Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
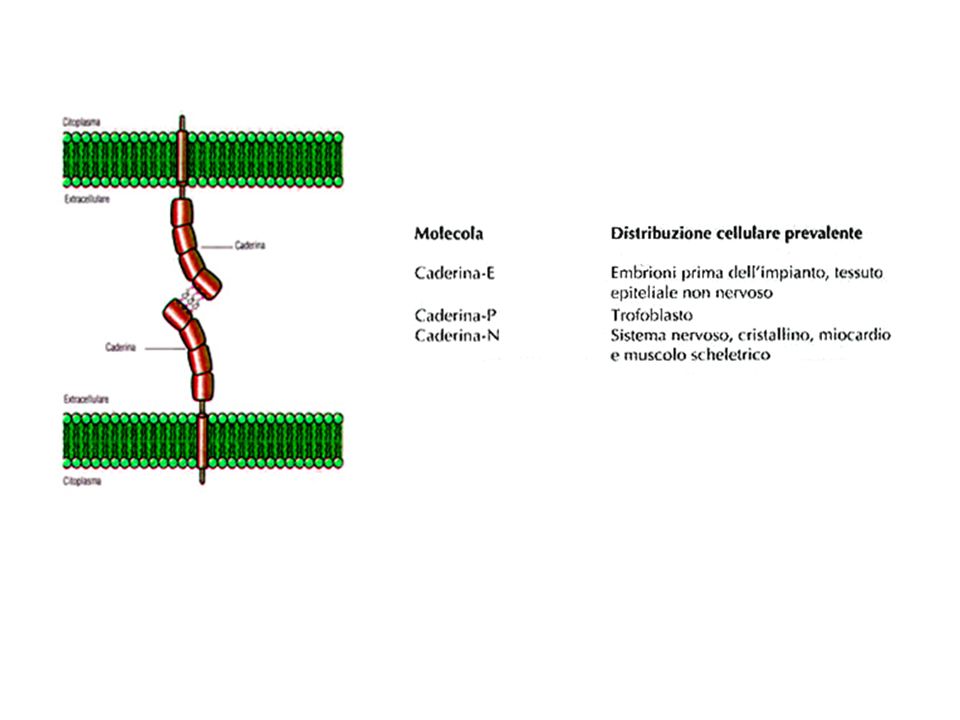
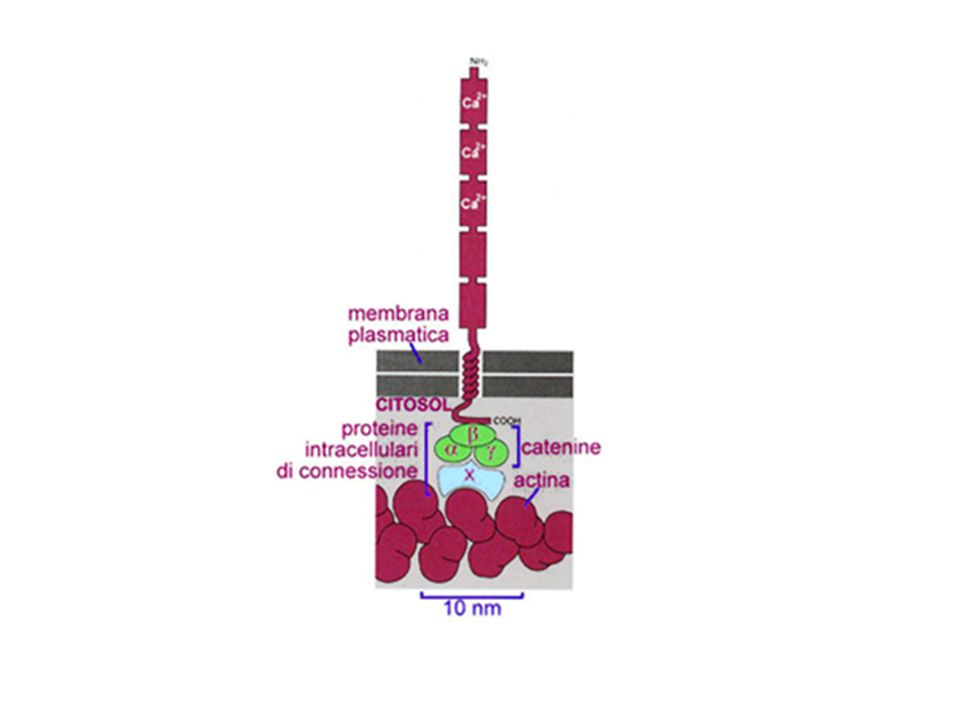
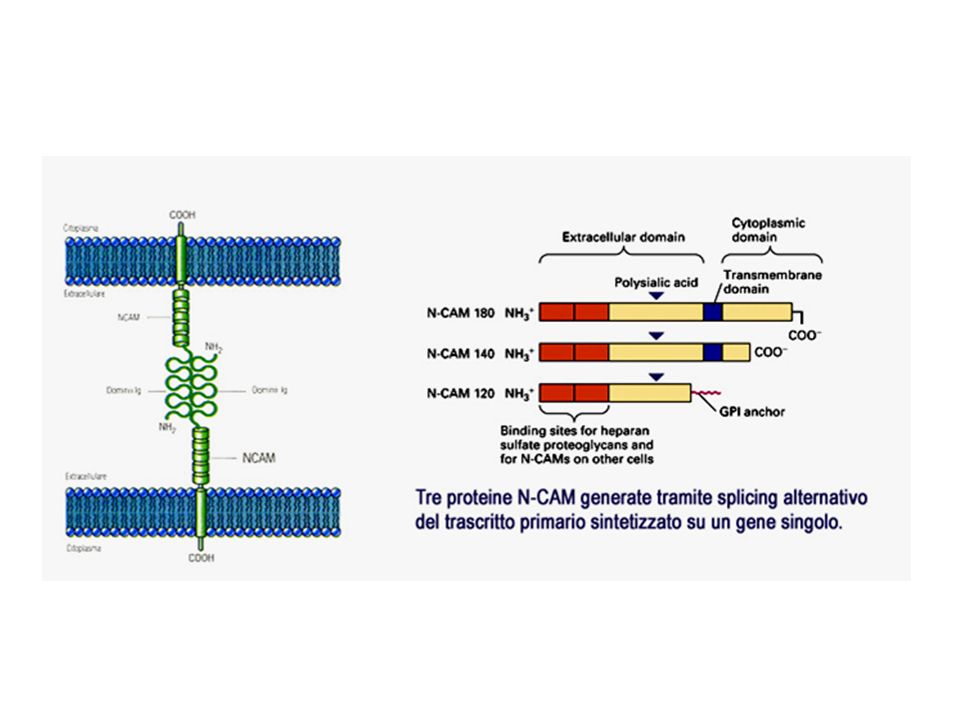
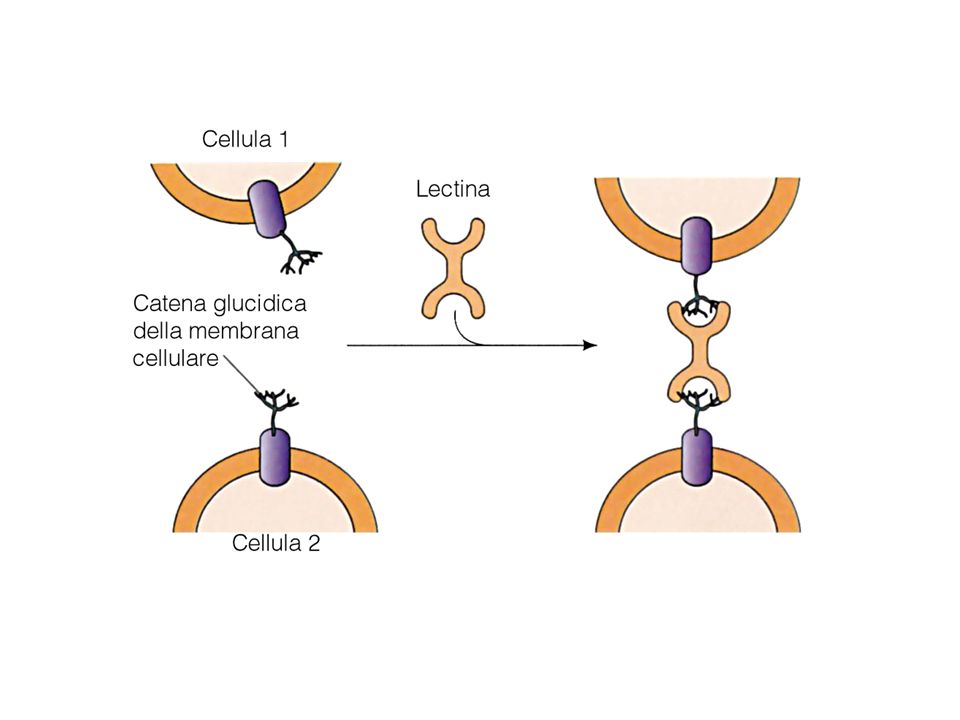
adesione

3
La matrice extracellulare La MEC delle cellule animali assume varie forme nei differenti tessuti: nell’osso la MEC è rigida e fortemente mineralizzata Nella cartilagine è meno mineralizzata e più flessibile Nel tessuto connettivo la MEC non è mineralizzata

4
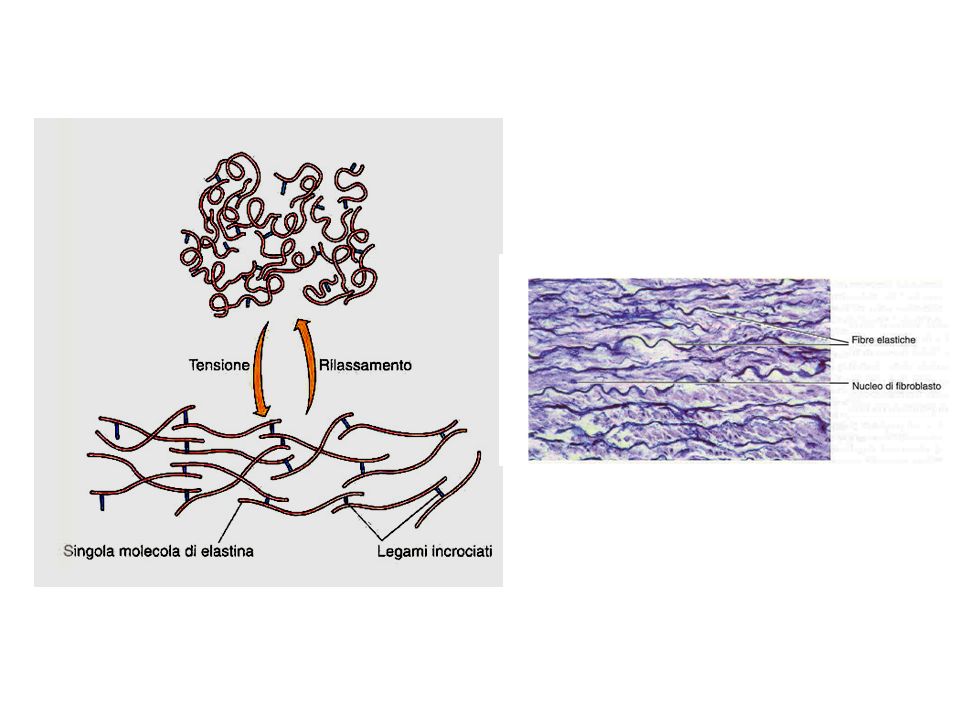
La MEC contribuisce a determinare la forma e le proprietà meccaniche dei tessuti e degli organi. Oltre a fornire un supporto strutturale influenza l’estensibilità del tessuto, la forma delle cellule ed il loro movimento. Nonostante le diverse funzioni la MEC è formata quasi sempre dalle stesse molecole: Proteine strutturali come il collagene e l’elastina che conferiscono restistenza e flessibilità Complessi di proteine e polisaccaridi, i proteoglicani, che costituiscono una matrice altamente idratabile Glicoproteine adesive, la fibronectina e la laminina, che collegano le cellule alla matrice

12
La fibronectina Le fibronectine sono glicoproteine adesive; esistono in forma solubile nel sangue, sotto forma di fibrille insolubili nella MEC ed in una forma intermedia associata alla superficie cellulare. Le diverse forme di fibronectina vengono sintetatizzate a partire da un unico RNA che viene poi processato in modi diversi per dare origine alle diverse fibronectine. La molecola di fibronectina è costituita da due catene polipeptidiche legate da due ponti disolfuro.

13
Ogni catena è organizzata in domini. Alcuni riconoscono e legano una o più molecole della MEC, altri riconoscono e legano recettori sulla membrana cellulare. La capacità della fibronectina di legare i recettori dipende da una sequanza specifica di aminoacidi, RGA (arginina-glicina- aspartato). Questa sequenza è comune a tutte le proteine adesive della MEC ed è riconosciuta dalle integrine
. Questa sequenza è comune a tutte le proteine adesive della MEC ed è riconosciuta dalle integrine.")
14
La fibronectina costituisce una molecola ponte tra le proteine della MEC e la superficie delle cellule. Essa è coinvolta sia nel mantenimento della forma cellulare che nel movimento cellulare

15
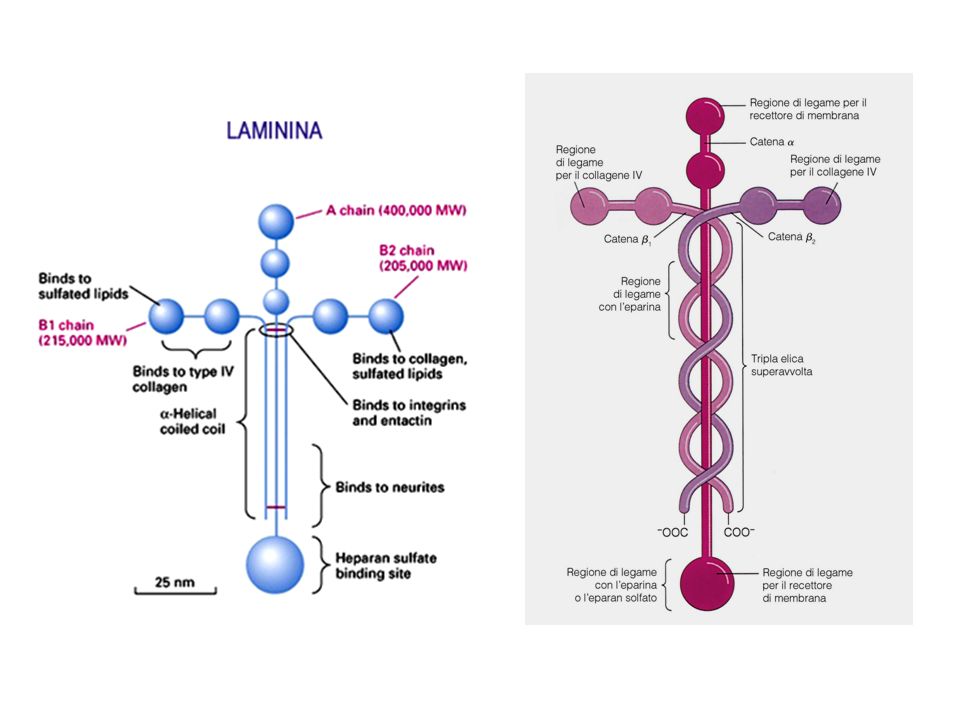
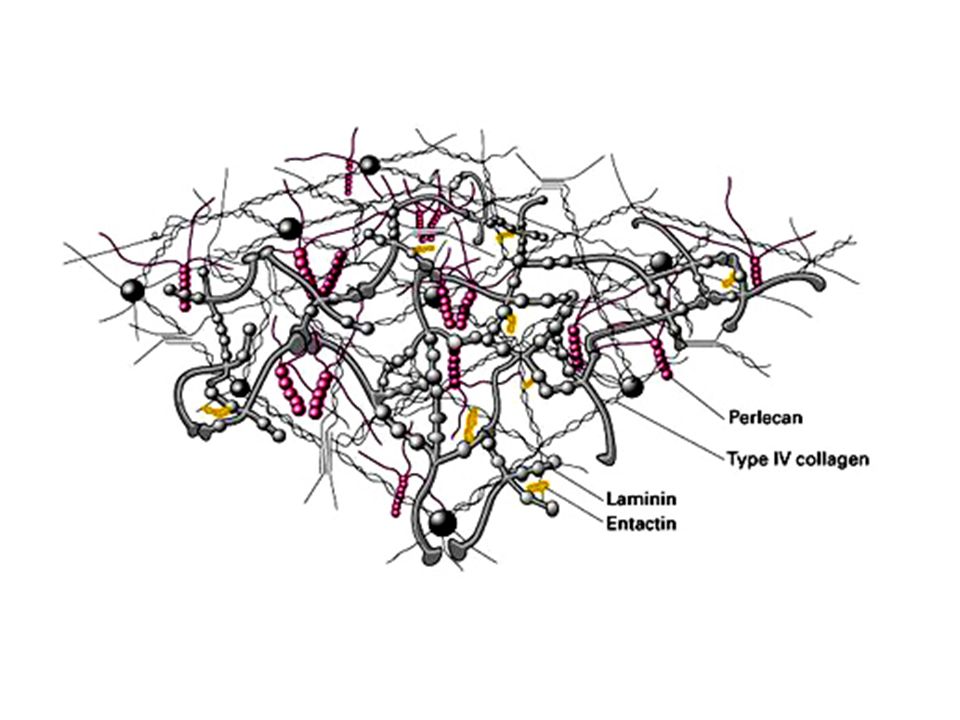
La laminina Le laminine rappresentano la seconda famiglia di glicoproteine della MEC. Sono presenti nella lamina basale, un sottile strato di materiale specializzato presente tra le cellule epiteliali e il tessuto connettivo sottostante. La membrana basale costituisce una struttura di supporto che mantiene l’organizzazione dei tessuti e costituisce una barriera di permeabilità. Tutte le lamine basali contengono, oltre alla laminina, collagene di tipo IV ed un’altra glicoproteina la entactina.

16
La laminina è una glicoproteina costituita da tre catene polipeptidiche, α, β e . Le tre catene formano una struttura a croce stabilizzata da ponti disolfuro. Come la fibronectina, anch’essa presenta domini capaci di legare il collagen di tipo IV, l’eparina, l’entactina, i proteoglicani e le superfici cellulari. Tramite questi legame essa funge da ponte tra le cellule e le lamine basali.

19
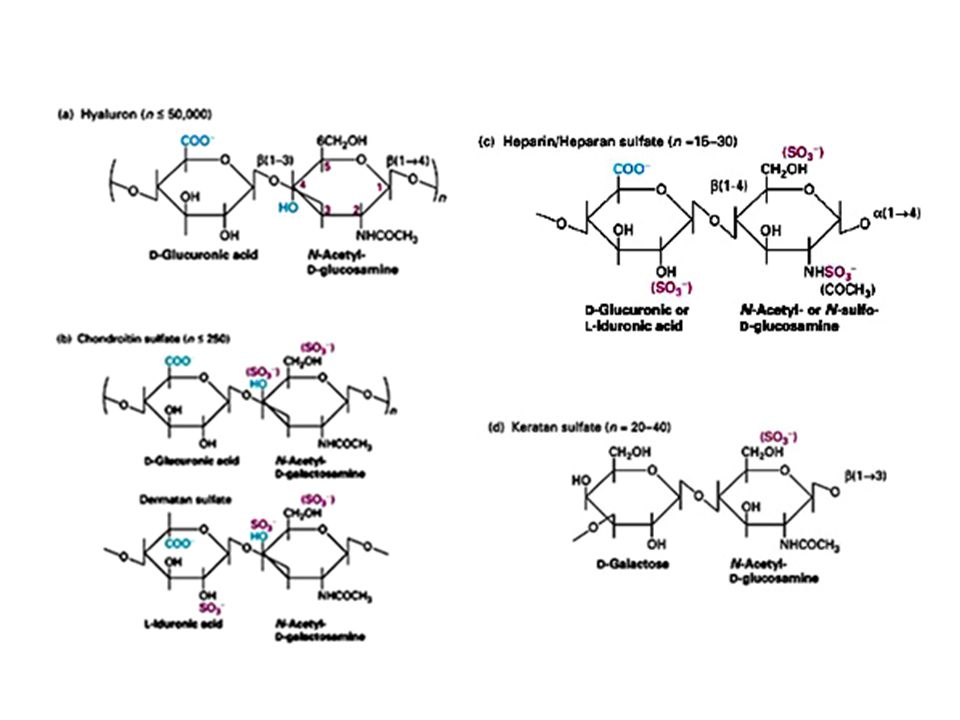
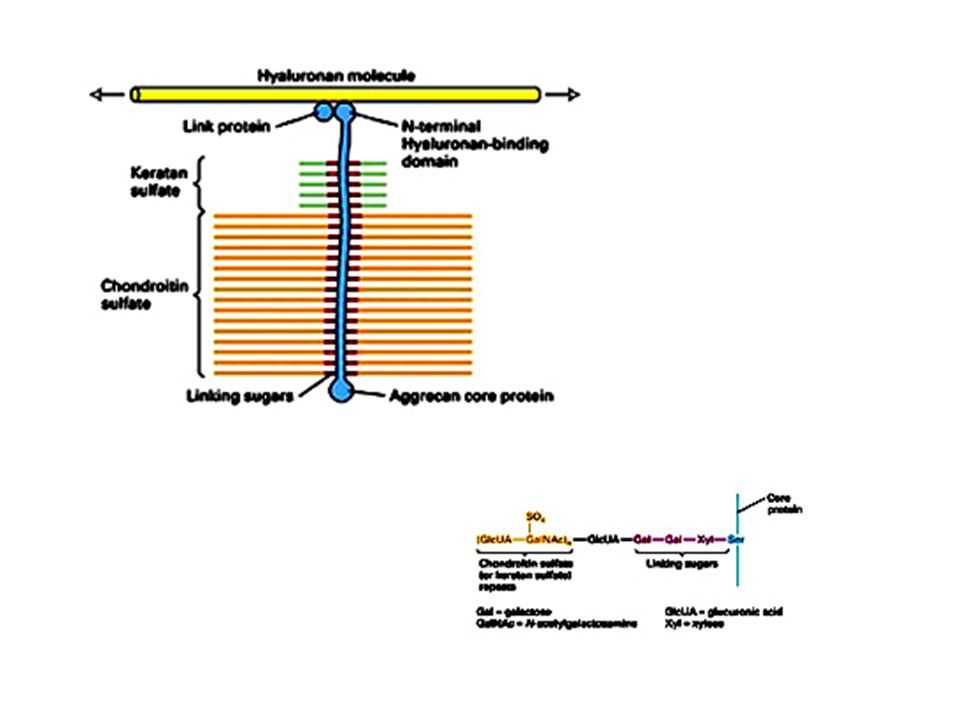
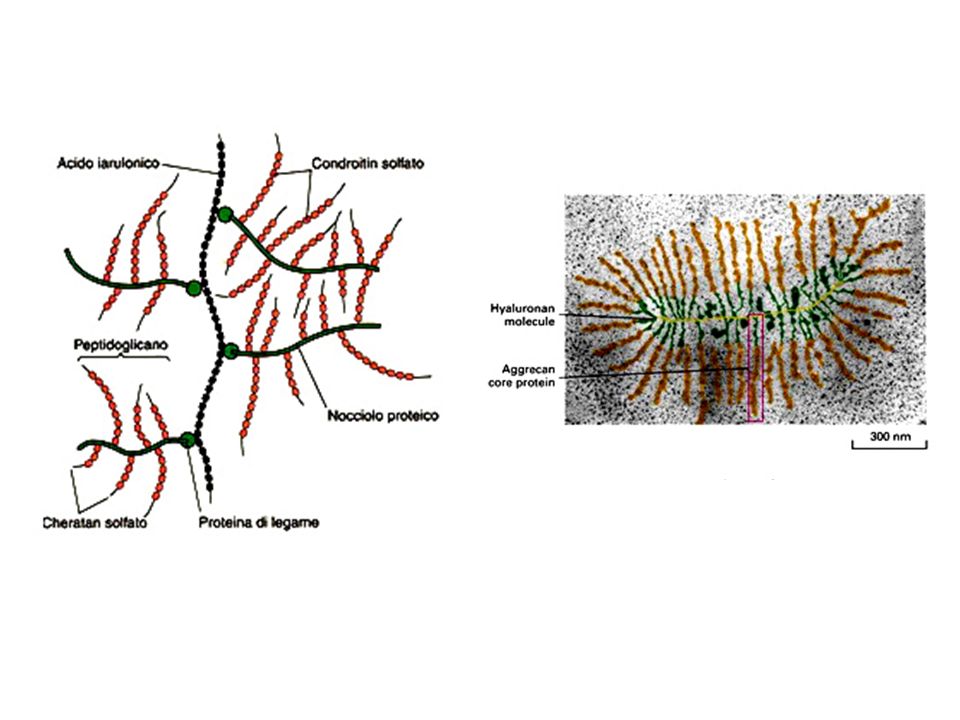
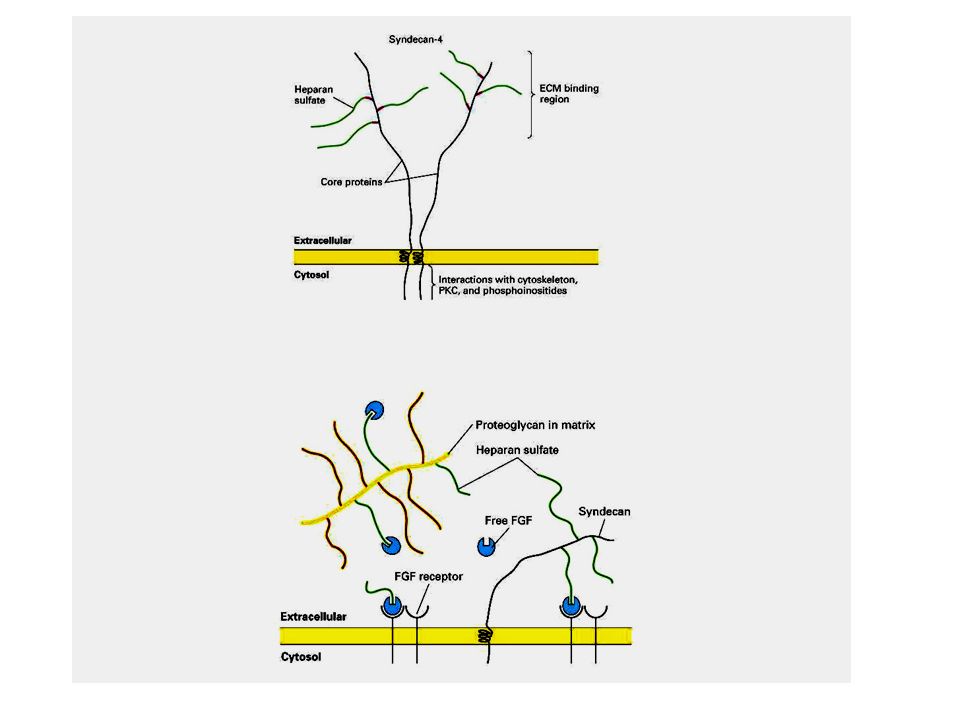
I proteoglicani Costituiscono una rete gelatinosa ed idratata in cui sono immerse le proteine della MEC. Sono glicoproteine che portano legati un certo numero di glicosaminoglicani (GAG). I GAG sono costituiti da unità disaccaridiche ripetute tra cui i più comuni sono l’acido ialuronico, il condroitin solfato, il cheratan solfato. In queste molecole uno degli zuccheri che compongono il disaccaride è un aminozucchero, l’altro è generalmente uno zucchero acido come il galattosio. I GAG si legano covalentemente alle proteine formando i proteoglicani
. I GAG sono costituiti da unità disaccaridiche ripetute tra cui i più comuni sono l’acido ialuronico, il condroitin solfato, il cheratan solfato. In queste molecole uno degli zuccheri che compongono il disaccaride è un aminozucchero, l’altro è generalmente uno zucchero acido come il galattosio. I GAG si legano covalentemente alle proteine formando i proteoglicani.")
23
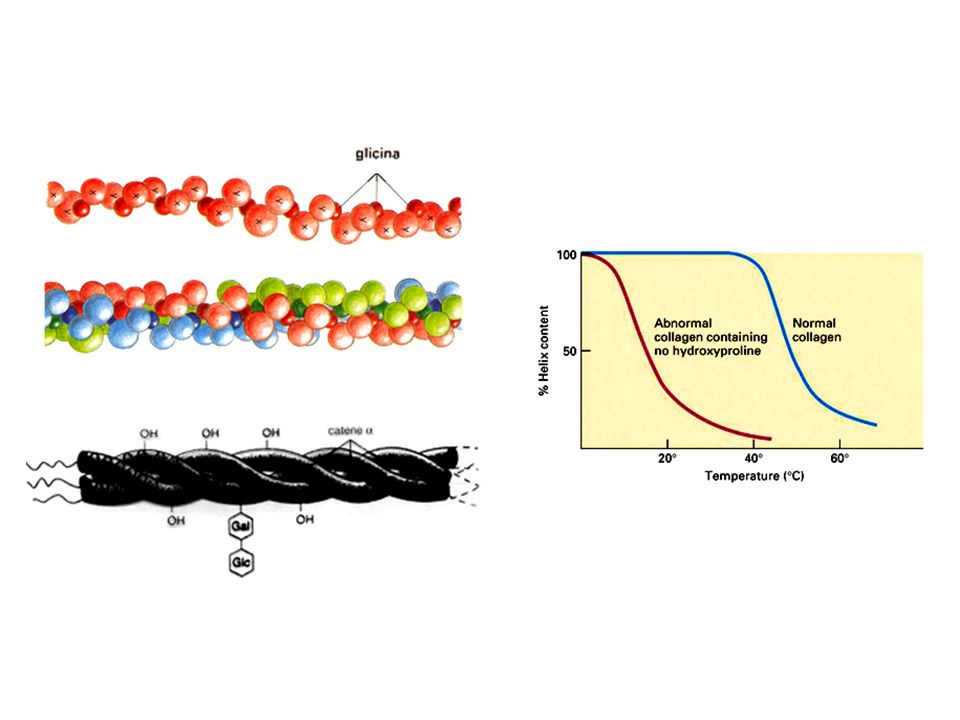
Il collagene Sono la componente più abbondante della MEC. Formano fibre ad elevata resistenza alla trazione Sono secreti da diversi tipi cellulari del tessuto connettivo Tutti i collageni hanno due caratteristiche specifiche: una tripla elica rigida e una composizione aminoacidica particolare

25
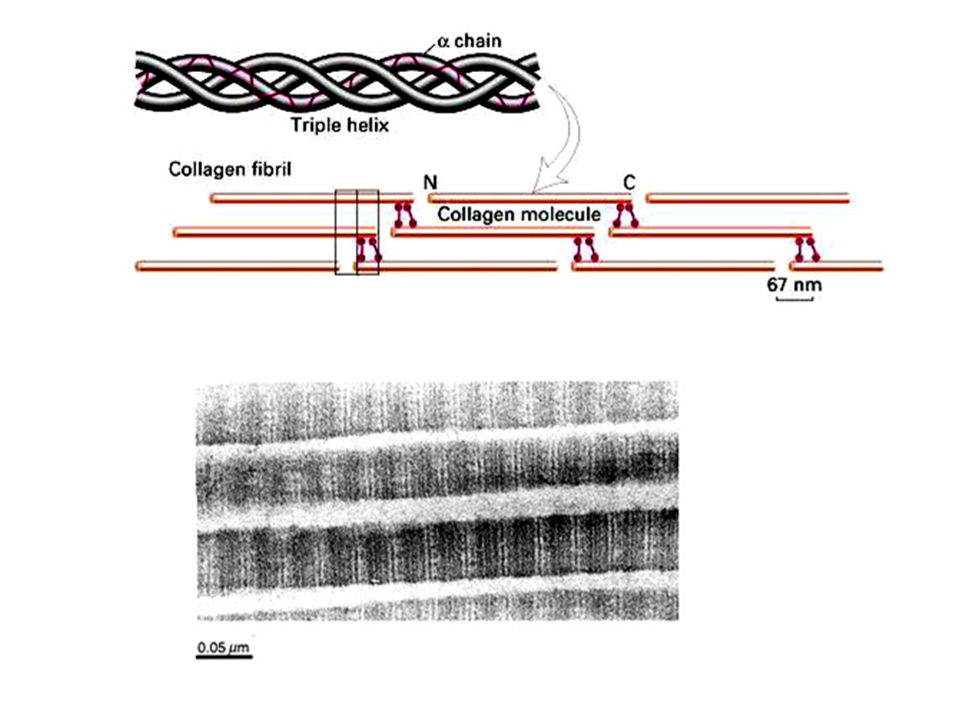
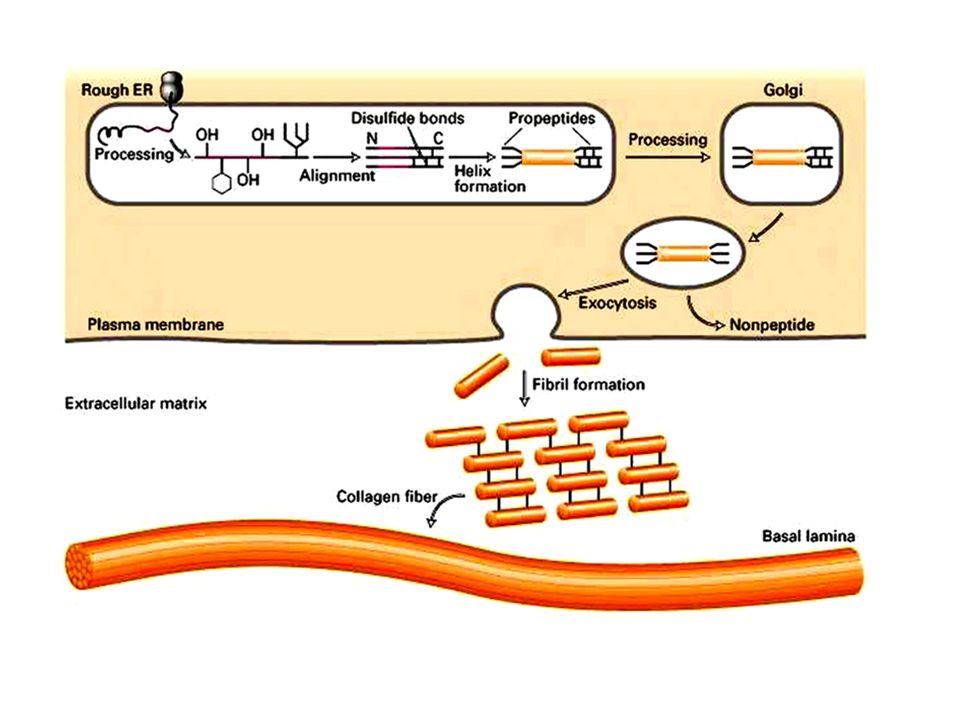
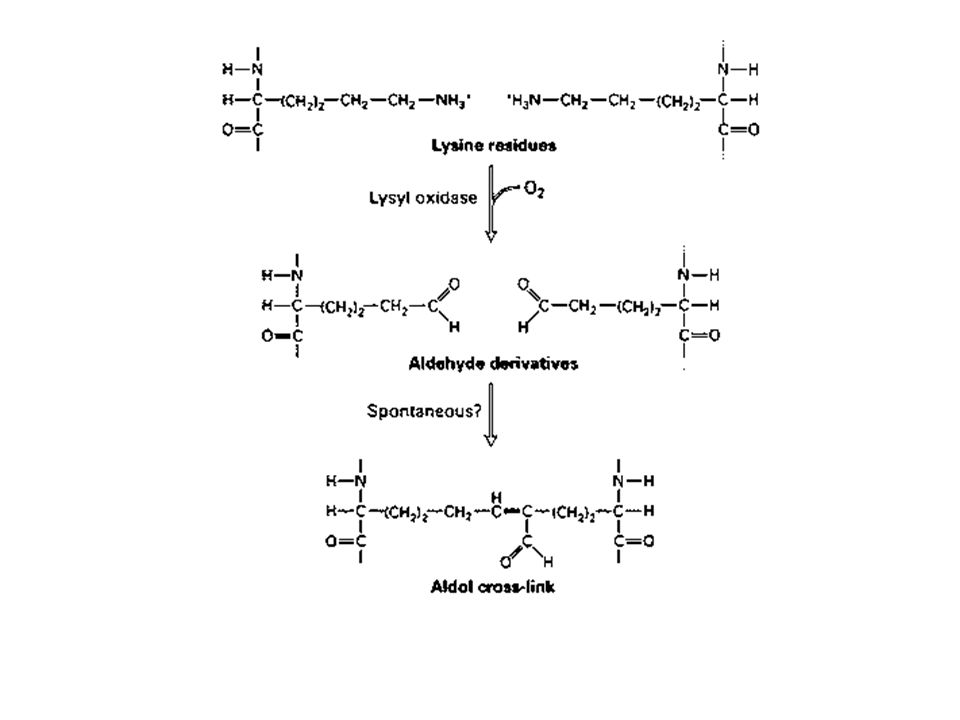
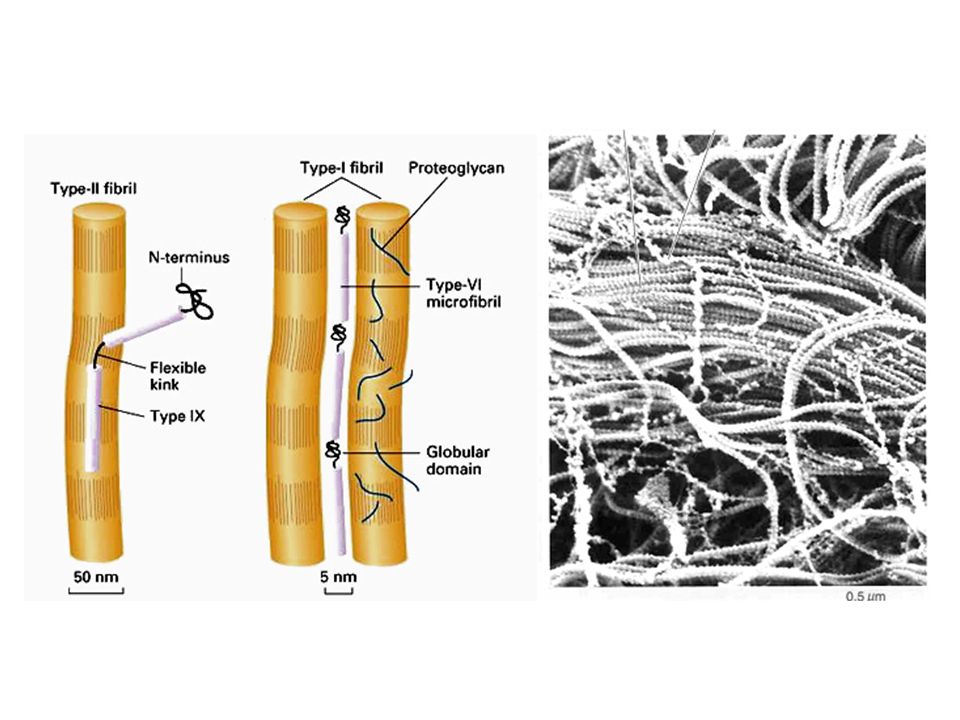
Nella maggior parte dei tessuti le fibre di collagene sono presenti in fasci sparsi nella MEC. Una caratteristica delle fibre collagene è quella di essere composte da numerose fibrille che sono a loro volta costituite da numerose molecole di collagene. Il collagene viene sintetizzato nel RE dove tre catene alfa si assemblano a formare una tripla eleica di protocollagene. Una volta secreto nello spazio extracellulare il protocollagene viene convertito in collagene da un enzima, la procollagen peptidasi, che rimouve le estremità aminoterminali e carbossiterminali. Le molecole risultanti si assemblano spontaneamente e formano le fibrille di colalgene mature che successivamente si associano in fibre collagen.

33
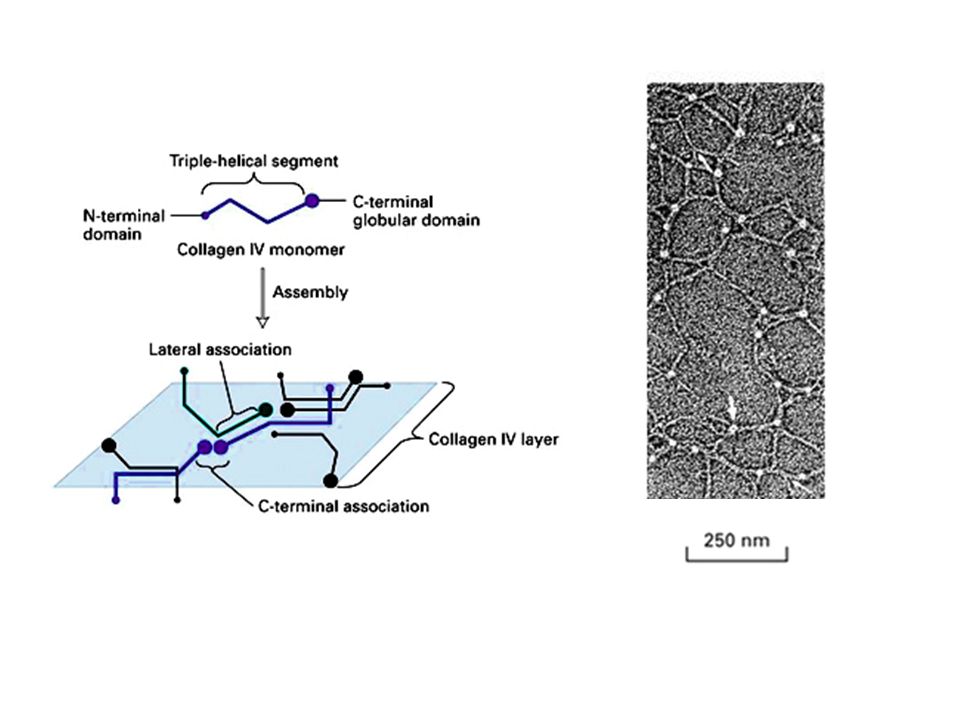
Sono noti diversi tipi di collagene: Il collagene di tipo I, II e III, che sono i più abbondanti ed il collagene di tipo IV presente particolarmente nelle lamine basali.

Presentazioni simili
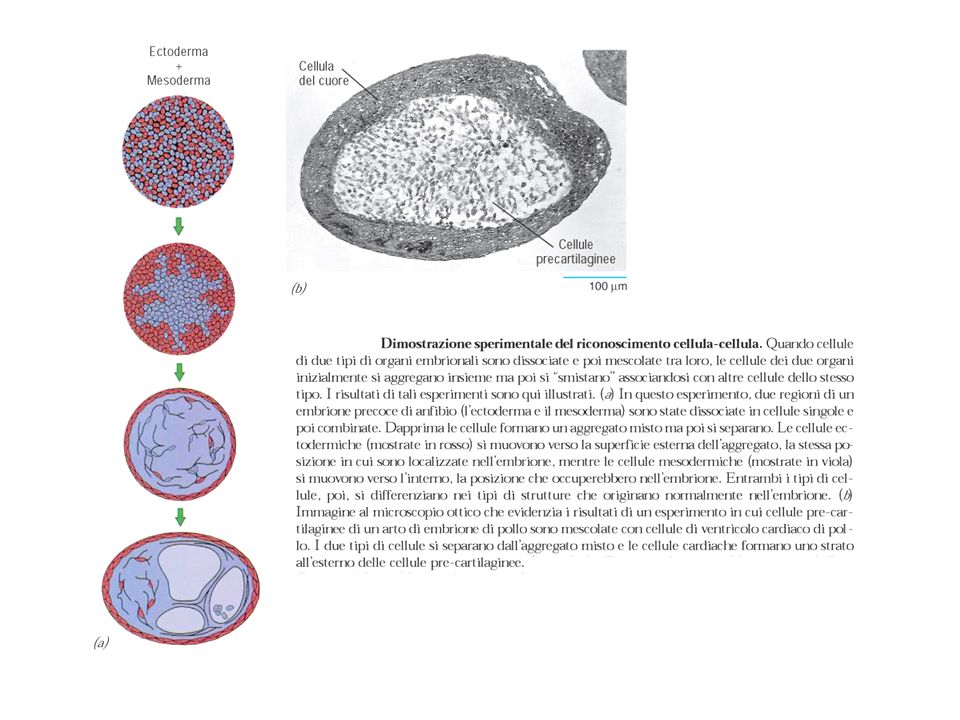
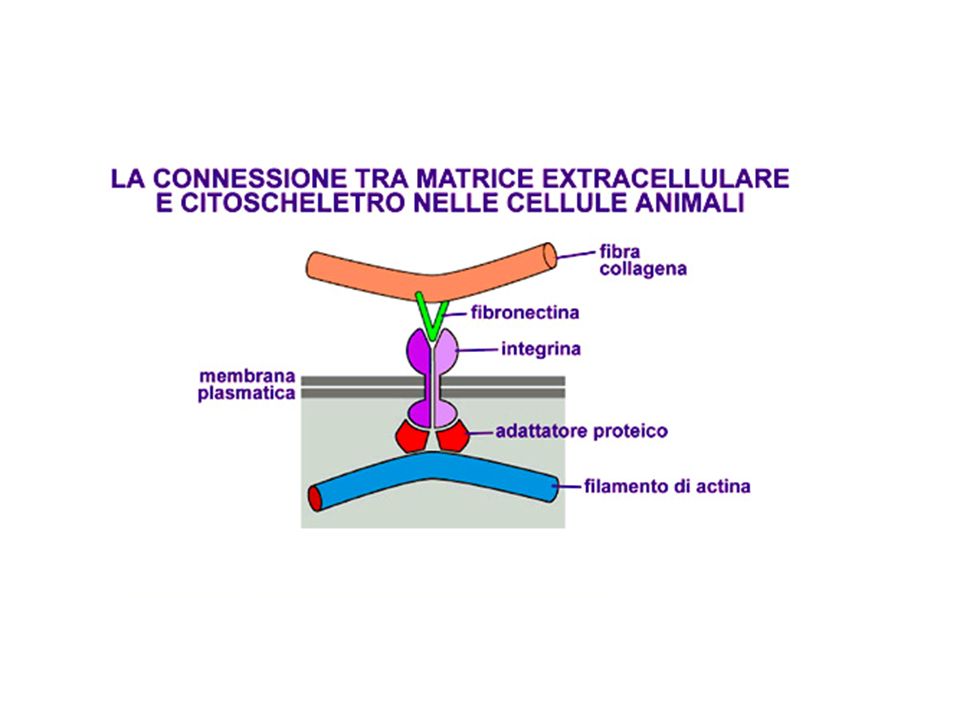
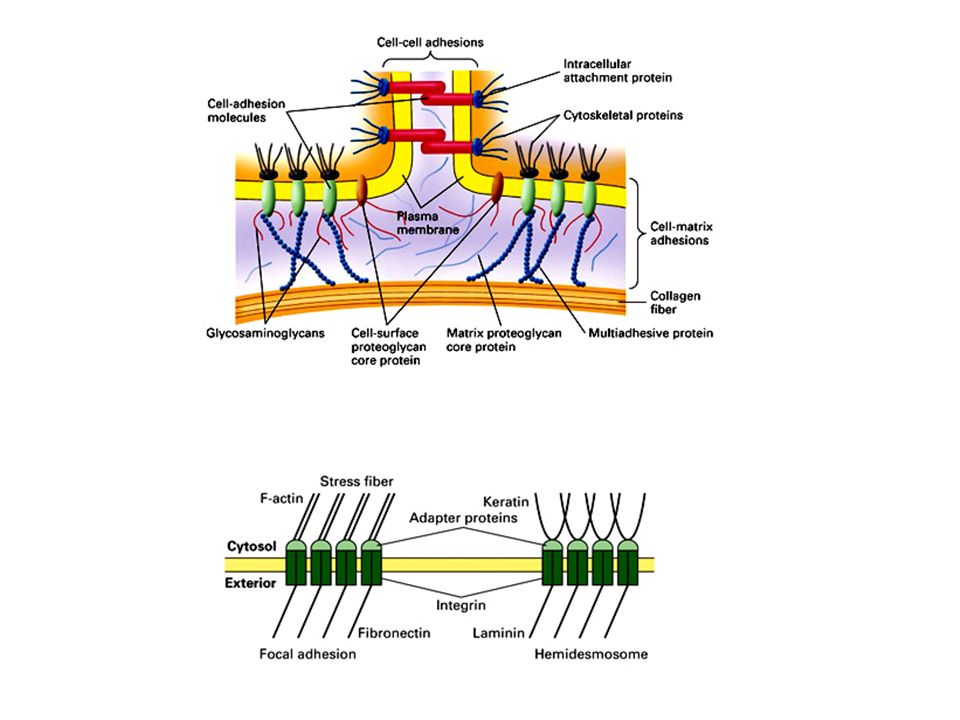
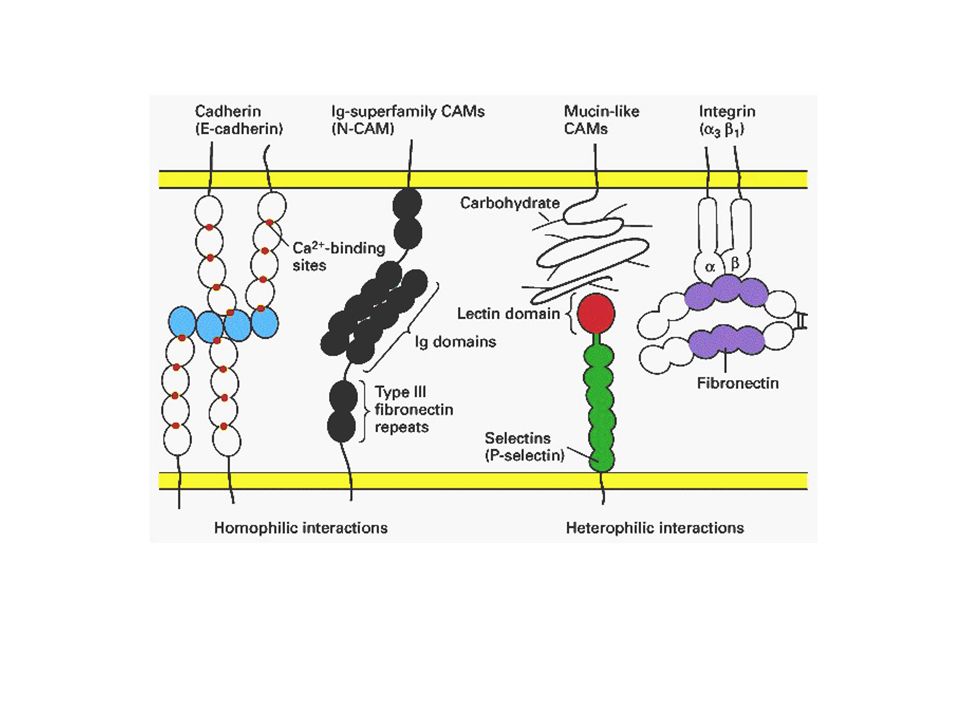


 formazione del centro di ossificazione b) accrescimento c) modellamento e mineralizzazione Es.: ossa del cranio,>")








>")







