Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
La fosforilazione ossidativa è il processo in cui viene formato ATP, mentre gli elettroni sono trasferiti dal NADH oppure dal FADH2 all’ossigeno, attraverso una serie di trasportatori di elettroni Nelle forme di vita superiore le sostanze alimentari, rappresentate principalmente da carboidrati e lipidi, sono metabolizzate a biossido di carbonio e ad acqua L’energia fornita da questa catena di reazioni ossidative è conservata sotto forma di un composto fosforilato ad alta energia, l’adenosina trifosfato (ATP)
")
2
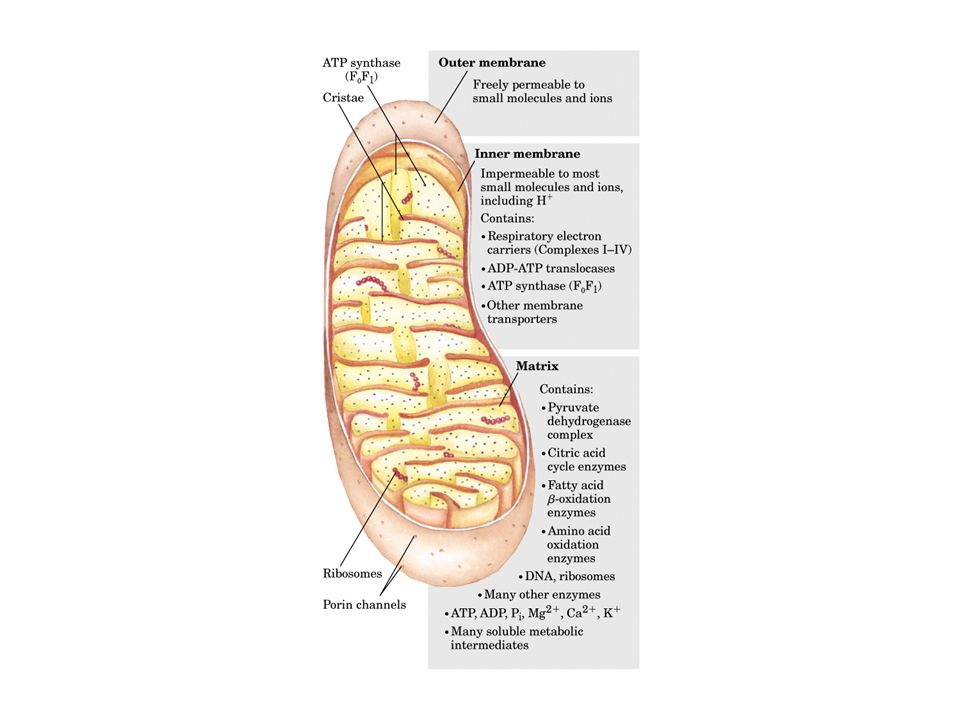
L’ATP è composto da una base purinica, l’adenina, uno zucchero a cinque atomi di carbonio, il ribosio, e dai gruppi fosforici , , e La fosforilazione ossidativa ha luogo su strutture respiratorie localizzate nella membrana interna dei mitocondri I mitocondri sono organelli di forma ovoidale costituiti da due sistemi di membrane: una membrana esterna ed una membrana interna molto estesa e ripiegata

4
La membrana esterna del mitocondrio è permeabile alla maggior parte delle molecole piccole ed agli ioni, in quanto contiene molte copie della proteina porina, una proteina transmembrana che forma un poro La membrana interna è impermeabile a quasi tutti gli ioni ed alle molecole polari. La membrana interna è attraversata da specifiche proteine trasportatrici, come quella per l’ADP e per gli acidi grassi a lunga catena

5
Nei mitocondri esistono due compartimenti: lo spazio intermembrana, tra la membrana interna e quella esterna, e la matrice, lo spazio delimitato dalla membrana interna La fosforilazione ossidativa ha luogo nella membrana mitocondriale interna, mentre la maggior parte delle reazioni del ciclo dell’acido citrico e della via di ossidazione degli acidi grassi avvengono nella matrice

6
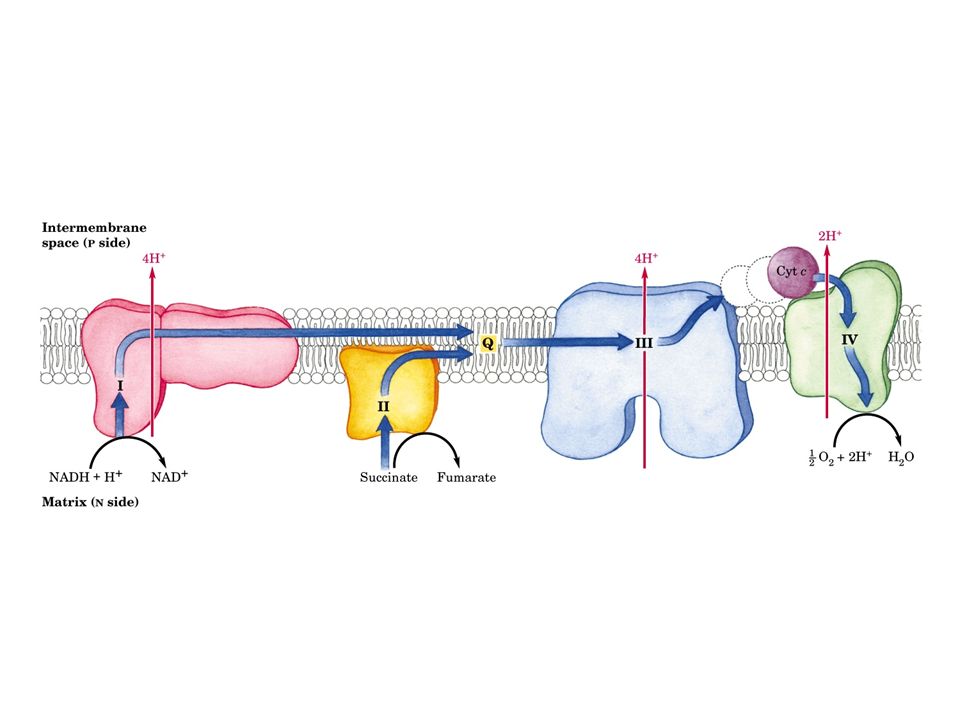
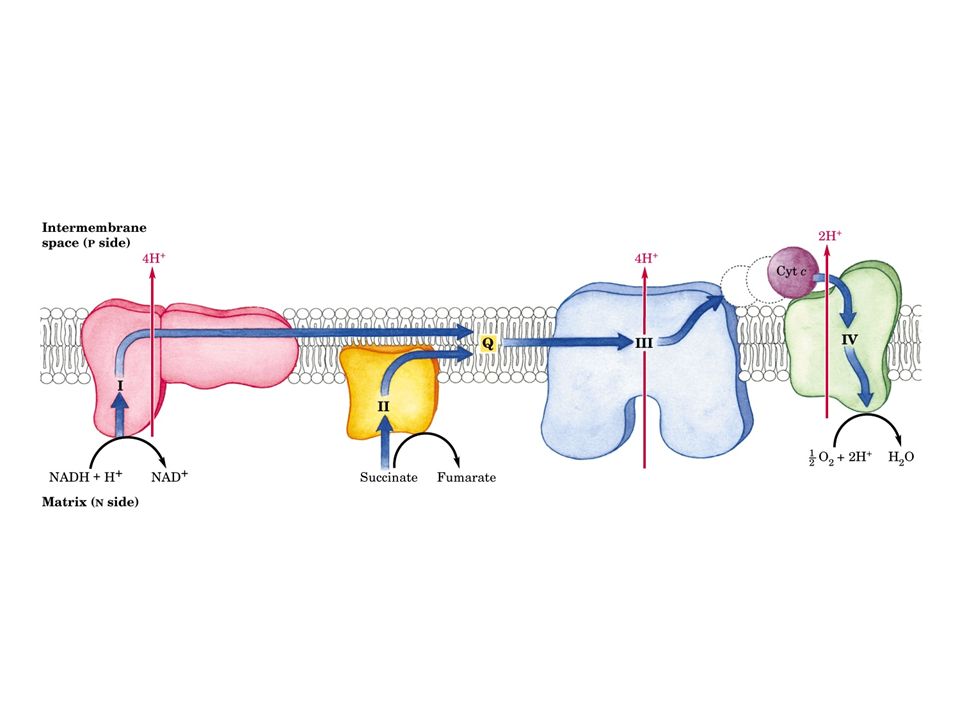
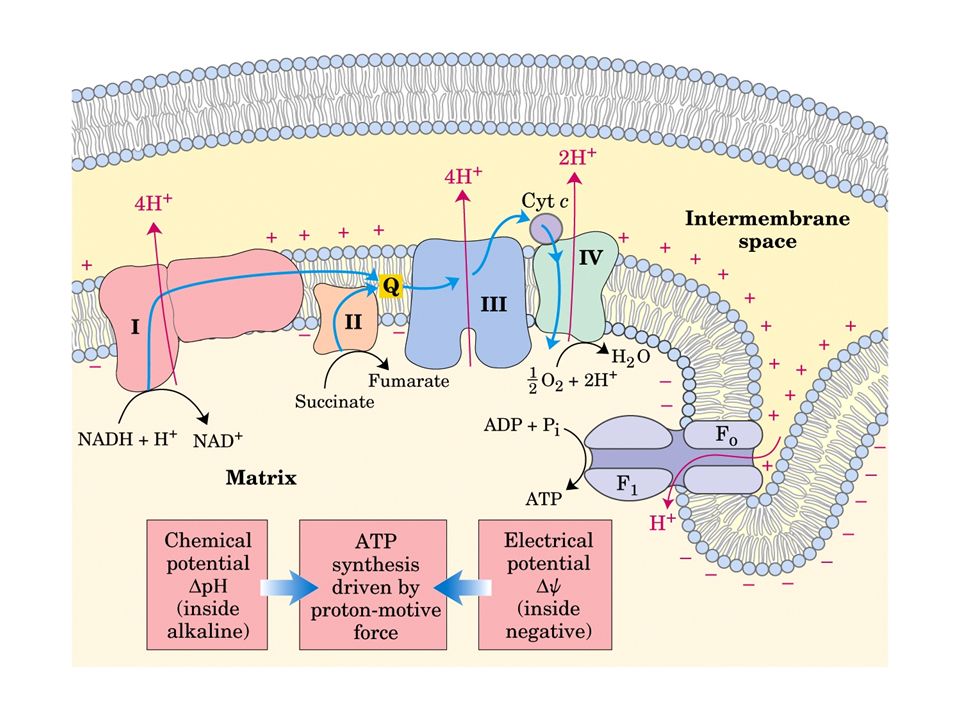
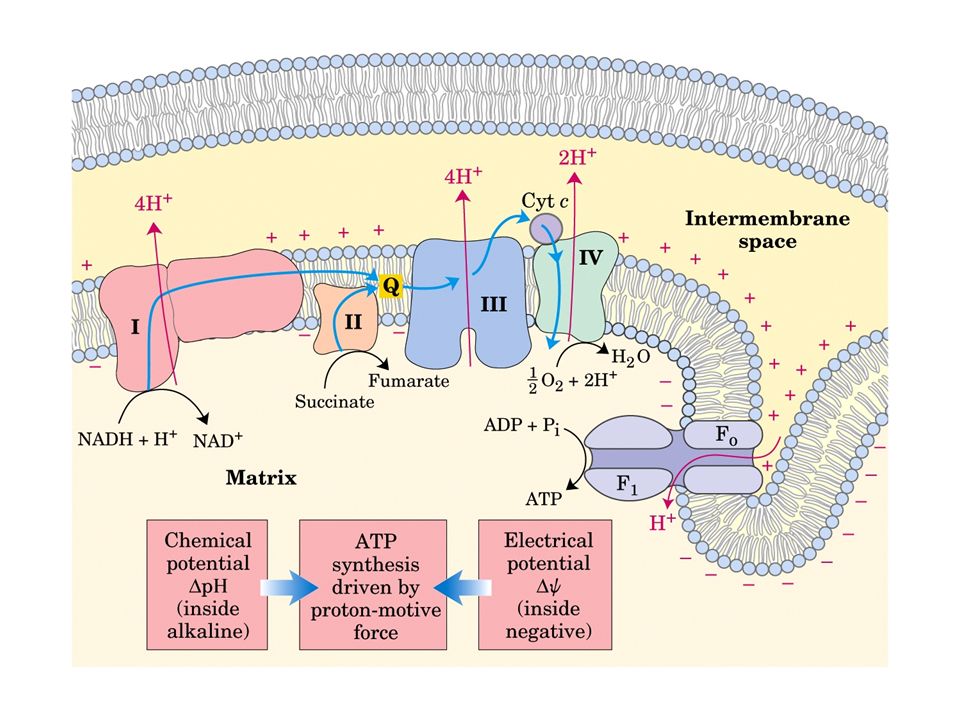
La catena di trasporto degli elettroni o catena respiratoria, è localizzata nella membrana mitocondriale interna, e consiste di diversi grandi complessi proteici e di due piccole componenti indipendenti: l’ubichinone e il citocromo c Gli elettroni attraversano questo sistema in una sequenza che li porta dai coenzimi ridotti all’ossigeno, e lungo la quale i cambiamenti di energia libera alimentano il trasporto di protoni dalla matrice allo spazio intermembrana

7
Vi sono quattro siti d’ingresso per gli elettroni nella catena respiratoria: uno per il NADH (Complesso I) e tre per il FADH2 (Complesso II). Queste vie confluiscono a livello della piccola molecola lipofila dell’ubichinone (coenzima Q10), all’inizio della porzione comune della catena di trasporto, che consiste nel Complesso III, del citocromo c e del Complesso IV L’accettore finale degli elettroni, al termine della catena, è l’ossigeno molecolare che viene ridotto ad acqua
, all’inizio della porzione comune della catena di trasporto, che consiste nel Complesso III, del citocromo c e del Complesso IV L’accettore finale degli elettroni, al termine della catena, è l’ossigeno molecolare che viene ridotto ad acqua.")
9
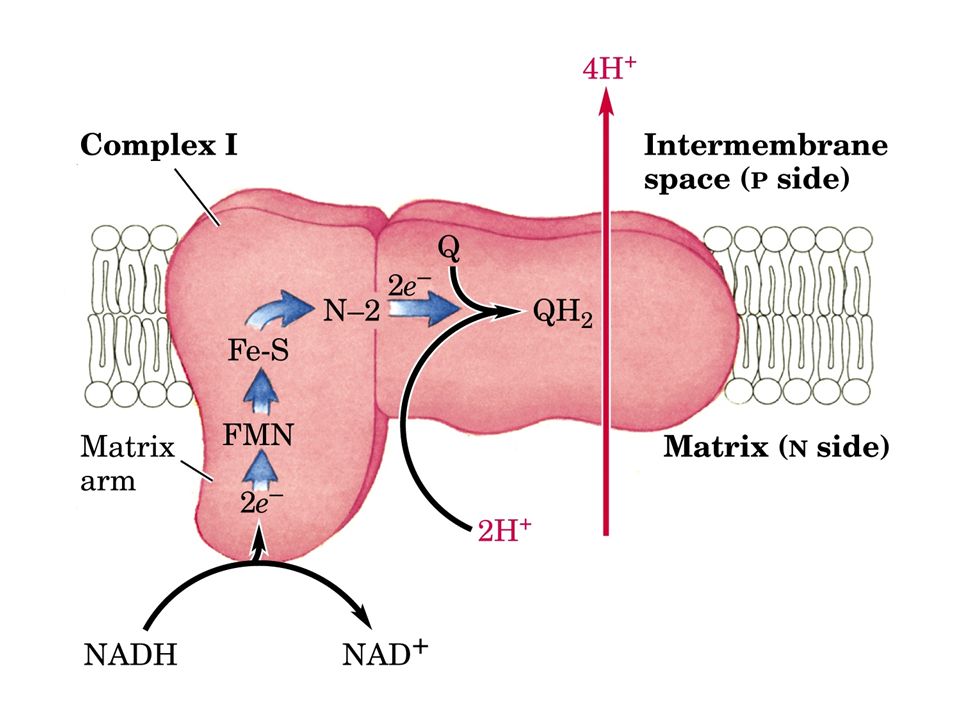
Il Complesso I, è una flavoproteina contenente FMN. Il Complesso I è responsabile dell’ossidazione del NADH mitocondriale e del trasferimento degli elettroni, attraverso il FMN e i complessi ferro-zolfo, all’ubichinone, fornendo cosi energia sufficiente, sotto forma di gradiente di protoni, per la sintesi di una mole di ATP

11
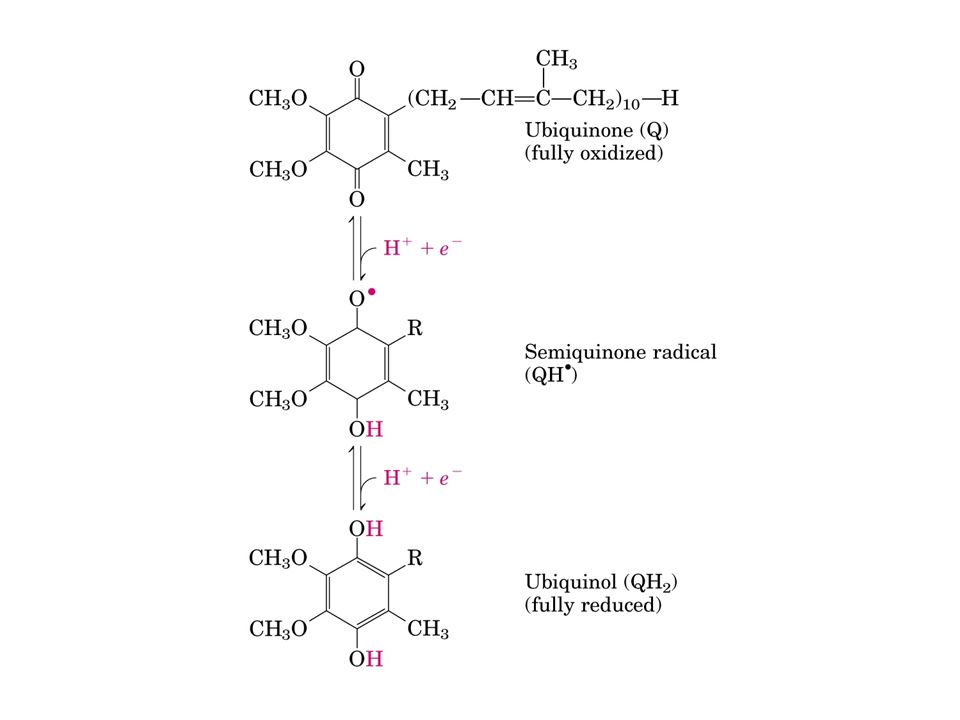
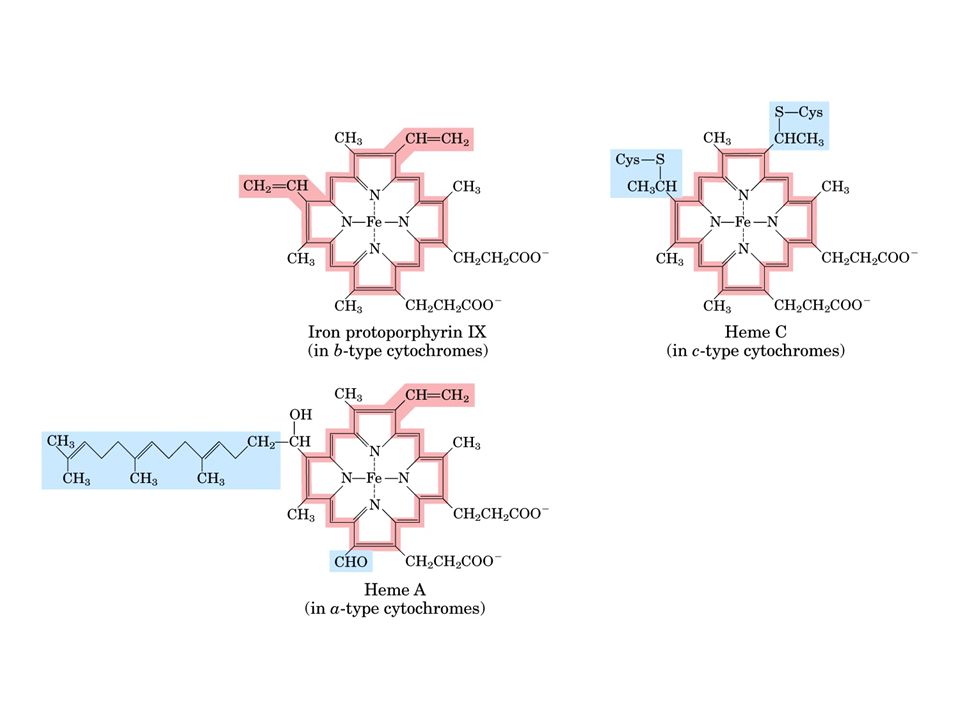
L’ubichinone che si trova in tutti i sistemi viventi, è una piccola molecola liposolubile localizzata nella membrana interna dei mitocondri di cellule vegetali ed animali L’ubichinone delle cellule dei mammiferi contiene una coda idrocarburica di 10 unità isopreniche, e viene perciò definito CoQ 10. E’ in grado di diffondere lungo la membrana mitocondriale interna dove accetta elettroni dalle flavoproteine e li trasferisce al Complesso III (QH 2 - citocromo c reduttasi)
.")
13
L’ubichinone incanala il flusso di elettroni provenienti dalle flavoproteine dei Complessi I e II verso il Complesso III, un oligomero di circa otto peptidi contenente il citocromo b, un centro ferro- zolfo e il citocromo c 1 Questo complesso enzimatico (ubichinone- citocromo c reduttasi) ossida l’ubichinone e riduce il citocromo c
 ossida l’ubichinone e riduce il citocromo c")
15
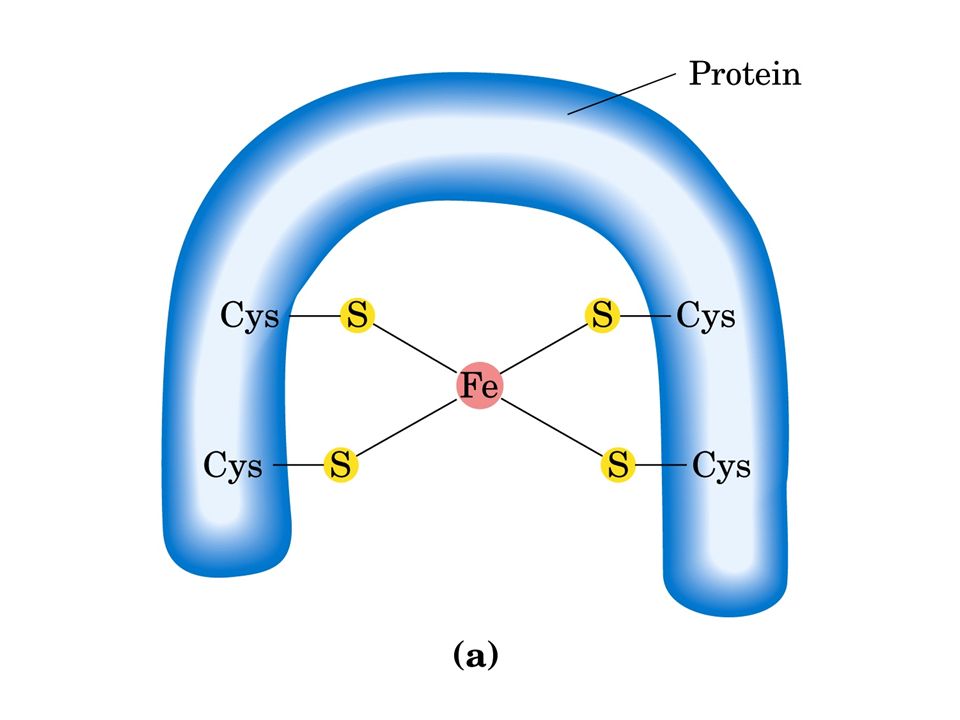
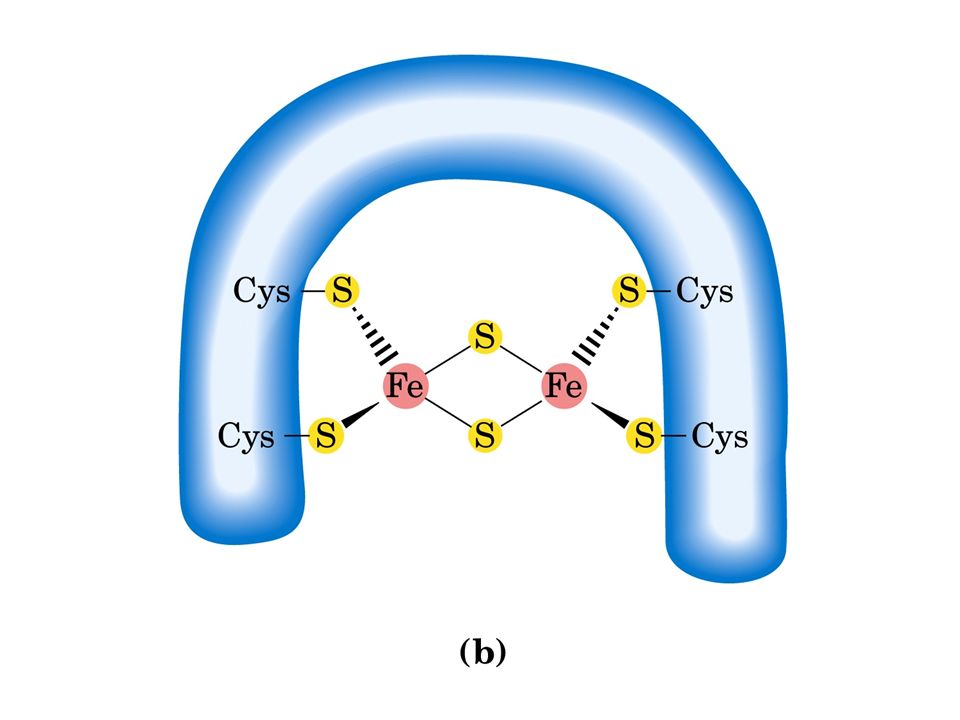
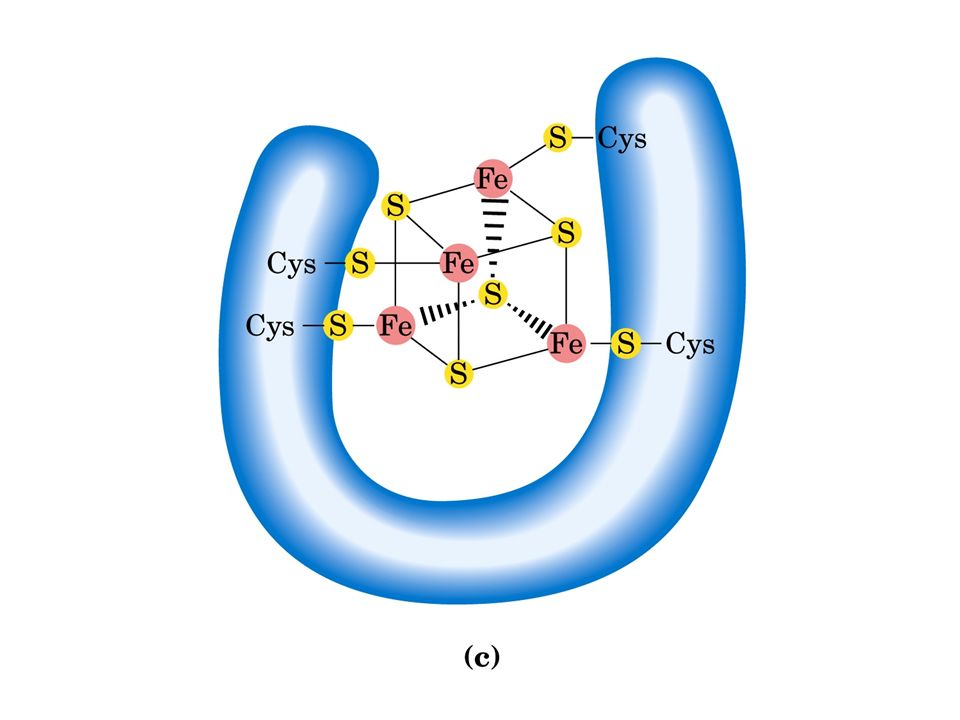
Esistono diversi tipi di centri ferro-zolfo: Fe 2 S 2 e Fe 4 S 4. In ciascun tipo il centro ferro-zolfo è legato a residui di cisteina della catena polipeptidica

19
Il citocromo c è una piccola proteina contenente eme, che si trova associata debolmente alla superficie esterna della membrana mitocondriale interna e trasporta elettroni dal complesso III al complesso IV La riduzione, ad opera del citocromo c 1, del ferricitocromo c (Fe 3+ ) a ferrocitocromo c (Fe 2+ ), induce un cambiamento nella conformazione tridimensionale di quest’ultimo, che favorisce il trasferimento di elettroni al citocromo a del complesso IV
 a ferrocitocromo c (Fe 2+ ), induce un cambiamento nella conformazione tridimensionale di quest’ultimo, che favorisce il trasferimento di elettroni al citocromo a del complesso IV")
21
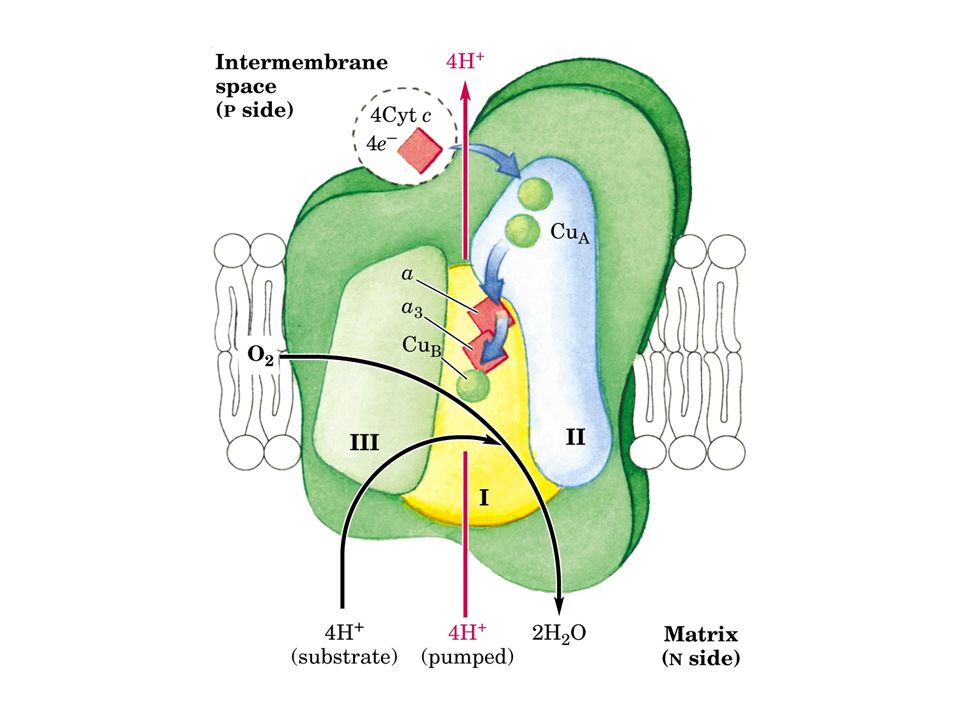
Il complesso IV, noto come citocromo c ossidasi, ossida il citocromo c e trasferisce gli elettroni, attraverso i citocromi a e a3, all’ossigeno che viene ridotto. Il rame è un componente comune a questo enzima ed ad altre ossidasi, alcuni veleni come l’azide ed il cianuro reagiscono con il rame ed inibiscono questo enzima. Come per i complessi I e IIII, il complesso citocromo ossidasi pompa protoni all’esterno del mitocondrio fornendo energia libera per la sintesi di ATP

23
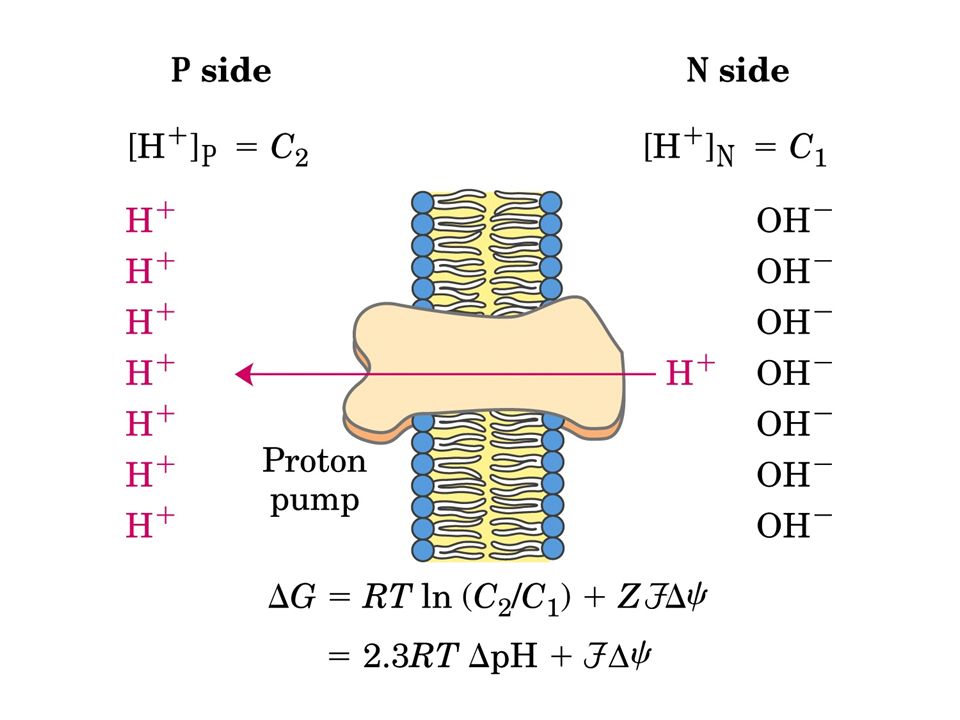
Durante il trasporto di elettroni, i protoni sono pompati dalla matrice nello spazio intermembrana, creando un potenziale elettrochimico attraverso la membrana interna. L’estero del mitocondrio diventa così più acido e più carico positivamente rispetto alla matrice La sintesi di ATP, una reazione non spontanea, è sostenuta dal ritorno dei protoni, in favore del gradiente elettrochimico, nella matrice attraverso l’ATP-sintasi

25
Il mitocondrio assomiglia quindi ad unna “batteria” ad ATP in cui l’energia per la sintesi di ATP è immagazzinata sotto forma di gradiente di protoni. Per funzionare, richiede l’integrità del sistema di membrane interne, che devono essere impermeabili ai protoni tranne che in corrispondenza del complesso ATP- sintasi.

27
In condizioni normali, i processi di ossidazione e fosforilazione sono strettamente accoppiati: i substrati sono ossidati, gli elettroni trasportati, e l’ossigeno consumato solo quando c’è richiesta di sintesi di ATP Perciò nei mitocondri a riposo, la velocità di consumo di ossigeno è bassa. Questa può essere fortemente aumentata dall’aggiunta di ADP I mitocondri sono definiti “disaccoppiati” quando la membrana mitocondriale interna perde la sua integrità strutturale: in questa circostanza i protoni possono diffondere attraverso la membrana lungo vie improduttive

28
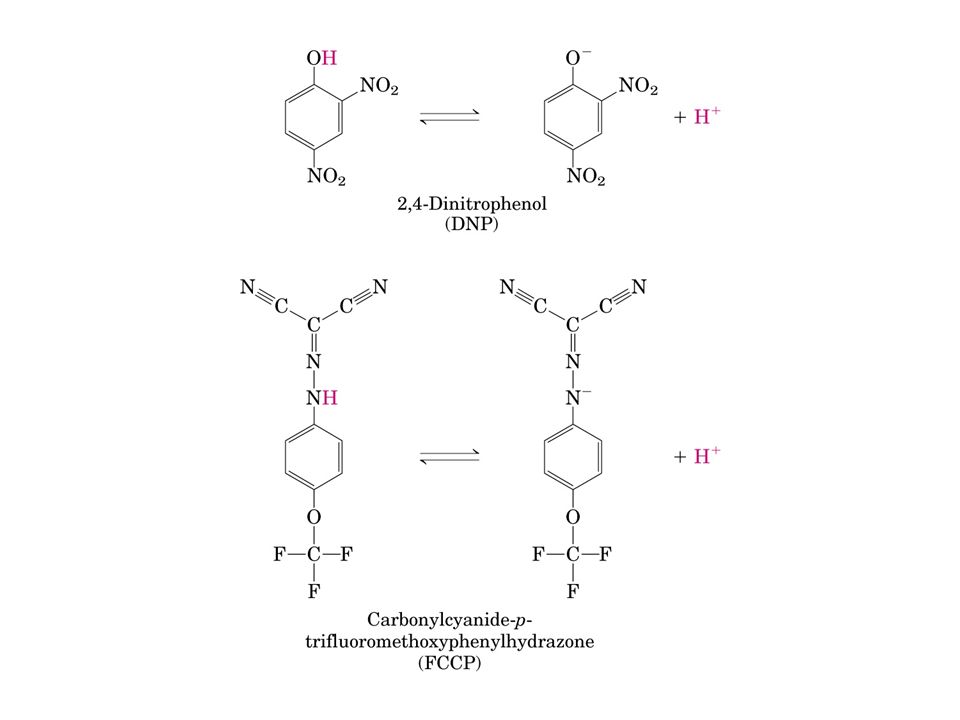
I disaccoppianti della fosforilazione ossidativa dissipano il gradiente di protoni favorendone il reflusso nella matrice mitocondriale scavalcando il sistema dell’ATP-sintasi Un disaccoppiante tipico è il 2,4-dinitrofenolo (DNP), altri comuni disaccopianti sono sostanze conservanti ed antimicrobiche come il pentaclorofenolo e il p- cresolo
, altri comuni disaccopianti sono sostanze conservanti ed antimicrobiche come il pentaclorofenolo e il p- cresolo")
30
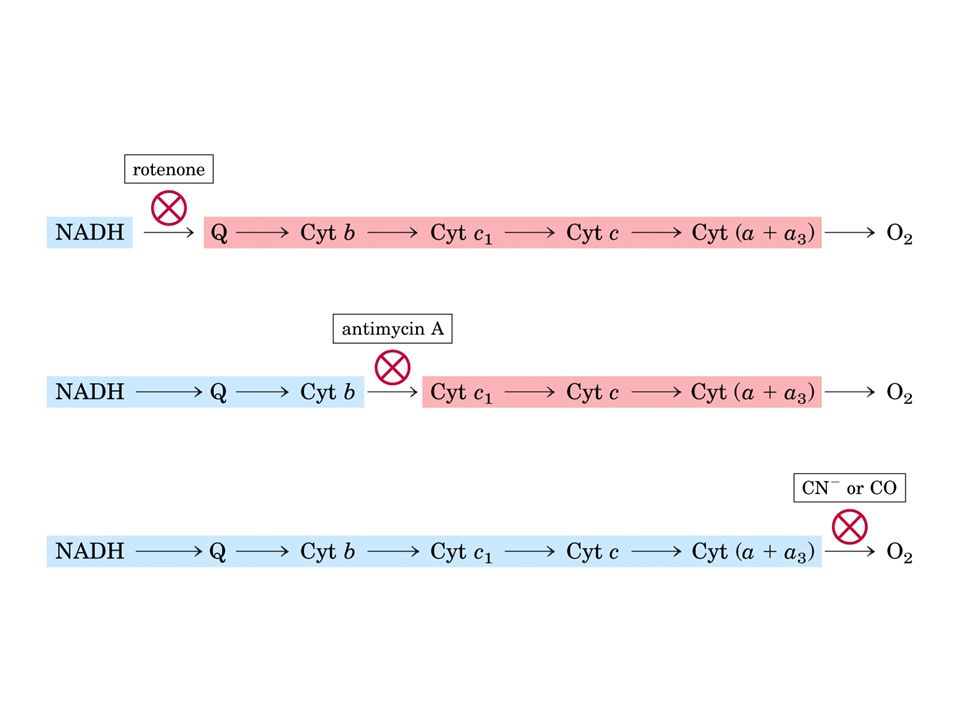
Gli inibitori del trasporto degli elettroni interrompono il flusso di elettroni lungo la catena respiratoria agendo selettivamente sui complessi I, III o IV Il risultato è sempre il blocco delle pompe protoniche, della sintesi di ATP e del consumo di ossigeno Molti di questi inibitori sono veleni, quali ad esempio il rotenone che è un insetticida che inibisce il complesso I del sistema di trasporto degli elettroni

32
Poiché il malato e il lattato sono ossidati dal NAD, il rotenone inibirà la loro ossidazione L’antimicina A, inibendo il complesso III, impedisce il trasferimento di elettroni dai complessi I o II al citocromo c Sia il cianuro che il monossido di carbonio si legano, inibendolo, al complesso IV

34
Le cellule rispondono all’avvelenamento da cianuro o da monossido di carbonio passando ad un metabolismo anaerobico, che causa acidosi e da ultimo morte, se non si prendono immediate contromisure. L’avvelenamento da monossido di carbonio può essere trattato con la somministrazione di ossigeno

Presentazioni simili


















>")
