Scaricare la presentazione
1
RISPOSTA CELLULARE ALL’AMBIENTE Molecole segnale Recettori Trasduzione del segnale

2
? Gli organismi multicellulari hanno GROSSI problemi di comunicazione
Oh ! Abbiamo bisogno di glucosio ! Ehi tu – dividiti adesso !!! Per piacere, vuoi smettere di dividerti ? Avanti N. 7, il tuo tempo è scaduto !

3
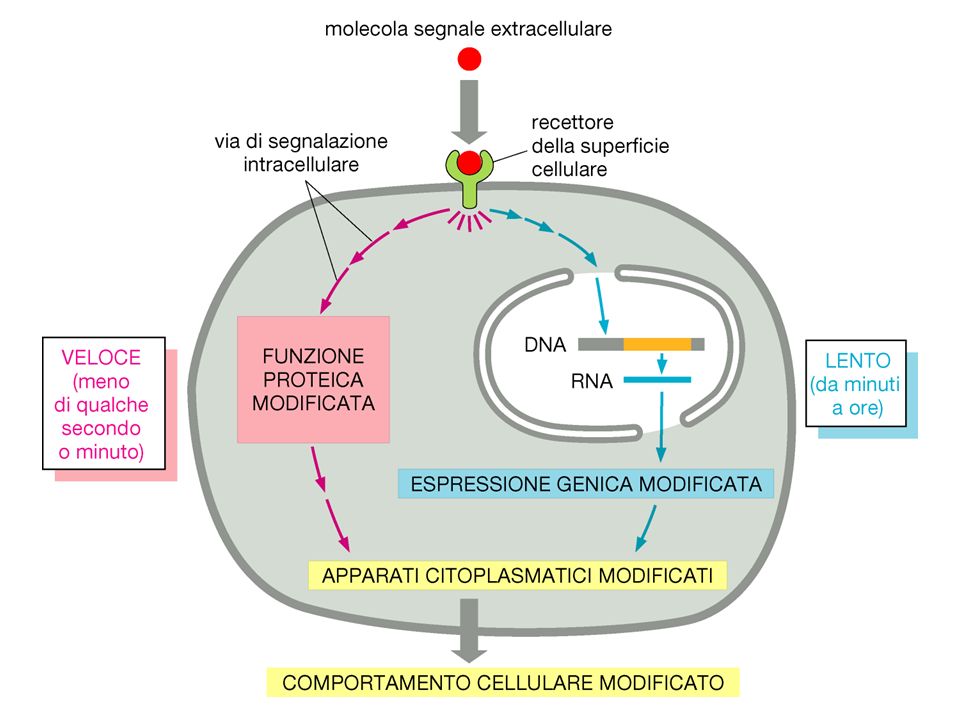
Come rispondono le cellule ai segnali ?
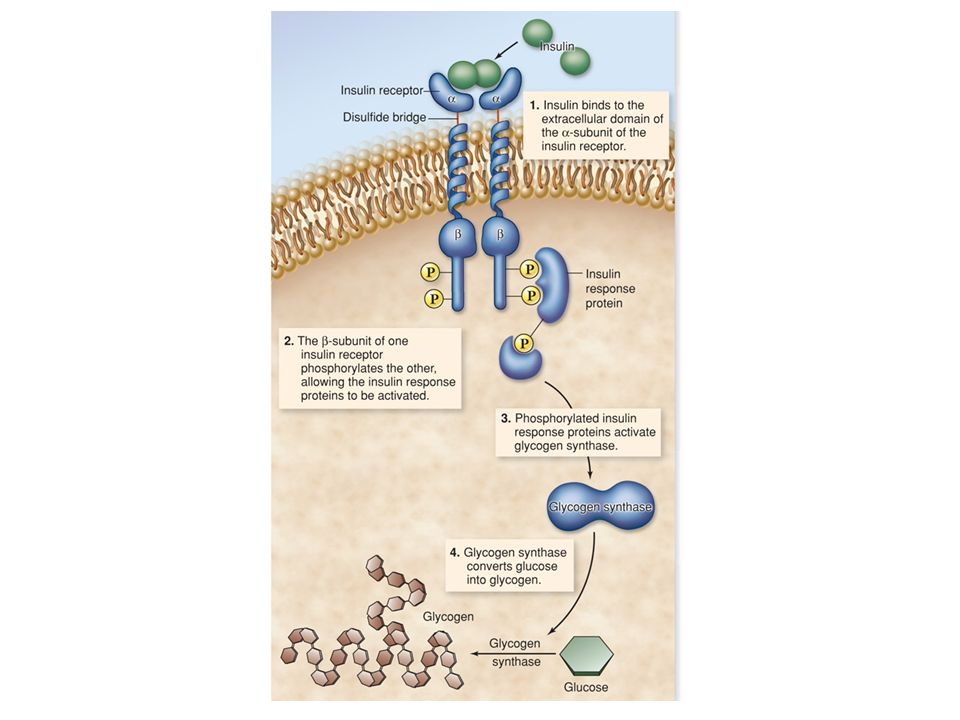
Alterazione del metabolismo alterazione metabolismo del glicogeno in risposta all’insulina Eccitamento propagazione impulso nervoso in risposta a neurotrasmettitori Crescita e Divisione risposta a fattori di crescita Morte cellulare programmata specifici fattori di “morte” o rimozione di alcuni fattori essenziali Espressione genica alterata sintesi di Ig in risposta a citochine

4
I Problemi del Signalling
Le cellule sono esposte a una moltitudine di segnali provenienti da altre cellule o da stimoli ambientali. Selezionare i segnali rilevanti da quelli irrilevanti Recettori con un alto grado di specificità Recettori ad alta affinità accoppiati ad un sistema di amplificazione Attivazione di vie di signalling basate su processi comuni Captare segnali a basse concentrazioni Tradurre segnali diversi in un comune “linguaggio”

5
Una risposta cellulare può essere determinata da:
presenza di mediatori chimici (ormoni o altre molecole) interazione con altre cellule o con strutture extracellulari
 interazione con altre cellule o con strutture extracellulari.")
7
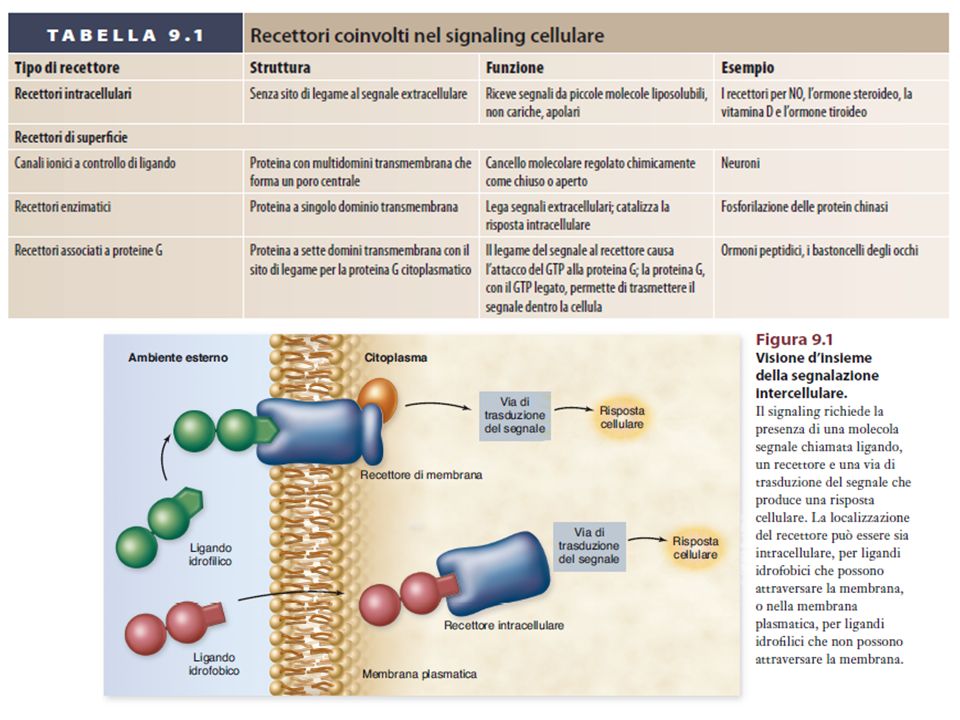
Il signaling endocrino è più lento e richiede recettori ad alta affinità (il segnale ha concentrazioni più basse nel sangue) Il controllo attraverso contatto è dovuto alla presenza di proteine sulla membrana plasmatica. Il controllo tramite mediatori chimici può avvenire con meccanismo endocrino, paracrino o sinaptico.

9
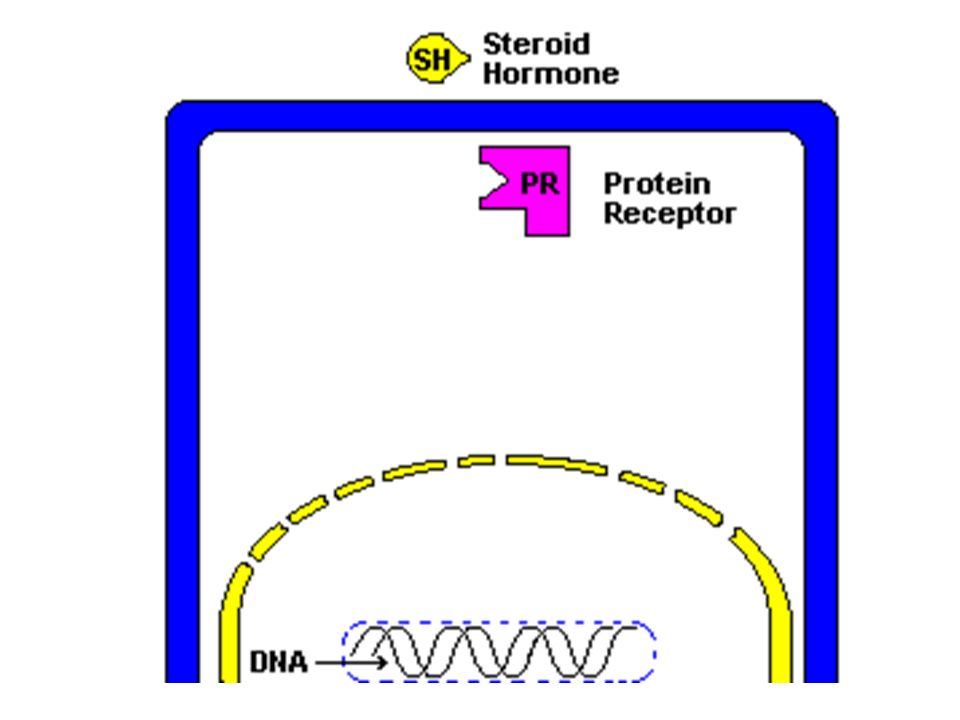
Gli ormoni steroidei e i mediatori chimici liposolubili attraversano la membrana plasmatica e si legano a recettori citoplasmatici

13
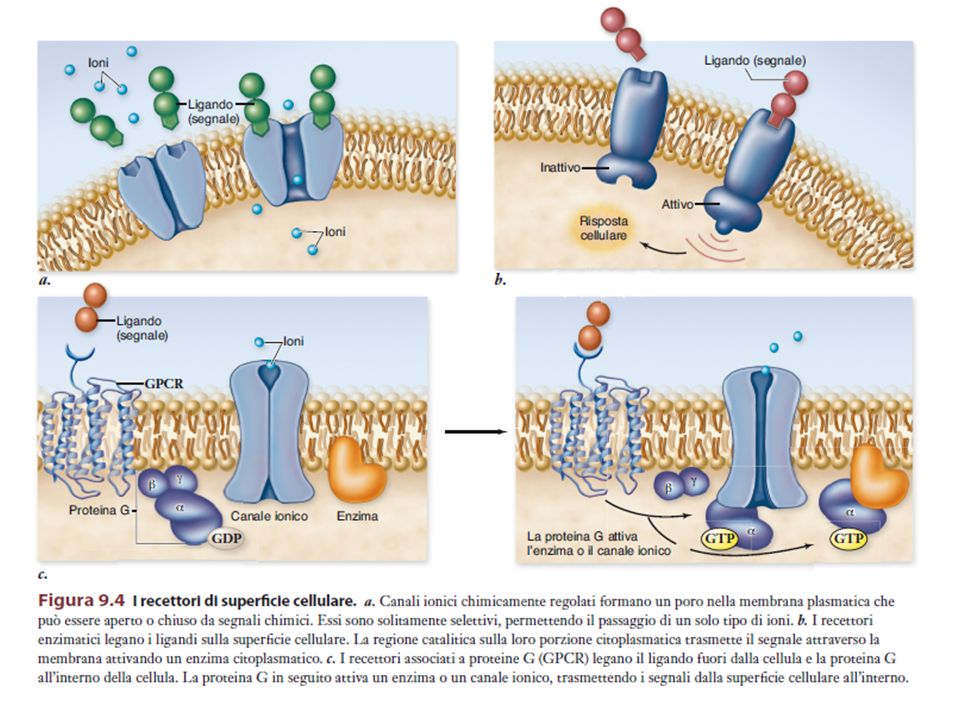
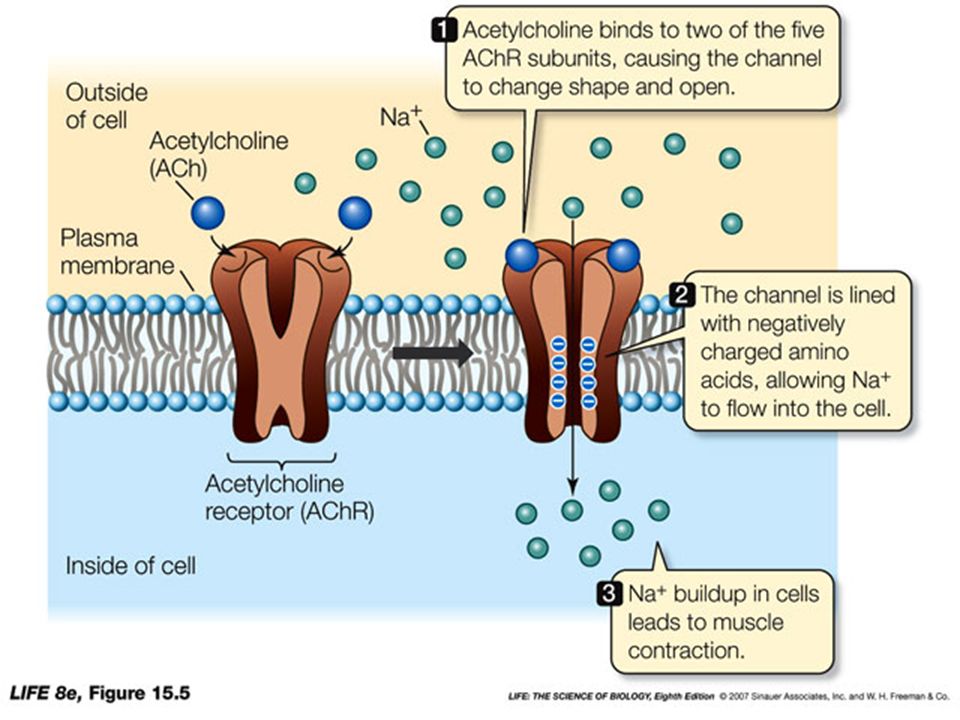
RECETTORI ENZIMATICI (es. TIROSIN-CHINASICI)
I RTK, presenti sulla membrana nella forma monomerica inattiva, dimerizzano e si attivano in seguito al legame di due ligandi. I residui di Tyr presenti nel segmento citoplasmatico dei RTK vengono fosforilati (autofosforilazione). Le Tyr fosforilate vengono riconosciute da molecole citoplasmatiche specifiche, che vengono a loro volta attivate.
. Le Tyr fosforilate vengono riconosciute da molecole citoplasmatiche specifiche, che vengono a loro volta attivate.")
15
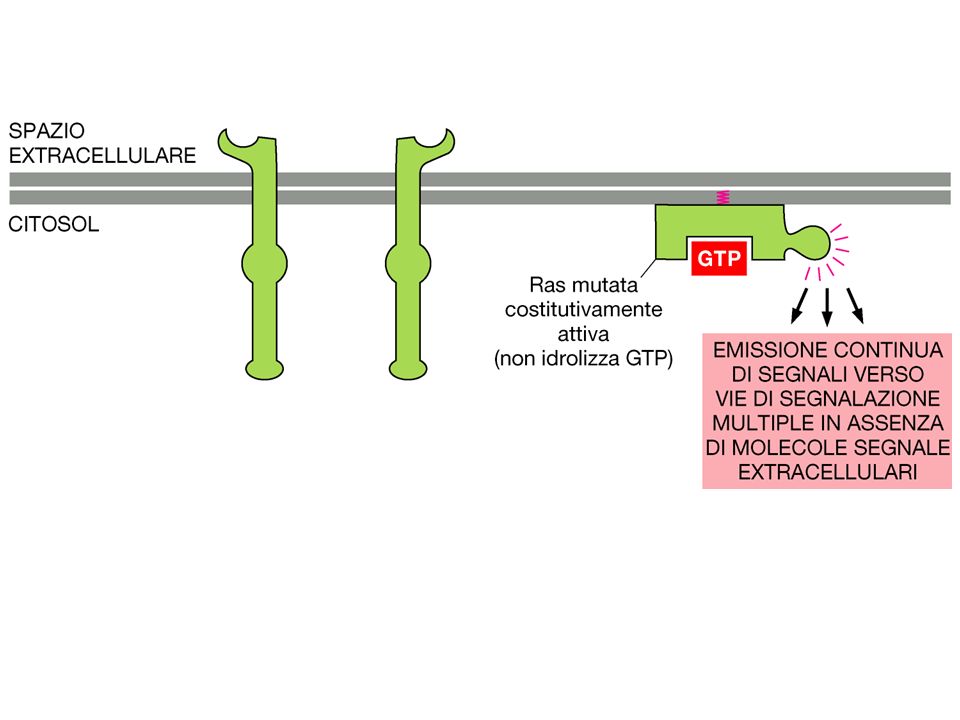
MOLTI RECETTORI PER FATTORI DI CRESCITA SONO RTK
La fosforilazione dei residui Tyr determina l’attivazione della proteina RAS, importante nel controllo della proliferazione cellulare.

16
MAP: protein chinasi attivate da mitogeni
Ad ogni passaggio l’azione enzimatica della chinasi su substrati multipli causa l’amplificazione del segnale

18
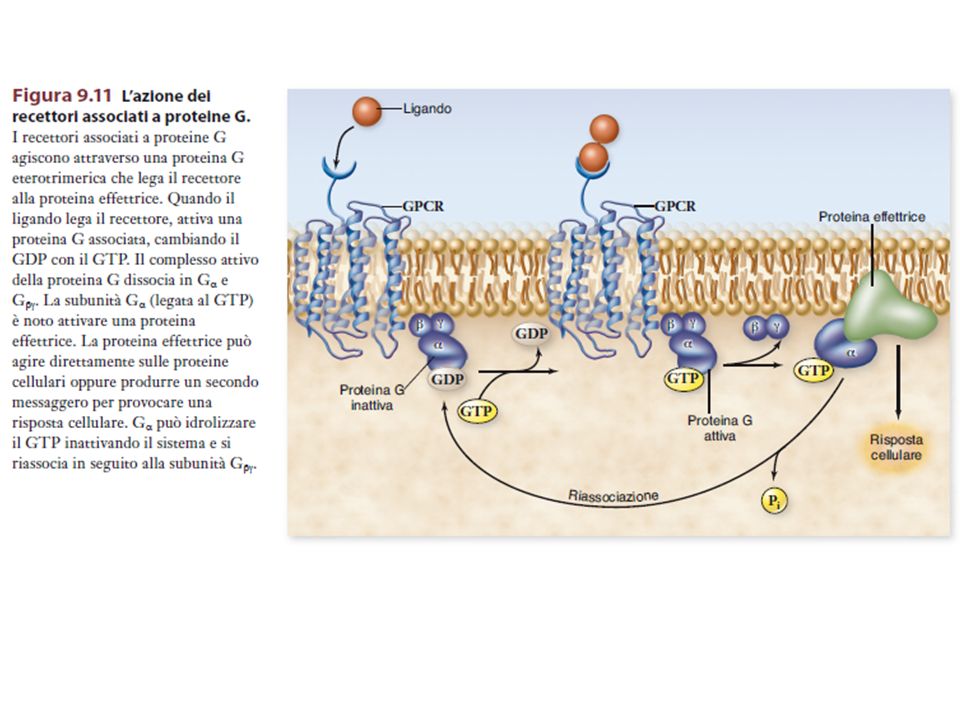
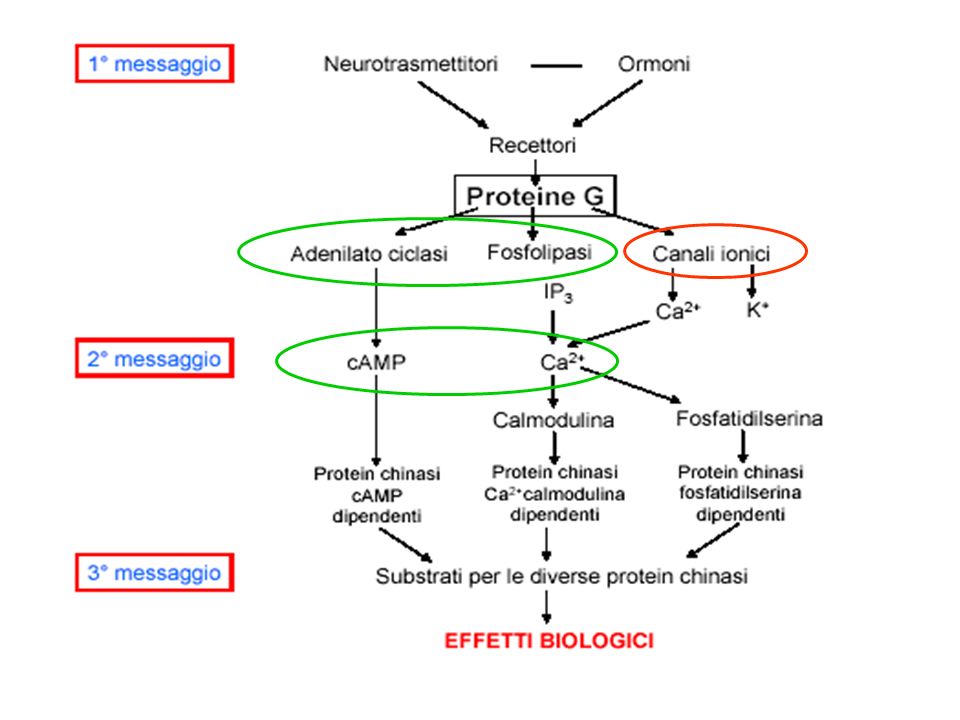
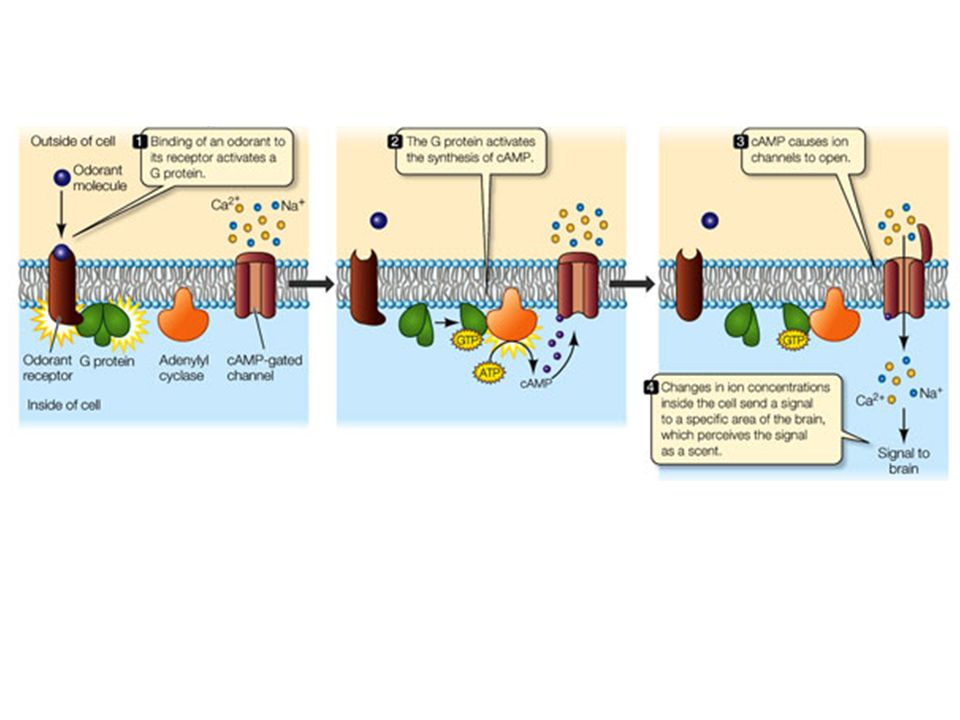
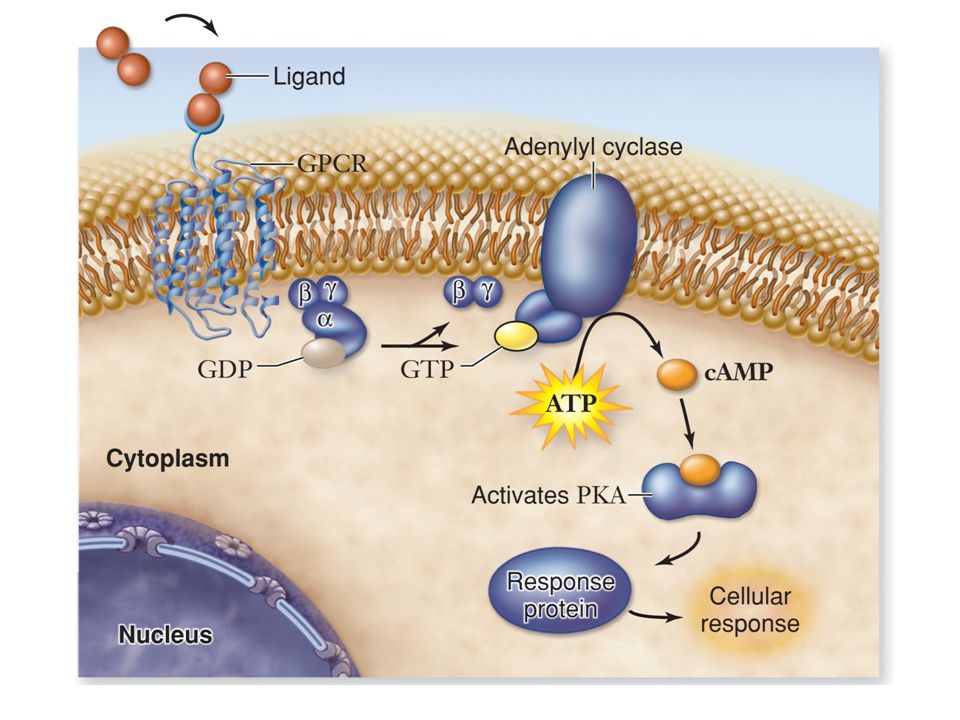
Alcuni recettori di membrana non attivano direttamente la risposta cellulare, ma un’altra proteina di membrana, la proteina G (proteina che lega il GTP), la quale attiva un effettore, direttamente legato alla formazione del secondo messaggero.
, la quale attiva un effettore, direttamente legato alla formazione del secondo messaggero.")
19
Recettori accoppiati alla proteina G
Presenti in tutti gli eucarioti > recettori Struttura simile 7 tratti transmembrana Funzioni biologiche diverse Molecole segnale: proteine, peptidi, lipidi e altre piccole molecole

20
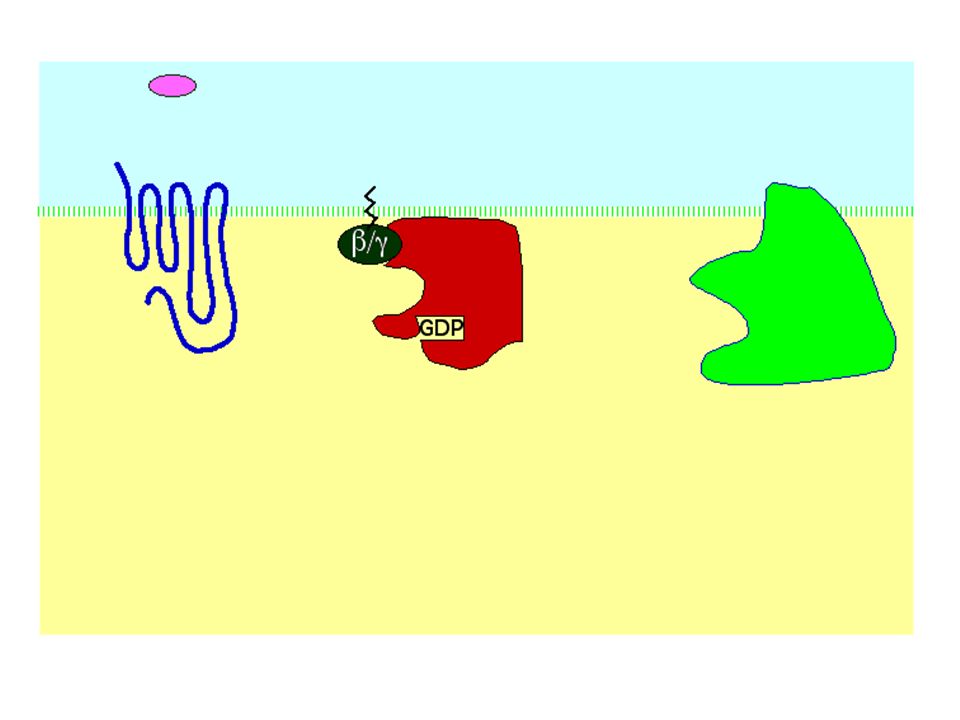
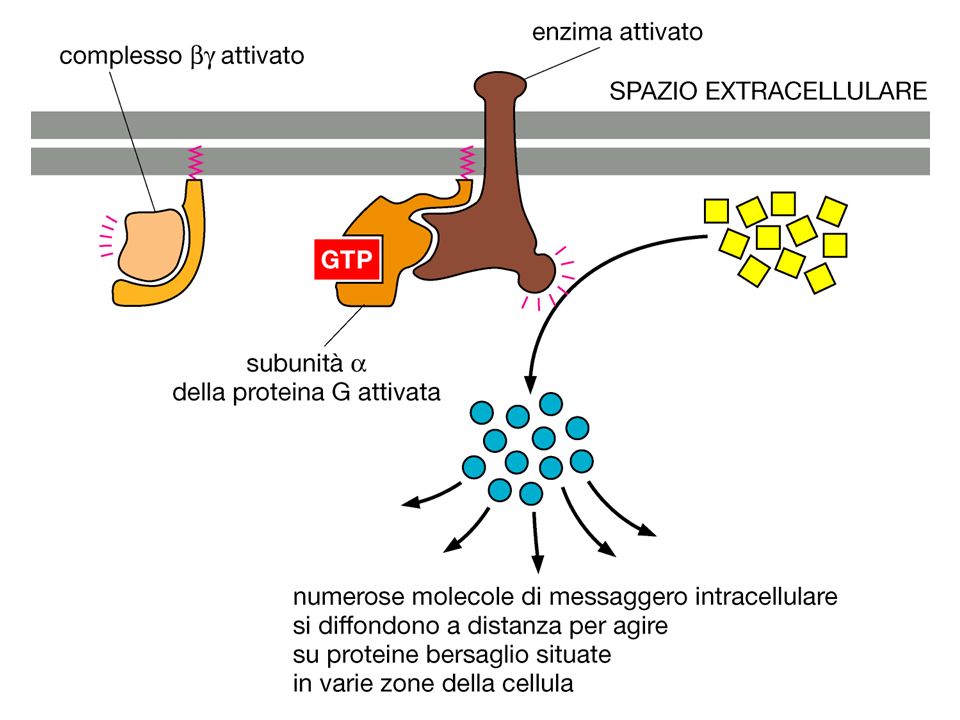
INATTIVA ATTIVA GTP GDP GDP GTP Pi GDP
Scambio del GDP legato col GTP INATTIVA GTP GDP GDP La subunità si dissocia da GTP ATTIVA Pi GDP Attività GTPasica della subunità GTP GDP+Pi La subunità attiva può interagire con lo step successivo della catena di signalling La subunità si riassocia a

22
I bersagli dei componenti dissociati delle proteina G sono enzimi o canali ionici che trasmettono in avanti il segnale

26
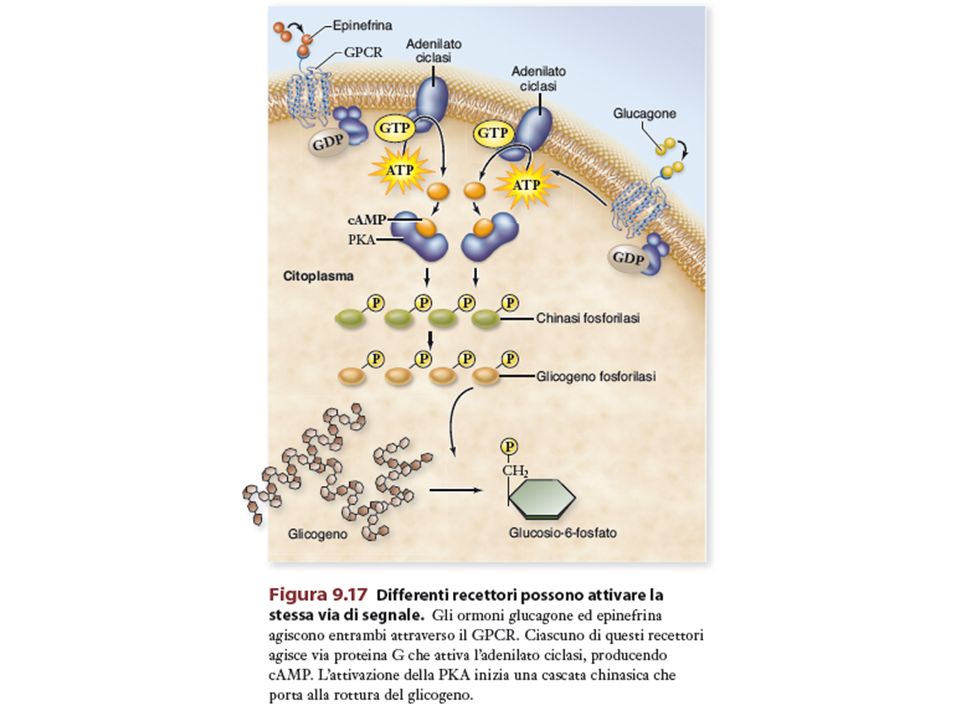
La formazione di cAMP a partire da ATP è catalizzata dall’ adenilato ciclasi, presente nella membrana plasmatica. Questo enzima è attivato dopo il legame del recettore con uno specifico mediatore. La demolizione di cAMP (altrettanto rapida quanto la sintesi) avviene ad opera di cAMP fosfodiesterasi (inibita da caffeina, teofillina).
 avviene ad opera di cAMP fosfodiesterasi (inibita da caffeina, teofillina).")
28
Il cAMP è un esempio di secondo messaggero molto utilizzato nelle cellule. Il cAMP attiva diverse chinasi in grado di iniziare processi cellulari differenti, oppure attivare lo stesso processo anche se la sua formazione deriva da vie di signalling diverse.

30
Lo stesso segnale può attivare vie multiple di segnalazione a seconda di: 1) tipo di recettore 2) tipo di proteine G a cui è accoppiato il recettore (Gs o Gi) 3) bersagli enzimatici attivati nella cellula cuore ADRENALINA intestino Attivazione proteina G Attivazione proteina G Adenilato ciclasi Adenilato ciclasi cAMP cAMP Aumento frequenza e contrazione cardiaca Rilassamento

31
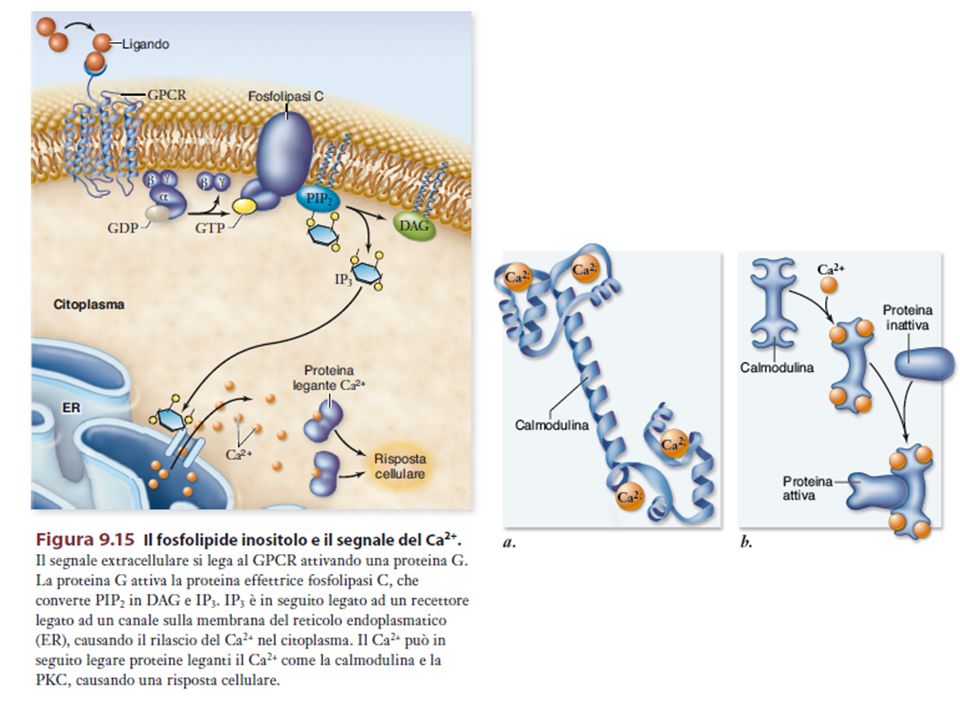
Calcio – il messaggero universale
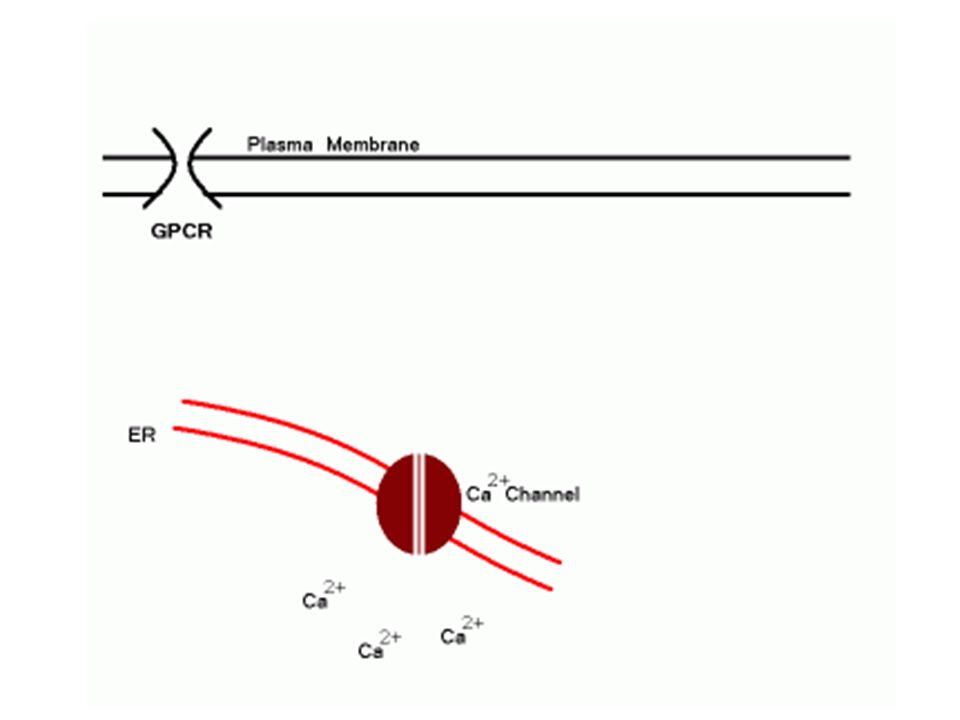
Il Ca2+ è mantenuto ad una concentrazione estremamente bassa nel citosol (a causa della sua tossicità); tuttavia la concentrazione del Ca2+ è alta fuori dalla cellula e all’interno di alcuni organelli (“compartimento che sequestra il calcio”) L’apertura rapida e transitoria di canali permette al Ca2+ di fluire nel citosol seguendo il suo gradiente di concentrazione e costituisce la base di un sistema di signalling.
; tuttavia la concentrazione del Ca2+ è alta fuori dalla cellula e all’interno di alcuni organelli ( compartimento che sequestra il calcio ) L’apertura rapida e transitoria di canali permette al Ca2+ di fluire nel citosol seguendo il suo gradiente di concentrazione e costituisce la base di un sistema di signalling.")
36
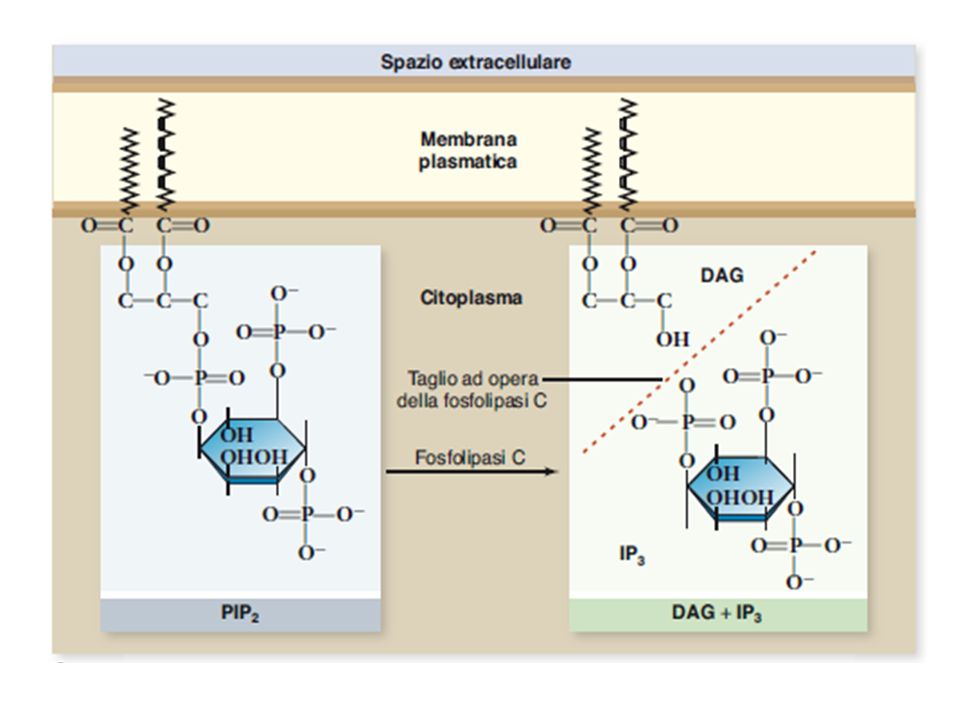
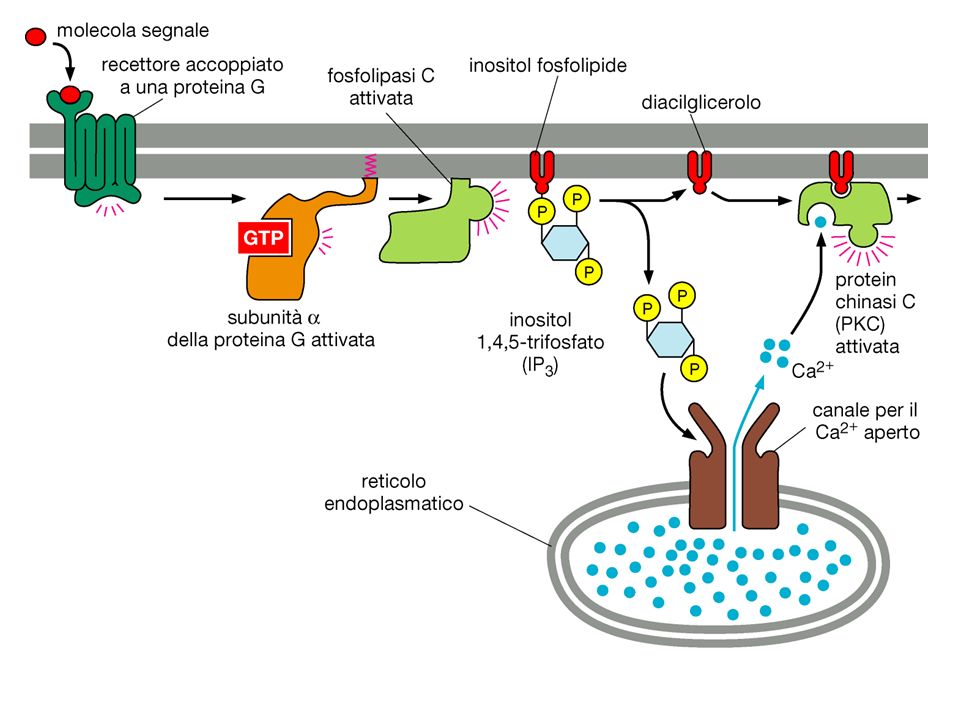
DAG PLC La Calmodulina può attivare pompe del Ca2+ sulla membrana plasmatica L’IP3 si lega al recettore per l’IP3 sul RE e apre un canale del Ca2+ Il Ca2+ rilasciato si lega alle Calmodulina che interagisce con altre proteine attivandole kinasi La Calmodulina attiva delle pompe del Ca2+ del RE La Calmodulina attiva una vasta gamma di proteine (chinasi calmodulina-dipendenti)
")
38
Amplificazione e diversi step di regolazione

39
Comunicazione incrociata (cross-talk)
Ciascun recettore attiva (frecce verdi) o inibisce (linea nera) la kinasi 1, la 2 o entrambe. Poichè i segnali convergono sulla kinasi 3, questa rete si attiverà solo quando saranno presenti combinazioni specifiche di stimoli extracellulari.
 o inibisce (linea nera) la kinasi 1, la 2 o entrambe. Poichè i segnali convergono sulla kinasi 3, questa rete si attiverà solo quando saranno presenti combinazioni specifiche di stimoli extracellulari.")
40
La risposta cellulare ai cambiamenti dell’ambiente esterno richiedono spesso l’attivazione coordinata di processi cellulari multipli. In questo caso, l’epinefrina e l’EGF attivano processi intracellulari diversi ma coordinati, aventi come risposta finale l’attivazione metabolica e la proliferazione cellulare.