Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
FILAMENTI DEL CITOSCHELETRO
2012

2
IL CITOSCHELETRO DI UNA CELLULA E’ COSTITUITO DA UNA VARIETA’ DI STRUTTURE FILAMENTOSE

3
Strutture dinamiche e adattabili
Filamenti di actina (microfilamenti): determinano la forma della superficie cellulare e sono necessari per la locomozione dell’intera cellula (lamellipodi, filipodi) Microtubuli: determinano le posizioni degli organelli e dirigono il trasporto intracellulare (formazione del fuso mitotico, cilgia e flagelli) Filamenti intermedi: forniscono forza meccanica e resistenza agli stress (involucro nucleare, assoni) Proteine accessorie sono essenziali per l’assemblaggio controllato dei filamenti del citoscheletro, comprendono i motori proteici che muovono gli organelli o i filamenti stessi. Strutture dinamiche e adattabili Cambiano o persistono per archi di tempo variabili Composte da diverse subunità macromolecolari Le caratteristiche di tali componenti unitarie conferiscono le proprietà del filamento finale Tutti e tre i tipi di filamenti del citoscheletro si formano come complessi elicoidali di subunità che si autoassociano, utilizzando una combinazione di contatti estremità-estremità o fianco-fianco. Differenze nelle forze che tengono unite le subunità determinano differenze cruciali nella stabilità e nelle proprietà meccaniche dei singoli filamenti.
: determinano la forma della superficie cellulare e sono necessari per la locomozione dell’intera cellula (lamellipodi, filipodi) Microtubuli: determinano le posizioni degli organelli e dirigono il trasporto intracellulare (formazione del fuso mitotico, cilgia e flagelli) Filamenti intermedi: forniscono forza meccanica e resistenza agli stress (involucro nucleare, assoni) Proteine accessorie sono essenziali per l’assemblaggio controllato dei filamenti del citoscheletro, comprendono i motori proteici che muovono gli organelli o i filamenti stessi. Strutture dinamiche e adattabili. Cambiano o persistono per archi di tempo variabili. Composte da diverse subunità macromolecolari. Le caratteristiche di tali componenti unitarie conferiscono le proprietà del filamento finale. Tutti e tre i tipi di filamenti del citoscheletro si formano come complessi elicoidali di subunità che si autoassociano, utilizzando una combinazione di contatti estremità-estremità o fianco-fianco. Differenze nelle forze che tengono unite le subunità determinano differenze cruciali nella stabilità e nelle proprietà meccaniche dei singoli filamenti.")
4
CIASCUN TIPO DI FILAMENTO E’ UN POLIMERO
MONOMERO POLIMERO actina > > > microfilamenti tubulina > > > microtubuli diverse proteine > > filamenti intermedi

5
Filamenti di Actina Polimeri elicoidali a 2 filamenti della proteina ACTINA Strutture flessibili 5-9nM Fasci lineari Reti bidimensionali Gel tridimensionaliconcentrati soprattutto nella corteccia

6
Microtubuli Cilindri cavi Unità: tubulina (alfa e beta) 25nM Rigidi
MTOC: centrosoma

7
Filamenti Intermedi Unità: famiglia eterogenea di proteine 10nM
Lamina nucleare, assoni, tessuti epiteliali

8
PROTOFILAMENTI I polimeri del citoscheletro sono costituiti lunghe file di subunità unite per le estremità, che si associano fra loro lateralmente. Tenute insieme da interazioni idrofobiche e legami non covalenti deboli Le posizioni e i tipi di contatti sono diversi per i diversi filamenti.

9
NUCLEAZIONE Assemblaggio di subunitàin un aggregato iniziale (o nucleo) Stabilizzazione dei contatti subunità-subunità Allungamento del polimero per aggiunta di altre subunità

10
NUCLEAZIONE la polimerizzazione dipende dalla concentrazione di subunità non polimeriche Concentrazione critica: quando il tasso di associazione e dissociazione si equivalgono Punto stazionario proteine speciali per catalizzare la nucleazione dei filamenti in siti specifici, determinando la posizione in cui si assemblano nuuovi filamenti del citoscheletro

11
FORMA E MOVIMENTO DELLE CELLULE MITOSI E TRAFFICO DI VESCICOLE
CIASCUN TIPO DI FILAMENTO HA PROPRIETA’ E FUNZIONI SPECIFICHE MICROFILAMENTI (ACTINA) FORMA E MOVIMENTO DELLE CELLULE MICROTUBULI (TUBULINA) MITOSI E TRAFFICO DI VESCICOLE FILAMENTI INTERMEDI IMPALCATURA E ORGANIZZAZIONE DI TESSUTI
 FORMA E MOVIMENTO. DELLE CELLULE. MICROTUBULI (TUBULINA) MITOSI E TRAFFICO. DI VESCICOLE. FILAMENTI INTERMEDI. IMPALCATURA E. ORGANIZZAZIONE DI. TESSUTI.")
12
I 3 TIPI DI FILAMENTI HANNO PROPRIETA’ FISICHE DIVERSE

13
L’ORGANIZZAZIONE DINAMICA DEL CITOSCHELETRO IN VIVO PUO’ ESSERE STUDIATA MEDIANTE L’USO DI PROTEINE FLUORESCENTI TUBULINA-GFP

14
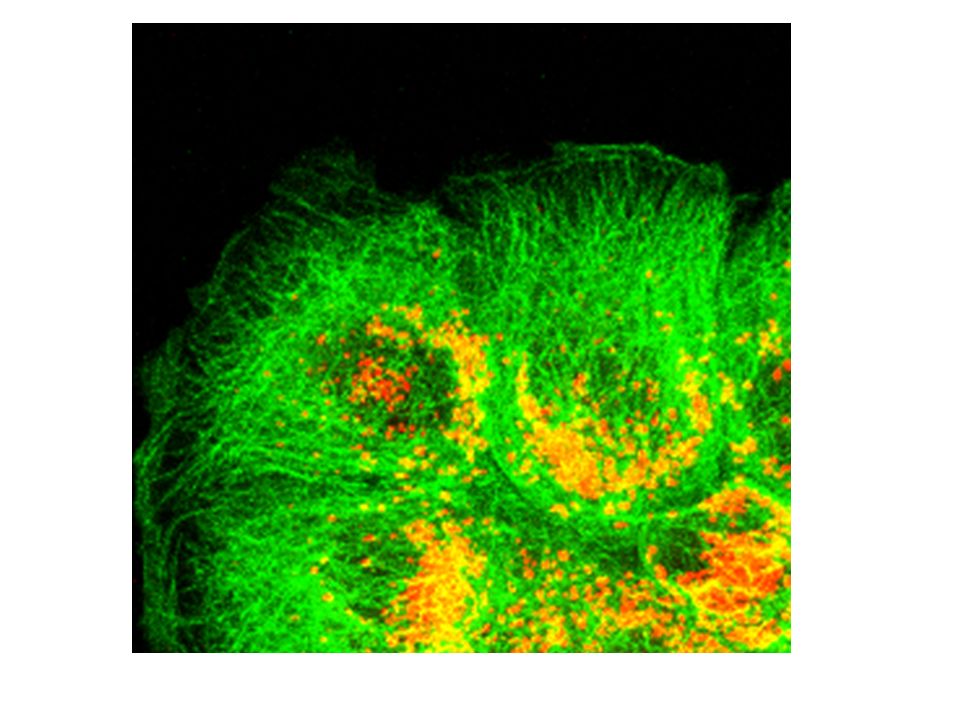
I MICROFILAMENTI SONO VARIAMENTE ORGANIZZATI IN FIBROBLASTI

15
ED IN CELLULE EPITELIALI

17
I microfilamenti

18
IL MONOMERO DI ACTINA LEGA ATP

19
I MONOMERI DI ACTINA NON SONO SIMMETRICI
ESTREMITA’ - COOH NH2 ESTREMITA’ +

20
I MICROFILAMENTI SONO POLIMERI DI ACTINA

21
I FILAMENTI DI ACTINA HANNO UNA POLARITA’
+ FILAMENTO + ATP/ADP - -

22
+ -

24
LA POLIMERIZZAZIONE DEI FILAMENTI DI ACTINA SI REALIZZA ATTRAVERSO 3 FASI

25
LA POLIMERIZZAZIONE HA LUOGO QUANDO LA G-ACTINA E’ AL DI SOPRA DELLA CONCENTRAZIONE CRITICA

26
CERTE CONCENTRAZIONI DI G-ACTINA POSSONO DETERMINARE ‘TREADMILLING’

27
L’ACTINA INTERAGISCE CON MOLTE PROTEINE
LIEVITO

28
MONOMERI SEQUESTRO NUCLEAZIONE TAGLIO POLIMERI FASCI MOVIMENTO FORMAZIONE DI UNA RETE LEGAMI LATERALI BLOCCO DELLE ESTREMITA’

29
Formina Complesso ARP TIMOSINA PROFILINA COFILINA TROPOMIOSINA GELSOLINA Proteina del cappuccio Fimbrina α-actinina Filamina Spettrina ERM

30
ACTINA DOMINI CH CALPONIN HOMOLOGY

31
1 bd 2 o + bd

32
1 PROTEINE CHE CONTROLLANO LA POLIMERIZZAZIONE DEI FILAMENTI
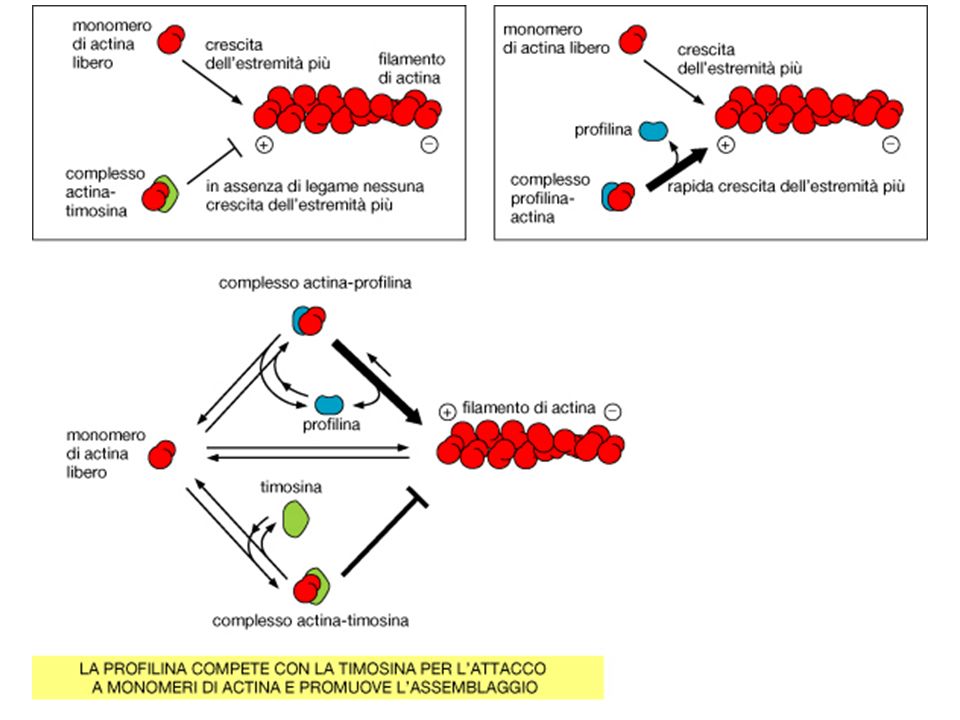
DI ACTINA LEGANDOSI ALLA G-ACTINA. 1 Formina Complesso ARP TIMOSINA PROFILINA

33
TIMOSINA b4 LA TIMOSINA 4 LEGA IL MONOMERO DI ACTINA IMPEDENDO AL MONOMERO DI ASSOCIARSI ALLE ESTREMITA’ DEL FILAMENTO. PROFILINA SI LEGA AL MONOMERO, AL LATO OPPOSTO ALLA FESSURA DI LEGAME PER L’ATP. IL COMPLESSO PROFILINA-ACTINA PUO’ LEGARSI AD UN FILAMENTO DI ACTINA. UN CAMBIO DI CONFORMAZIONE FA DISTACCARE LA PROFILINA. LA PROFILINA COMPETE CON LA TIMOSINA 4.

35
LA PROFILINA E’ FREQUENTEMENTE LEGATA A LIPIDI DI MEMBRANA

36
SEGNALI EXTRACELLULARI POSSONO ATTIVARE LA PROFILINA CHE ANTAGONIZZA LA TIMOSINA 4

37
PROTEINE RICCHE IN PROLINA POSSONO CONTRIBUIRE AD ATTIVARE LOCALMENTE LA PROFILINA

38
LE PROTEINE ARP PROMUOVONO LA NUCLEAZIONE DI FILAMENTI DI ACTINA DALLA MEMBRANA PLASMATICA

39
PROTEINE CHE CONTROLLANO LA LUNGHEZZA DEI FILAMENTI DI ACTINA MEDIANTE FRAMMENTAZIONE O INCAPPUCCIAMENTO. 2 COFILINA TROPOMIOSINA GELSOLINA Proteina del cappuccio

40
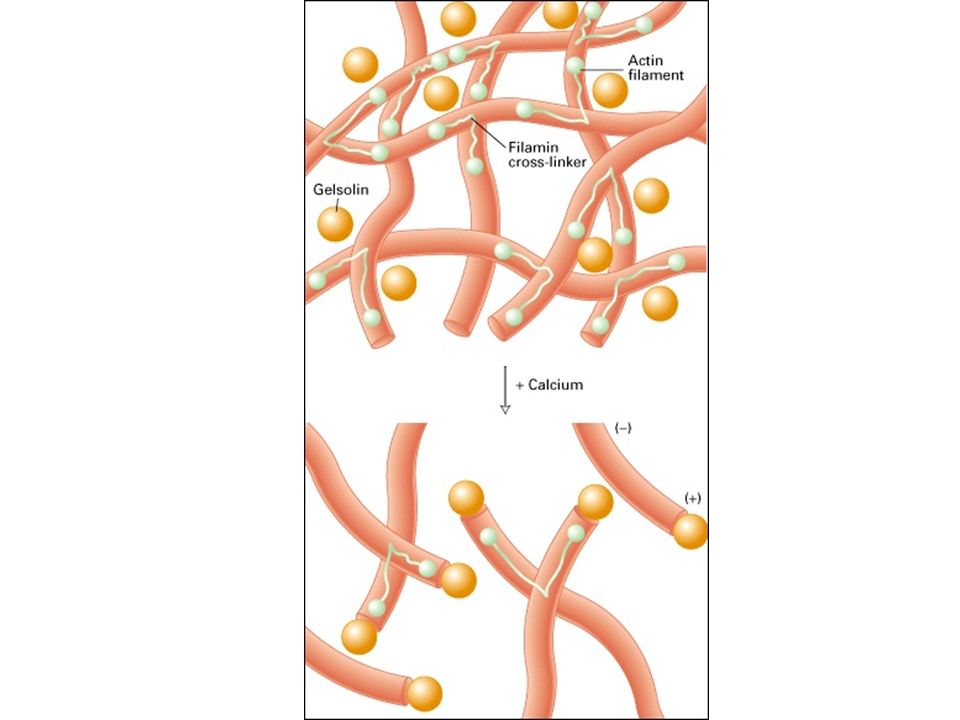
LA GELSOLINA TAGLIA ED INCAPPUCCIA
I FILAMENTI DI ACTINA

41
LE PROTEINE CAPPING INCAPPUCCIANO UNA ESTREMITA’ DEI FILAMENTI DI ACTINA
(tropomodulina, etc.) (capZ, etc.)
 (capZ, etc.)")
42
PROTEINE CHE DETERMINANO IL TIPO DI ORGANIZZAZIONE DEI FILAMENTI DI ACTINA.
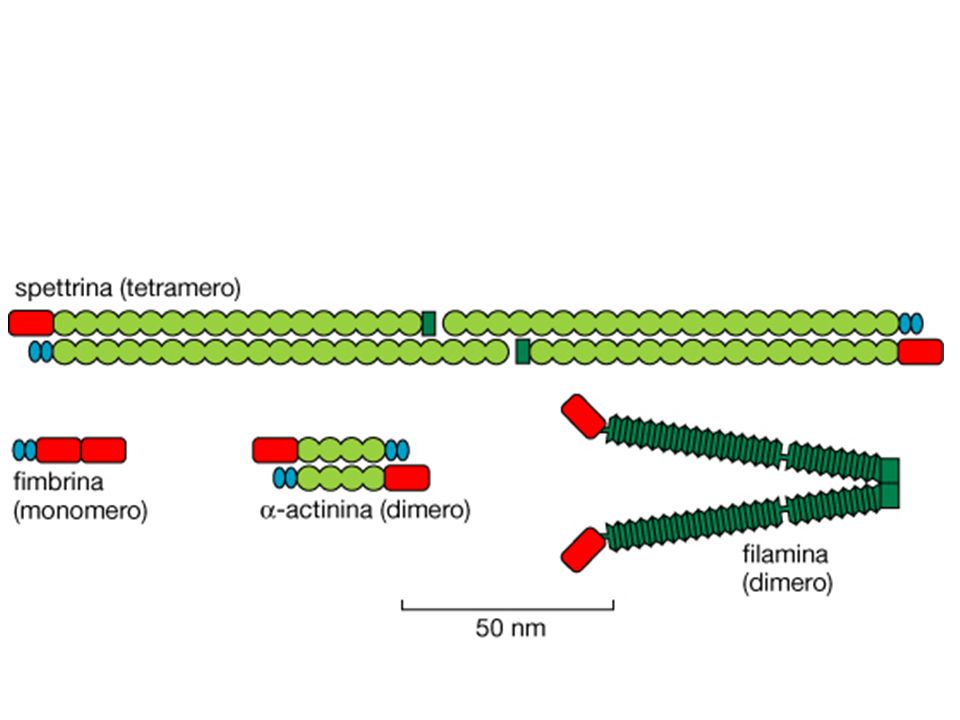
3 Fimbrina α-actinina Filamina Spettrina ERM

43
-ACTININA E FIMBRINA PROMUOVONO LA FORMAZIONE DI FASCI DI ACTINA

44
FASCINA PROMUOVE LA FORMAZIONE DI FASCI DI ACTINA

45
LA FILAMINA INDUCE LA FORMAZIONE DI UN GEL
actina filamina

46
dimero di filamina actina
IL DIMERO DI FILAMINA LEGA DUE FILAMENTI DI ACTINA dimero di filamina actina

47
FILAMINA E TROPOMIOSINA SI LEGANO ALLA ACTINA IN MODO ESCLUSIVO

49
ALCUNE PROTEINE PROMUOVONO L’INTERAZIONE
DI FILAMENTI DI ACTINA CON LA MEMBRANA.

50
ALCUNE PROTEINE PROMUOVONO L’INTERAZIONE
DI FILAMENTI DI ACTINA CON LA MEMBRANA.

51
MICROVILLI

53
lamellipodio filopodio fibre da stress cortex

54
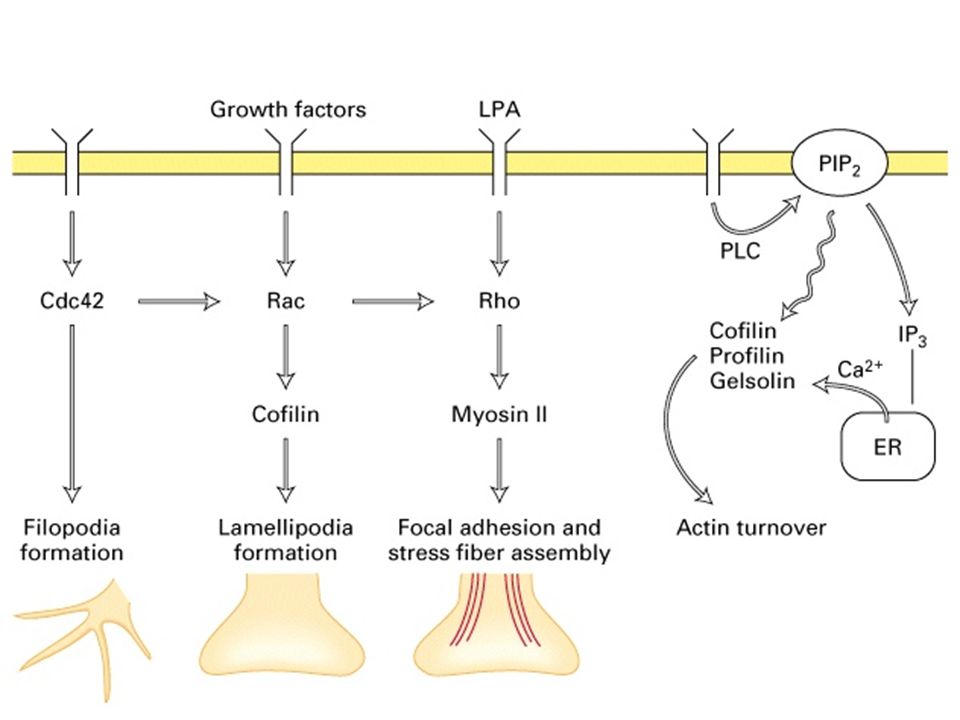
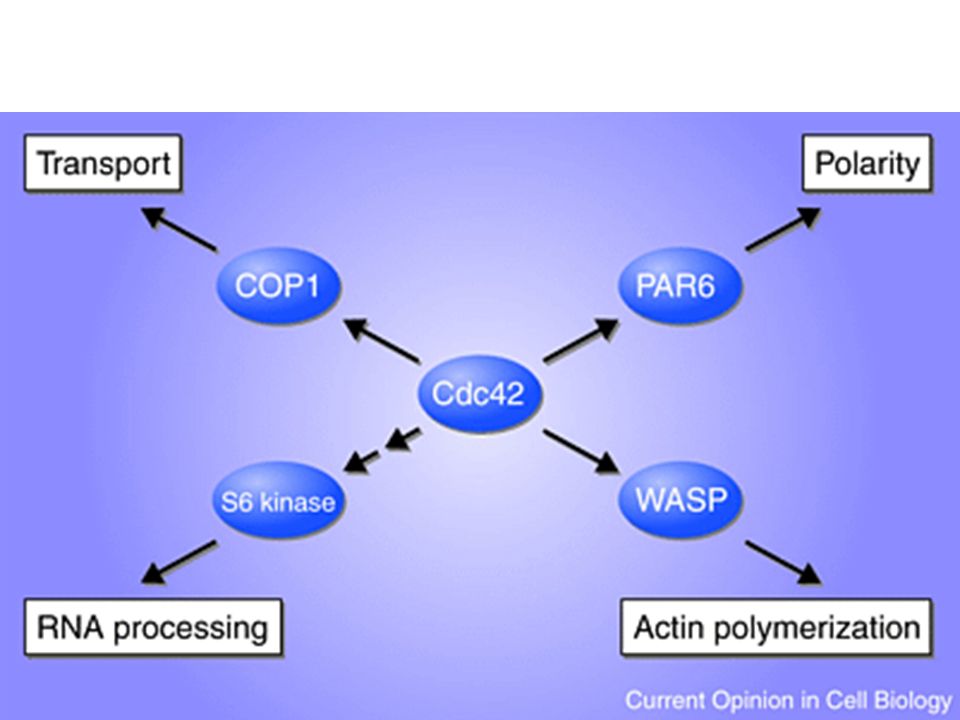
Cosa sono le piccole GTPasi?
Le piccole GTPasi sono proteine monomeriche che appartengono ad una famiglia il cui capostipite è ras. Le GTPasi che sono essenziali nella trasduzione di segnali.

55
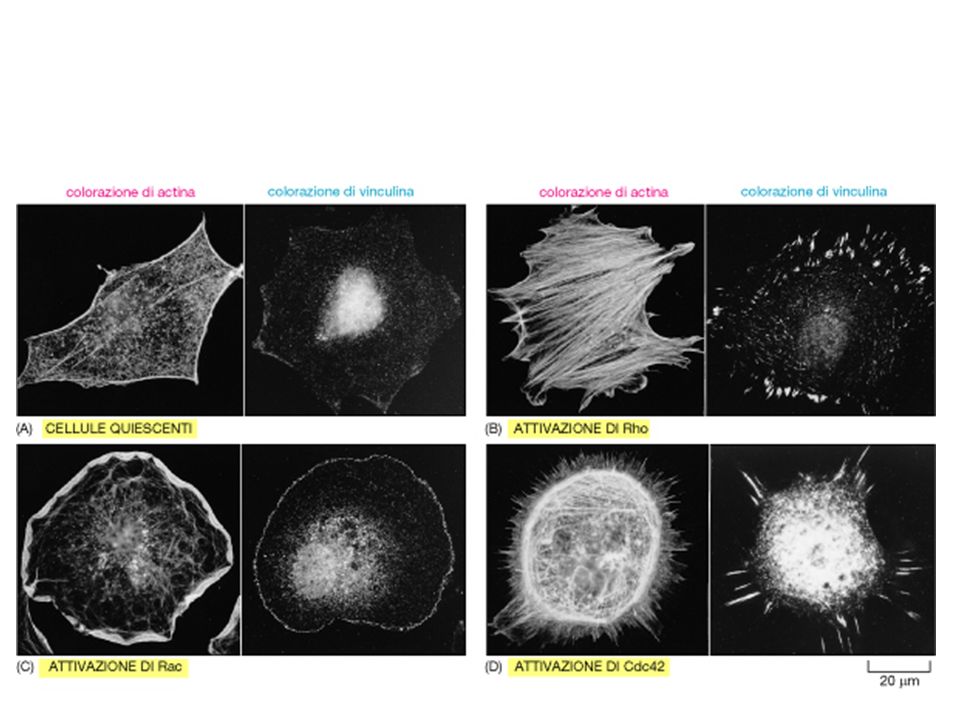
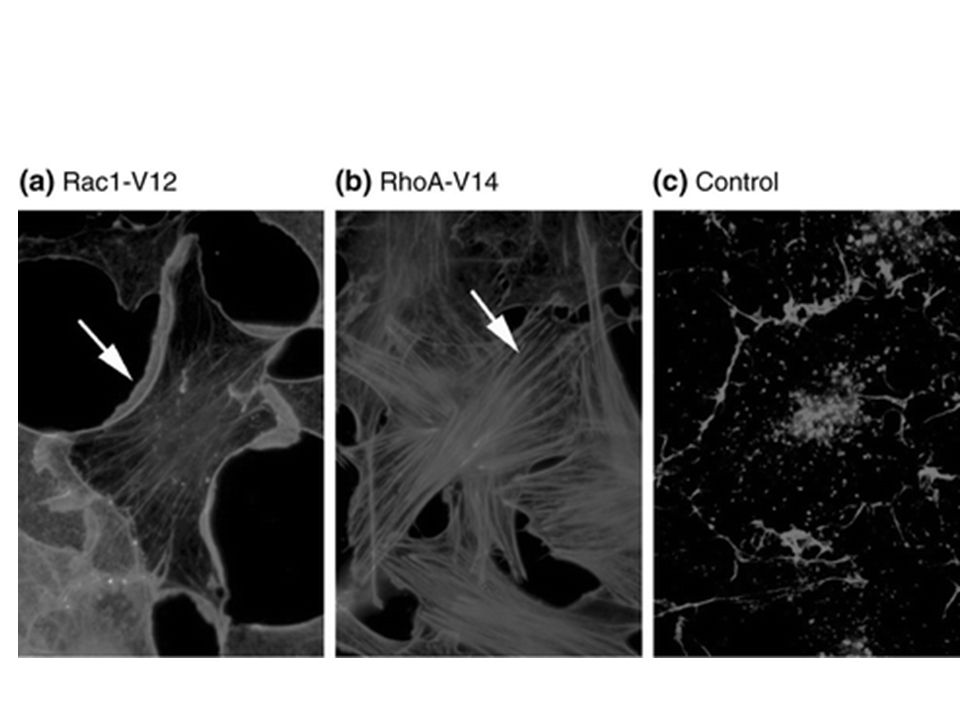
Rho - fibre da stress Rac - lamellipodi Cdc42 - filopodi

59
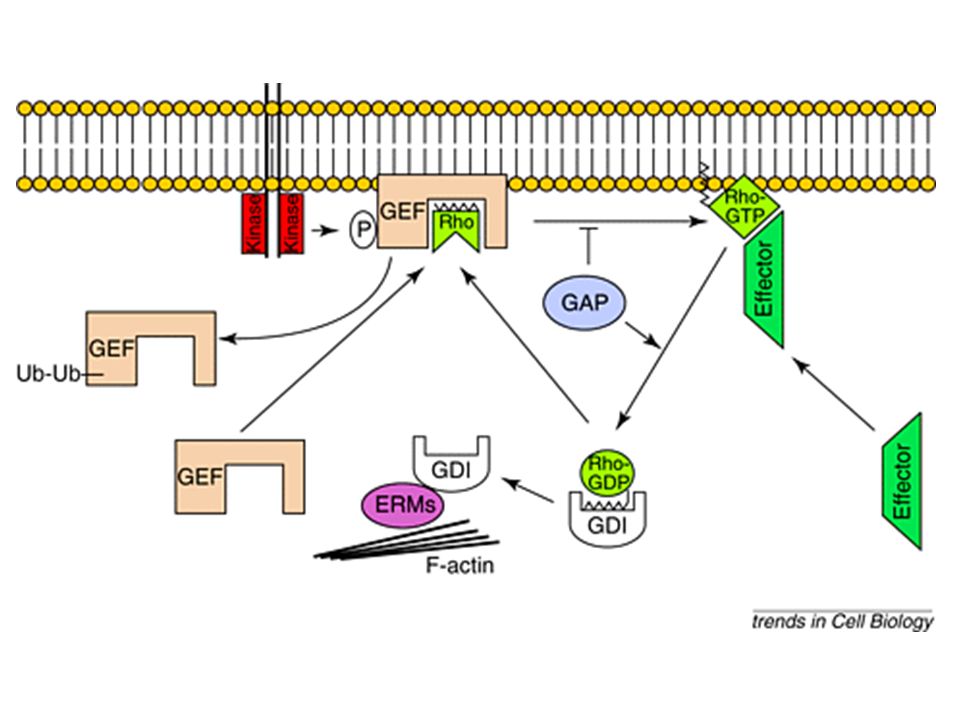
Come viene attivata la proteina Rho?
> GDP GTP GEF RhoA GDP inattivo RhoA GTP attivo Pi GAP

62
Rho A è regolato così

63
Rho induce così la formazione di fibre da stress

64
WISKOTT-ALDRICH SINDROME (WAS)
")
65
DROGHE CHE STABILIZZANO O DESTABILIZZANO
I MICROFILAMENTI CITOCALASINE FALLOIDINA CNF

66
CNF treated FRT-b1B cells organize microfilaments

Presentazioni simili


.>")










>")


>")
.>")

