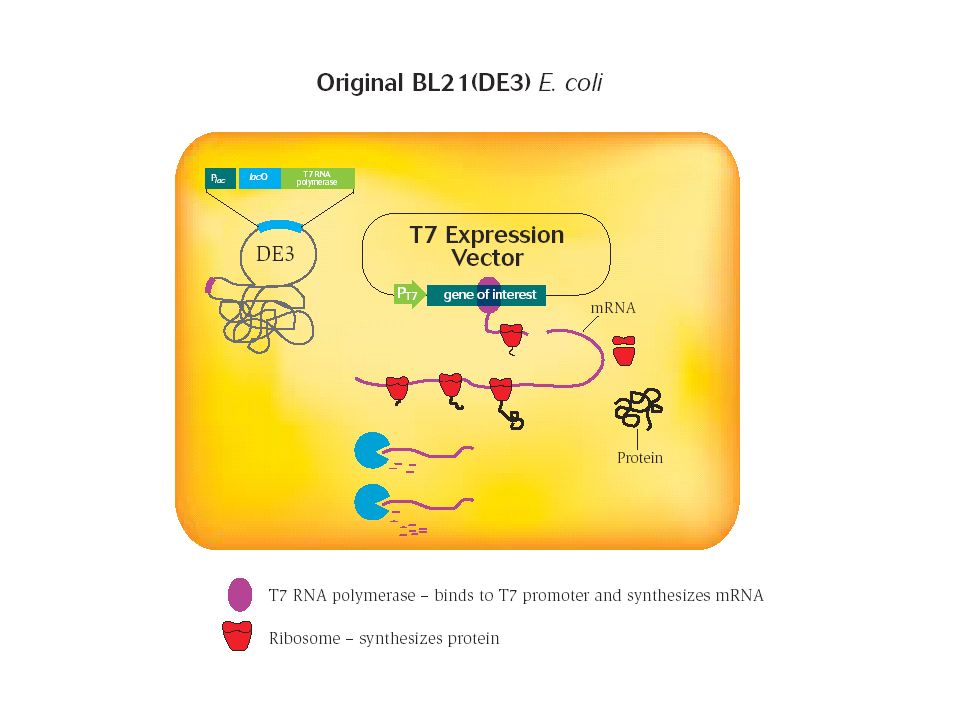
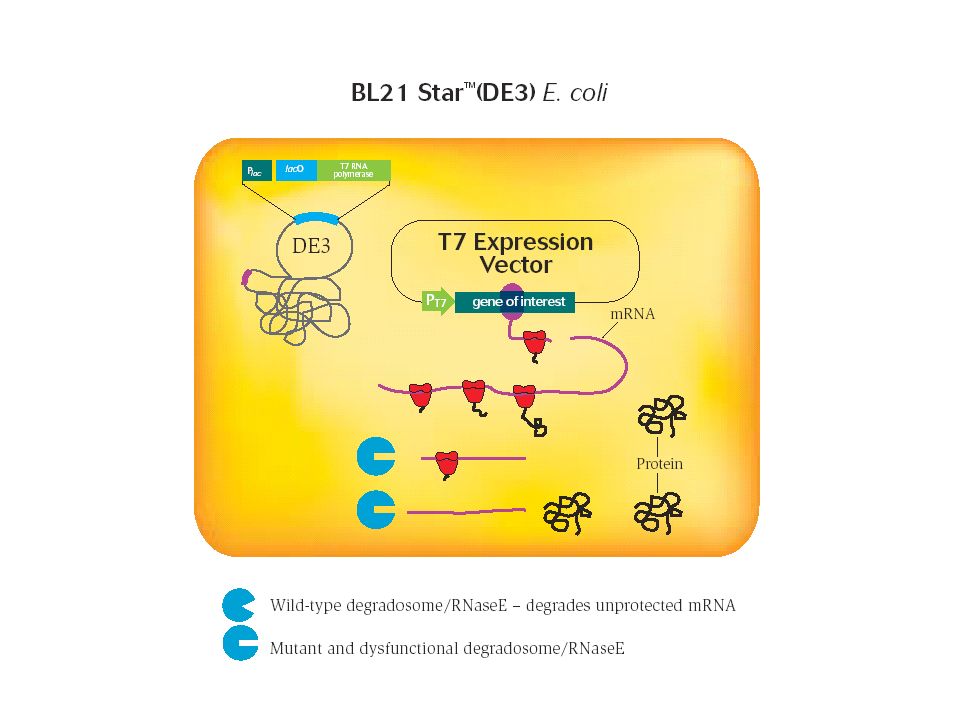
Degradazione dell'mRNA ad opera di RNasi intracellulari
La ribonucleasi E (RNasi E) è un componente chiave del complesso di degradazione dell’RNA E’ stato osservato che in presenza di mutazioni che inattivano la RNasi E della cellula batterica ospite, il livello di espressione di una proteina ricombinante aumenta da 2 a 10 volte E’ stato creato un ceppo di E.coli, denominato BL21 StarTM, caratterizzato da una mutazione del gene codificante la RNasi E
 è un componente chiave del complesso di. degradazione dell’RNA. E’ stato osservato che in presenza di mutazioni che inattivano la RNasi. E della cellula batterica ospite, il livello di espressione di una proteina. ricombinante aumenta da 2 a 10 volte. E’ stato creato un ceppo di E.coli, denominato BL21 StarTM, caratterizzato da una mutazione del gene codificante la RNasi E.")
11
Analisi dell’espressione dell’enzima β-galattosidasi
BL21(DE3) BL21 Star(DE3) U I U I
 BL21 Star(DE3) U I U I.")
12
Il miglioramento della stabilità delle proteine
Il tempo di dimezzamento di diverse proteine varia da qualche minuto a qualche ora L’origine di questa stabilità differenziale risiede sia nella misura in cui si formano i ponti disolfurici sia nella presenza di certi aminoacidi all’estremità N-terminale E’ stato osservato che, aggiungendo certi aminoacidi all’estremità N-terminale della β-galattosidasi, la sopravvivenza in vitro della proteina modificata passa da circa 2 minuti a oltre 20 ore Spesso è sufficiente la presenza di un unico aminoacido in più all’estremità N-terminale per stabilizzare la proteina bersaglio Le proteine con una emivita maggiore possono accumularsi nella cellula aumentando, di conseguenza, la resa in prodotto

13
Analisi della stabilità della β-galattosidasi
Aminoacido Tempo di dimezzamento Met, Ser, Ala, Thr, Val, Gly 20 h Ile, Glu 30’ Tyr, Gln 20’ Pro 10’ Phe, Leu, Asp, Lys 3’ Arg 1’

14
Al contrario degli aminoacidi all’N-terminale della proteina, certe sequenze
intermedie rendono la proteina maggiormente suscettibile alla degradazione proteolitica Queste regioni della proteina, dette sequenze PEST, sono ricche di prolina (P), acido glutammico (E), serina (S) e treonina (T) Tali sequenze sono spesso fiancheggiate da gruppi di aminoacidi a carica positiva, i quali possono agire marcando le proteine ai fini della degradazione all’interno della cellula Alterando le regioni PEST è possibile migliorare la stabilità di una proteina
, acido glutammico (E), serina (S) e treonina (T) Tali sequenze sono spesso fiancheggiate da gruppi di aminoacidi a carica. positiva, i quali possono agire marcando le proteine ai fini della. degradazione all’interno della cellula. Alterando le regioni PEST è possibile migliorare la stabilità di una proteina.")
15
L'impiego di ceppi ospiti carenti di proteasi
Non è semplice, dal momento che E.coli possiede almeno 25 proteasi e solo alcune di esse sono state caratterizzate Si tratta di enzimi coinvolti nella degradazione di proteine difettose o anomale e dunque necessari ad assicurare la continua vitalità delle cellule E’ stato osservato che i ceppi maggiormente carenti crescevano più lentamente → la diminuzione dell’attività proteasica debilita le cellule Nel caso di ceppi di E.coli portatori di mutazioni sia a carico del gene per il fattore sigma della RNA polimerasi responsabile della sintesi della proteina dello shock termico (rpoH) sia a carico del gene per una proteasi necessaria alla crescita cellulare a temperature elevate (degP), le proteine secrete possedevano un’attività specifica volte maggiore di quella delle proteine prodotte da cellule ospiti di tipo naturale Questo aumento apparente di attività riflette la diminuzione della degradazione proteolitica delle proteine secrete
 sia a carico del gene per una proteasi necessaria. alla crescita cellulare a temperature elevate (degP), le proteine secrete. possedevano un’attività specifica 36 volte maggiore di quella delle. proteine prodotte da cellule ospiti di tipo naturale. Questo aumento apparente di attività riflette la diminuzione della degradazione. proteolitica delle proteine secrete.")
16
Il ceppo BL21(DE3) presenta una mutazione del gene codificante la proteasi
Lon Si tratta di una proteasi ATP-dipendente coinvolta nella degradazione di proteine estranee o non correttamente ripiegate Il ceppo RF presenta mutazioni dei geni degP e ompT, le quali determinano l’inattivazione di una proteasi periplasmatica (degP) e di una proteasi della membrana esterna (ompT) L’aumento dei livelli di proteine ricombinanti nei ceppi carenti di proteasi è da ascrivere sia ad una ridotta degradazione allinterno della cellula sia ad una ridotta proteolisi in seguito alla lisi cellulare
 e di una. proteasi della membrana esterna (ompT) L’aumento dei livelli di proteine ricombinanti nei ceppi carenti di proteasi è. da ascrivere sia ad una ridotta degradazione allinterno della cellula sia ad. una ridotta proteolisi in seguito alla lisi cellulare.")
17
Solubilità e localizzazione cellulare
Sono disponibili diverse strategie per esprimere proteine ricombinanti solubili ed attive La solubilità di una determinata proteina è determinata da una varietà di fattori, inclusa la sua specifica sequenza aminoacidica L’ospite batterico, il tipo di vettore di espressione e le condizioni di crescita possono determinare un incremento o una riduzione della quantità di proteina ricombinante in forma solubile o insolubile STRATEGIE PER FAVORIRE IL FOLDING E/O INCREMENTARE LA SOLUBILITA’ DELLA PROTEINA RICOMBINANTE DI INTERESSE: Fusione del prodotto proteico ricombinante ad un polipeptide altamente solubile Fusione ad un enzima che catalizza la formazione di ponti disolfurici Traslocazione della proteina ricombinante nello spazio periplasmico

18
Temperatura La crescita degli ospiti batterici a 37ºC determina l’accumulo di alcune proteine ricombinanti nei corpi di inclusione L’incubazione a 30ºC favorisce la produzione di proteine ricombinanti solubili ed attive La crescita e l’induzione a 25ºC o 30ºC possono risultare ottimali se si desidera la secrezione della proteina ricombinante nello spazio periplasmico, sfruttando la presenza del peptide segnale In alcuni casi un’induzione prolungata (overnight) a basse temperature (15ºC-20ºC) ha consentito di ottenere maggiori rese di proteine ricombinanti solubili
 a basse. temperature (15ºC-20ºC) ha consentito di ottenere maggiori rese di. proteine ricombinanti solubili.")
19
Integrazione della sequenza codificante nel cromosoma batterico
Ridurre il sovraccarico metabolico Eludere il problema del mantenimento del plasmide (tramite antibiotici o altri metaboliti costosi) in scala industriale
 in scala industriale.")
20
Solubilità delle proteine ricombinanti
In generale le condizioni sperimentali che determinano una riduzione dei livelli di espressione, quali induzione a basse temperature o crescita in un terreno minimo, determinano un incremento della percentuale di proteina bersaglio in forma solubile Per numerose applicazioni è necessario esprimere la proteina bersaglio nella sua forma solubile e biologicamente attiva Una proteina solubile non è necessariamente rinaturata in maniera corretta → alcune proteine formano specie solubili inattive Il tipo di vettore, il ceppo ospite, la sequenza della proteina ricombinante e le condizioni di crescita sono tutti fattori che contribuiscono ad incrementare o ridurre la proporzione di proteina in forma solubile o insolubile

21
Secrezione nel periplasma
Sito di taglio per proteasi specifica Rinaturazione in ambiente ossidante leader proteina Traslocazione attraverso la membrana interna N C leader proteina 20-30 aa

22
Secrezione nel periplasma
Una strategia per ottenere proteine solubili ed attive consiste nell’utilizzo di vettori che consentono il trasporto della proteina ricombinante nel periplasma, un ambiente idoneo al folding ed alla formazione di ponti disolfurici A tale scopo sono adoperati vettori di espressione contenenti una sequenza codificante un peptide segnale Nel corso del clonaggio è importante fare in modo che la sequenza codificante la proteina ricombinante di interesse sia in frame con la sequenza codificante il peptide segnale Alcune proteine non sono risultate buoni candidati per la localizzazione periplasmatica La carica netta degli aminoacidi N-terminali della proteina matura può inibire la traslocazione

23
pET-22b(+) peptide segnale sito NcoI
 peptide segnale sito NcoI")
24
Elevati livelli di espressione possono inibire la secrezione sovraccaricando il macchinario secretorio della cellula Strategie • Variare le condizioni sperimentali in modo da ridurre i livelli di espressione • Rifornire la cellula di alcuni dei componenti limitanti del percorso secretorio Sono disponibili ceppi di E.coli trasformati con plasmidi contenenti i geni prlA4 e secE. Tali geni codificano i principali componenti del sistema di trasporto attraverso la membrana. Adoperando tali ceppi si è osservato un effettivo incremento della secrezione di proteine ricombinanti.

25
INCONVENIENTI... Basse rese Può accadere che il peptide segnale non sia rimosso o che il taglio avvenga in un sito inappropriato Talvolta può risultare utile l’inserimento di uno spaziatore aminoacidico tra il sito di taglio del peptide segnale e la proteina ricombinante o l’utilizzo di ceppi di E.coli che esprimono ad elevati livelli uno o più geni codificanti peptidasi specifiche per il peptide segnale

26
PREPARAZIONE DELL'ESTRATTO PERIPLASMICO
Le proteine secrete nello spazio periplasmico di E.coli possono essere isolate mediante shock osmotico Il trattamento dei batteri con soluzioni ipertoniche contenenti EDTA determina una variazione della permeabilità della membrana/parete cellulare La successiva incubazione con una soluzione ipotonica (ad es. MgCl2 0.1 mM) determina la permeabilizzazione della parete cellulare ed il rilascio dei componenti dello spazio periplasmico Le cellule batteriche raccolte in fase esponenziale di crescita sono più suscettibili allo shock osmotico rispetto alle cellule in fase stazionaria Circa il 4% delle proteine totali di E.coli è secreta nello spazio periplasmico Le proteine secrete includono un certo numero di enzimi degradativi L’utilizzo di ceppi di E.coli carenti di proteasi e l’aggiunta di inibitori delle proteasi possono determinare un incremento della resa
 determina la permeabilizzazione della parete cellulare. ed il rilascio dei componenti dello spazio periplasmico. Le cellule batteriche raccolte in fase esponenziale di crescita sono più. suscettibili allo shock osmotico rispetto alle cellule in fase stazionaria. Circa il 4% delle proteine totali di E.coli è secreta nello spazio. periplasmico. Le proteine secrete includono un certo numero di enzimi degradativi. L’utilizzo di ceppi di E.coli carenti di proteasi e l’aggiunta di inibitori. delle proteasi possono determinare un incremento della resa.")
27
TUTTE LE PROTEINE SONO TRASLOCATE IN MANIERA
EFFICIENTE NELLO SPAZIO PERIPLASMICO? Alcune proteine restano legate alla membrana e non possono essere isolate mediante shock osmotico In tali casi può risultare utile il trattamento con detergenti

28
CORPI DI INCLUSIONE Elevati livelli di espressione di proteine ricombinanti in E.coli spesso determinano la formazione di granuli citoplasmatici, i corpi di inclusione, composti da aggregati insolubili della proteina espressa Ai fini della preparazione dei corpi di inclusione, le cellule, raccolte mediante centrifugazione, sono lisate con metodi meccanici, quali sonicazione o lisozima e detergenti I corpi di inclusione, raccolti mediante centrifugazione, sono lavati (Triton X EDTA o urea) allo scopo di separare il più possibile proteine solubili dalle proteine ricombinanti aggregate Nella maggior parte dei casi sono isolati corpi di inclusione contenenti più del 90% di proteina ricombinante pura
 allo scopo di separare il più possibile. proteine solubili dalle proteine ricombinanti aggregate. Nella maggior parte dei casi sono isolati corpi di inclusione. contenenti più del 90% di proteina ricombinante pura.")
29
ISOLAMENTO DELLA PROTEINA RICOMBINANTE
I corpi di inclusione lavati sono solubilizzati e la proteina ricombinante è sottoposta ad una procedura di rinaturazione Varie condizioni possono essere adoperate per solubilizzare i corpi di inclusione → guanidina-HCl, urea, pH basico, SDS, acetonitrile/propanolo Indipendentemente dalle procedure è importante lisare le cellule in maniera efficiente per ottenere i corpi di inclusione in grandi quantità Le procedure di rinaturazione variano a seconda della proteina e devono essere determinate in maniera empirica In generale la rinaturazione è realizzata mediante rimozione graduale dell’agente denaturante

30
Denaturazione riduttiva e rinaturazione ossidativa della RNasi A
DENATURAZIONE E RINATURAZIONE DELLA RNasi A Ossidazione O2 a pH 8 (condizioni rinaturanti) Riduzione con b-mercaptoetanolo (condizioni denaturanti) Molecola denaturata ridotta 1 Denaturazione riduttiva e rinaturazione ossidativa della RNasi A
 Riduzione con b-mercaptoetanolo (condizioni denaturanti) Molecola denaturata ridotta. 1. Denaturazione riduttiva e rinaturazione ossidativa della RNasi A.")
31
Allontanamento dei denaturanti per DIALISI
GSH 5 mM GSSG 1 mM 5:1 pH 8.4

32
UTILIZZO DI E.coli VANTAGGI SVANTAGGI Crescita rapida
Elevata densità cellulare su substrati poco costosi Organismo ben caratterizzato, con una ricca collezione di mutanti Numero elevato di vettori di clonaggio e di espressione SVANTAGGI Formazione di corpi di inclusione Rinaturazione ossidativa limitata allo spazio periplasmico Modifiche post-traduzionali non adeguate (ad es. glicosilazione)
")
33
Bacillus subtilis è un batterio gram-positivo adoperato
per la produzione di proteine ricombinanti

34
Strumento utile per lo studio di complessi proteici
che giocano un ruolo chiave nei processi cellulari Si tratta di vettori che consentono il clonaggio e l’espressione di due diversi geni bersaglio Contengono due unità di espressione, ciascuna regolata da un promotore T7lac distinto

35
Terreni di coltura L’utilizzo di diversi terreni di
coltura spesso determina differenze notevoli nei livelli di espressione Per definire i parametri generali dell’espressione è di solito adoperato il terreno LB Si può poi procedere ad effettuare delle prove utilizzando altri terreni

36
Resistenza all'antibiotico
Le colture batteriche tendono a perdere la resistenza all’antibiotico ampicillina, dal momento che gli elevati livelli di enzima β-lattamasi secreto e la riduzione del pH del mezzo di coltura associata alla fermentazione determinano la degradazione dell’antibiotico Si può sostituire il mezzo di coltura con un mezzo di coltura fresco contenente ampicillina o si può utilizzare l’antibiotico correlato carbenicillina, il quale è meno sensibile a bassi valori di pH Nella maggior parte dei vettori pET il gene codificante la β-lattamasi è localizzato a valle del promotore T7, nello stesso orientamento → l’enzima si accumula nelle colture di cellule batteriche indotte L’orientamento del gene codificante la β-lattamasi è stato invertito nei vettori pET- 43 e pET-44 → la trascrizione guidata dall’RNA polimerasi T7 non determina l’aumento dei livelli dell’enzima β-lattamasi

37
SCARSITA' DI OSSIGENO CONSEGUENZE RIMEDI
L’ossigeno si discioglie solo in misura limitata nei mezzi acquosi Con l’aumento della densità cellulare si verifica una riduzione della percentuale di ossigeno disciolto nel mezzo di coltura Immettere nel mezzo di coltura grandi quantità di ossigeno o applicare un’agitazione vigorosa non sempre risolve il problema → l’ossigeno si discioglie molto lentamente CONSEGUENZE In condizioni di scarsità di ossigeno, la crescita esponenziale rallenta e la coltura entra presto in fase stazionaria Una conseguenza è la produzione, da parte delle cellule ospiti, di proteasi RIMEDI Ottimizzare l’areazione e l’agitazione Aggiungere al mezzo sostanze chimiche che aumentino la solubilità dell’ossigeno

38
Concentrazione di induttore
L’aggiunta di IPTG ad una concentrazione di 0,4 mM è raccomandata nel caso dei sistemi pET Per i promotori lac o T7lac è consigliata una concentrazione di IPTG pari ad 1 mM La concentrazione ottimale di IPTG è determinata in maniera empirica → variare la concentrazione di IPTG tra 0.01 e 5 mM In alcuni casi è importante indurre la trascrizione lentamente (basse concentrazioni di IPTG) per non caricare eccessivamente il macchinario biosintetico del batterio Un parametro di fondamentale importanza è la temperatura alla quale sono fatti crescere i batteri prima e durante l’induzione
 per non caricare eccessivamente il macchinario. biosintetico del batterio. Un parametro di fondamentale importanza è la temperatura alla quale. sono fatti crescere i batteri prima e durante l’induzione.")
39
Sistemi su larga scala Nei recipienti di coltura piccoli (da 1 a 5 litri), l’induzione si realizza facilmente variando la temperatura o aggiungendo un induttore chimico Nei bioreattori di dimensione industriale la variazione di temperatura richiede tempo (da 30 a 60 minuti) ed energia In maniera simile l’aggiunta di un induttore chimico (IPTG) può rendere il processo molto costoso, inoltre l’aggiunta alle vasche di coltura/fermentatori riciede tempo e crea disomogeneità Si può adoperare un sistema a due plasmidi → il repressore cI è posto sotto il controllo del promotore trp in un plasmide a basso numero di copie mentre il gene di interesse è posto sotto il controllo del promotore pL In assenza di triptofano il promotore trp si attiva ed è sintetizzata la proteina repressore che inattiva il promotore pL In presenza di triptofano è disattivato il promotore trp con l’arresto della sintesi della proteina repressore cI → il promotore pL acquista piena attività
 ed energia. In maniera simile l’aggiunta di un induttore chimico (IPTG) può. rendere il processo molto costoso, inoltre l’aggiunta alle vasche di. coltura/fermentatori riciede tempo e crea disomogeneità. Si può adoperare un sistema a due plasmidi → il repressore cI è. posto sotto il controllo del promotore trp in un plasmide a basso. numero di copie mentre il gene di interesse è posto sotto il. controllo del promotore pL. In assenza di triptofano il promotore trp si attiva ed è sintetizzata. la proteina repressore che inattiva il promotore pL. In presenza di triptofano è disattivato il promotore trp con l’arresto. della sintesi della proteina repressore cI → il promotore pL. acquista piena attività.")
40
Le cellule possono essere coltivate in un terreno non costoso contenente
piccolissime quantità di triptofano libero. L’induzione è poi realizzata aggiungendo al mezzo il triptone che contiene abbastanza triptofano libero da indurre la trascrizione in maniera efficiente.

Presentazioni simili


















>")
