Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
L’UNITÁ FOTOSINTETICA E IL CICLO FOTOCHIMICO NEI BATTERI ROSSI Struttura e funzionalità dei complessi molecolari coinvolti nella conversione primaria dell’energia luminosa

2
Negli organismi fotosintetici la conversione dell’energia è possibile grazie ad un macchinario supramolecolare costituito da molecole e complessi molecolari differenti opportunamente organizzati in vivo ADP + P ATP h Sistema supramolecolare sede degli eventi primari della fotosintesi

3
I“pezzi” del macchinario supramolecolare I “pezzi” del macchinario supramolecolare (Rb. sphaeroides) membrana biologica complessi antenna (Light Harvesting) centro di reazione (RC) ubichinolo:citocromo c ossidoreduttasi (bc 1 complex) ATP-ase citocromo c 2 ubichinone/ubichinolo
 membrana biologica complessi antenna (Light Harvesting) centro di reazione (RC) ubichinolo:citocromo c ossidoreduttasi (bc 1 complex) ATP-ase citocromo c 2 ubichinone/ubichinolo.")
4
Morfologia di R. sphaeroides Parete cellulare Membrana plasmatica con invaginazioni periplasma citoplasmaSacca di poliidrossibutirrati

5
Organizzazione dei componenti LH ICM periplasma citoplasma

6
Complessi Antenna + Centro di Reazione UNITÁ FOTOSINTETICA Raccolta e conversione primaria dell’energia elettromagnetica

7
Complessi Antenna (Light Harvesting)
")
8
LH2 complex

9
LH II

10
Supramolecolar organization

11
Efficient exciton funnelling in PSU

12
Meccanismi di trasferimento dell’eccitazione: FORSTER DEXTER Meccanismi di trasferimento dell’eccitazione: FORSTER DEXTER ASSORBIMENTO FOTONI TRASFERIMENTO ECCITONI

13
REAZIONE FOTODINAMICA Chl hν 1 Chl* 1 Chl* 3 Chl* 3 Chl* + 3 O 2 Chl + 1 O 2 * 3 Chl* + Car Chl + 3 Car* 3 Car* Car + calore RC: LH I = 1:1 RC: LH II = f (T, P) EFFETTO FOTOPROTETTIVO DEI CAROTENOIDI
 EFFETTO FOTOPROTETTIVO DEI CAROTENOIDI")
14
CENTRO DI REAZIONE

15
Spettro ottico del RC

16
Banda “extra” dovuta all’accoppiamento eccitonico delle batterioclorofille del dimero

17
Risposta alla luce del RC

18
Setup sperimentale per la misura di velocità di ricombinazione di carica in centri di reazione isolati

19
Un lampo di luce genera lo stato D + Q B - ; successivamente si assiste al ripristino della forma neutra DQ B secondo una cinetica del primo ordine: Integrando questa espressione fra 0 e t, ponendo = 1/k ed osservando che D + Q B - A, possiamo scrivere:

20
La cinetica di ricombinazione di carica da Q A segue un andamento esponenziale (traccia a); la presenza del chinone nella tasca Q B determina la comparsa di una “fase lenta” ovvero di una seconda componente esponenziale con costante di velocità più piccola (tracce b e c a grado di occupazione di Q B crescente)
; la presenza del chinone nella tasca Q B determina la comparsa di una fase lenta ovvero di una seconda componente esponenziale con costante di velocità più piccola (tracce b e c a grado di occupazione di Q B crescente)")
21
Conversione dell’energia nel Centro di Reazione

22
Trasferimenti elettronici nel RC Notiamo che al procedere della separazione di carica le reazioni di ricombinazione diventano sempre più lente

23
Energetica della separazione di carica G = nF E = nF[E°’(Q/Q ) E°’(P + /P)] E°’(Q/Q ) = +100 mV E°’(P + /P)] = +450 mV G° = +33.8 kJmol Reazione non spontanea
![Energetica della separazione di carica G = nF E = nF[E°’(Q/Q ) E°’(P + /P)] E°’(Q/Q ) = +100 mV E°’(P + /P)] = +450 mV G° = kJmol Reazione non spontanea](http://images.slideplayer.it/33/10538779/slides/slide_23.jpg "Energetica della separazione di carica G = nF E = nF[E°’(Q/Q ) E°’(P + /P)] E°’(Q/Q ) = +100 mV E°’(P + /P)] = +450 mV G° = kJmol Reazione non spontanea")
24
Effetto dell’assorbimento di radiazione E°’(P + /P*) = E°’(P + /P) + E(P-P*) = (450 - 1430) mV = 980 mV Shift di potenziale di riduzione: Assumendo trascurabili il cambiamento di volume ed entropia tra i due stati posso scrivere, essendo a P e T costanti dG = dU + pdV– TdS, G = U
 = E°’(P + /P) + E(P-P*) = ( ) mV = 980 mV Shift di potenziale di riduzione: Assumendo trascurabili il cambiamento di volume ed entropia tra i due stati posso scrivere, essendo a P e T costanti dG = dU + pdV– TdS, G = U")
25
Ricalcoliamo il G G° = nF E°’ = nF[E°’(Q/Q ) E°’(P + /P*)] E°’(Q/Q ) = +100 mV E°’(P + /P*)] = 980 mV G°’ = 84.9 kJmol Reazione spontanea Efficienza di conversione = 1 85/138 = 0.38 Energia scaricata = 62%
![Ricalcoliamo il G G° = nF E°’ = nF[E°’(Q/Q ) E°’(P + /P*)] E°’(Q/Q ) = +100 mV E°’(P + /P*)] = 980 mV G°’ = 84.9 kJmol Reazione spontanea Efficienza di conversione = 1 85/138 = 0.38 Energia scaricata = 62%](http://images.slideplayer.it/33/10538779/slides/slide_25.jpg "Ricalcoliamo il G G° = nF E°’ = nF[E°’(Q/Q ) E°’(P + /P*)] E°’(Q/Q ) = +100 mV E°’(P + /P*)] = 980 mV G°’ = 84.9 kJmol Reazione spontanea Efficienza di conversione = 1 85/138 = 0.38 Energia scaricata = 62%")
26
Il fotociclo del RC Il risultato netto è l’ossidazione di due citocromi c2, la riduzione di un chinone e il sequestramento di due protoni nella membrana

27
La chiusura del fotociclo UQH 2 + 2cyt c 2 (Fe 3+ )+ 2H + (citopl.) UQ + 2cyt c 2 (Fe 2+ ) + 4H + (peripl.) Tale reazione ripristina i reagenti iniziali chiudendo così il ciclo fotosintetico. Vengono inoltre sequestrati altri due protoni dal citoplasma e rilasciati quattro protoni nel periplasma. Il catalizzatore è il bc1 e il G è negativo, cioè la reazione è spontanea.

28
Efficienza quantica di separazione di carica E’ stato misurato che per ogni fotone assorbito dal dimero, si forma uno stato a cariche separate. La resa quantica del processo è quindi del 100%. Tale resa è il risultato dell’ottimale arrangiamento spaziale dei cofattori, e dei loro potenziali redox, per cui le reazioni di trasferimento elettronico “in avanti” sono sempre almeno due ordini di grandezza più veloci delle ricombinazioni. Poiché tutte le reazioni di trasferimento elettronico sono temperatura-indipendenti si è concluso che non hanno energia di attivazione. Questo garantisce che la velocità di reazione sia la massima possibile.

29
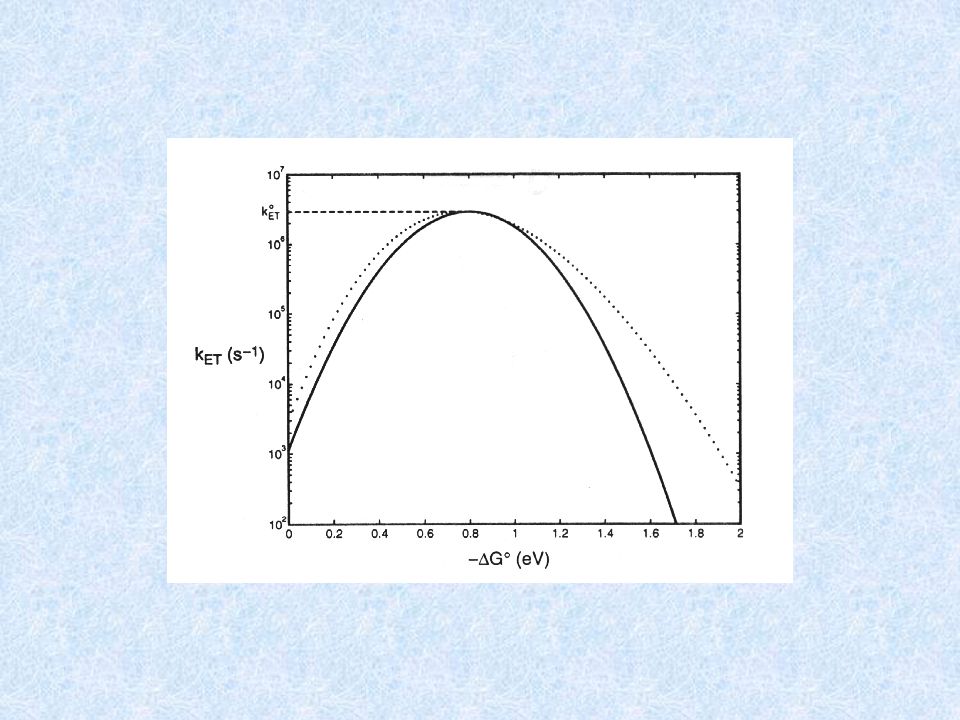
Teoria di Marcus k è massima se G ATT = 0 Tale relazione è verificata se G° = = energia di riorganizzazione a seguito della reazione Per valori di G° più piccoli o più grandi, la reazione rallenta G ATT ( G°- ) 2
 2")
30
Trasferimento di elettroni in sistemi omogenei Pre-equilibrio di formazione del complesso donatore- accettore Trasferimento di un elettrone Scissione del complesso e diffusione degli ioni componenti

31
Dalla teoria del complesso attivato sappiamo che: Dove: è il coefficiente di trasmissione è la frequenza vibrazionale con la quale il complesso attivato si avvicina allo stato di transizione ‡ G è l’energia di Gibbs di attivazione

32
Teoria dei processi di trasferimento elettronico Teoria di Marcus 1)Gli elettroni attraversano per effetto tunnel una barriera di energia di potenziale che dipende dall’energia di ionizzazione dei complessi DA e D+A-; Quantità influenzata: 2)Il complesso DA e le molecole del solvente che lo attorniano devono subire modificazioni strutturali prima che avvenga il trasferimento di elettroni Quantità influenzata: ‡G
Gli elettroni attraversano per effetto tunnel una barriera di energia di potenziale che dipende dall’energia di ionizzazione dei complessi DA e D+A-; Quantità influenzata: 2)Il complesso DA e le molecole del solvente che lo attorniano devono subire modificazioni strutturali prima che avvenga il trasferimento di elettroni Quantità influenzata: ‡G")
33
L’attraversamento in tunnel Principio di Frank-Condon: Le transizioni (e i trasferimenti) elettronici avvengono entro una intelaiatura nucleare stazionaria La coordinata della dislocazione corrisponde al cambiamento della geometria, ovvero un modo di insieme di datore, accettore e solvente ‡G‡G G° DA D+AD+A Energia di Gibbs Dislocazione, q Si assume che i reagenti possano trasformarsi nei prodotti solo quando le rispettive curve di energia potenziale si intersecano. In questo caso i reagenti devono “deformarsi” per raggiungere la conformazione reattiva, indicata con q*, spendendo l’energia ‡ G

34
a)In q 0 R l’elettrone si trova in un livello energetico occupato di DA (HOMO) mentre il livello energetico vacante più basso di D + A - (LUMO) ha energia troppo alta b)A seguito di riarrangiamento dei nuclei nella config. q*, l’HOMO di DA ed il LUMO di D + A - diventano degeneri e si ha il trasferimento di e - mediante attraversamento in tunnel della barriera di E potenziale. V dipende dall’energia di ionizzazione dei complessi DA e D + A -, r è la distanza da margine a margine fra D ed A. c)Il sistema si rilassa nella configurazione nucleare di equilibrio di D + A - (q 0 p ) nella quale il LUMO di DA ha energia superiore dell’HOMO di D + A -
Il sistema si rilassa nella configurazione nucleare di equilibrio di D + A - (q 0 p ) nella quale il LUMO di DA ha energia superiore dell’HOMO di D + A -.")
35
HOMO = highest occupied molecular orbital LUMO = lowest unoccupied molecular orbital L’elettrone passa quindi dall’HOMO di D (con funzione d’onda D ) al LUMO di A (con funzione d’onda A ). Se l’interazione fra queste funzioni d’onda è debole (per r > 1 nm) possiamo scrivere : v et 2 e r Dove: 2 è un operatore quanto-meccanico che descrive la sovrapposizione delle funzioni d’onda A e B r è la distanza da margine a margine fra D ed A è un parametro che misura la sensibilità nei confronti della distanza
 possiamo scrivere : v et 2 e r Dove: 2 è un operatore quanto-meccanico che descrive la sovrapposizione delle funzioni d’onda A e B r è la distanza da margine a margine fra D ed A è un parametro che misura la sensibilità nei confronti della distanza.")
36
L’energia di Gibbs di attivazione è: In cui G° è la variazione di energia libera relativa al processo di trasferimento elettronico DA D + A - e è l’energia di riorganizzazione, definita come l’energia di Gibbs necessaria per deformare la configurazione di equilibrio di DA trasformandola in quella di equilibrio di D + A -.

37
L’espressione completa di k et risulta: Limitazioni: 1)vale solo nel caso di debole accoppiamento tra le funzioni d’onda A e B, (r > 1 nm) come nel caso di quasi tutte le reazioni che prevedono trasferimenti elettronici tra proteine nel corso del metabolismo 2)Vale solo a temperature elevate per le quali le fluttuazioni termiche sono sufficienti a portare i reagenti fino allo stato di transizione, base della teoria del complesso attivato.
vale solo nel caso di debole accoppiamento tra le funzioni d’onda A e B, (r > 1 nm) come nel caso di quasi tutte le reazioni che prevedono trasferimenti elettronici tra proteine nel corso del metabolismo 2)Vale solo a temperature elevate per le quali le fluttuazioni termiche sono sufficienti a portare i reagenti fino allo stato di transizione, base della teoria del complesso attivato.")
38
Se è possibile fissare la distanza r fra donatore ed accettore, come nel caso di cofattori legati a proteine si può studiare la dipendenza di k et da r. Poiché non varia la natura di D ed A, il valore di ‡ G rimane costante, per cui prendendo il logaritmo della precedente equazione abbiamo: lnk et = -r + costante Il valore di che è possibile ricavare sperimentalmente è compreso tra 28 e 35 nm -1 nel vuoto, mentre se fra D ed A vi è un giunto molecolare 9. Se il mezzo fra A e D è una proteina, sembra essere influenzato dalla struttura secondaria, essendo: 12.5 nm -1 < <16 nm -1 per le a-eliche 9 nm -1 < <11 nm -1 per i foglietti b Altri studi indicherebbero che l’elettrone imbocchi vie specifiche lungo la proteina allo scopo di massimizzare k et. Dipendenza di k et dalla distanza

39
Dipendenza di k et da G° Per sistemi in cui siano costanti r, e , possiamo scrivere: Si prevede che il grafico di lnk et vs G° abbia forma di parabola concava verso il basso. La costante cinetica aumenta al diminuire di G°, ma solo fino a quando -G° =, condizione per la quale k et è massima. Oltre questo limite la reazione accede alla cosiddetta regione invertita, dove la costante cinetica diminuisce man mano che la reazione diventa più esoergonica. Tale comportamento si può spiegare considerando come varia ‡ G al variare di G°.

42
Teoria di Marcus: caso del RC GF°GF° ‡ G =0 GB°GB° ‡G‡G Reazione in avanti Ricombinazione di carica ‡ G = 0 Più veloce ‡ G 0 Più lenta, nonostante G B °> G F ° F B

Presentazioni simili










![N2O4 2 NO2 d[N2O4] d[NO2] V = - = 1/ 2 dt dt V = k [N2O4]n k, costante specifica di velocità, aumenta all’aumentare della temperatura. n, ordine.](/1/553488/big_thumb.jpg "N2O4 2 NO2 d[N2O4] d[NO2] V = - = 1/ 2 dt dt V = k [N2O4]n k, costante specifica di velocità, aumenta all’aumentare della temperatura. n, ordine.>")








