Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Lezione 5 DNA struttura e replicazione Cromosomi

2
Natura del materiale genetico
Nei primi decenni del ‘900 si raccolgono evidenze che i cromosomi sono costituiti da DNA e proteine. - Qual è la molecola responsabile dell’informazione biologica? - Qual è la sua struttura? Esperimenti di importanza fondamentale (Griffith, Avery et al., Hershey et al.) vengono effettuati negli anni ’30-50 su organismi semplici, quali batteri e batteriofagi e dimostrano che il materiale genetico è il DNA Aver capito la sua struttura ha permesso di capire come avviene la sua replicazione
 vengono effettuati negli anni ’30-50 su organismi semplici, quali batteri e batteriofagi e dimostrano che il materiale genetico è il DNA. Aver capito la sua struttura ha permesso di capire come avviene la sua replicazione.")
3
Regola di Chargaff 1950 E. Chargaff scopre che nel DNA di specie diverse c’è proporzionalità nella composizione in basi azotate Organismo A% T% G% C% Pneumococco 29.8 31.6 20.5 18.0 Lievito 31.3 32.9 18.7 17.1 Riccio di mare 32.8 32.1 17.7 18.4 E.coli 26 24.9 24.8 25.2 Homo s. 30.9 29.4 19.9 19.8 Rattus n. 28.6 28.4 21.4 21.5 A=T, C=G La quantità totale delle purine (A+G) è uguale a quella delle pirimidine (T+C)
 è uguale a quella delle pirimidine (T+C)")
4
Cristallografia a raggi X (1952 R. Franklin)
M. Wilkins I raggi X vengono deviati dalla loro traiettoria quando colpiscono il campione. L’ immagine di diffrazione ottenuta è compatibile con una struttura elicoidale.

5
Nel 1953 James Watson e Francis Crick, basandosi sulle osservazioni di cristallografia ai raggi X di Rosalind Franklin e sull’osservazione di Erwin Chargaff, propongono il modello a doppia elica per la molecola di DNA

6
Il DNA è un polimero di nucleotidi
Nucleotide: molecola formata da un gruppo fosfato (1), uno zucchero pentoso (desossiribosio) (2) e una base azotata (3) pirimidine purine 5’ Adenina, citosina, guanina, timina 3’
, uno zucchero pentoso (desossiribosio) (2) e una base azotata (3) pirimidine. purine. 5’ Adenina, citosina, guanina, timina. 3’")
7
Modello a doppia elica del DNA
Il DNA ha una struttura ad elica (in accordo coi dati di cristallografia) a doppio filamento Gli scheletri zucchero-fosfato sono posizionati esternamente, mentre le basi sono dirette verso l’interno e si appaiano (tramite legami idrogeno) alle basi “complementari” del filamento opposto in accordo con quanto osservato da Chargaff (in tutti i DNA % A = % T; % C = % G). L’appaiamento è sempre purina-pirimidina e questo mantiene il diametro dell’elica uniforme I due filamenti sono antiparalleli 2nm
 a doppio filamento. Gli scheletri zucchero-fosfato sono posizionati esternamente, mentre le basi sono dirette verso l’interno e si appaiano (tramite legami. idrogeno) alle basi complementari del filamento. opposto in accordo con quanto osservato da. Chargaff (in tutti i DNA % A = % T; % C = % G). L’appaiamento è sempre purina-pirimidina. e questo mantiene il diametro dell’elica uniforme. I due filamenti sono antiparalleli. 2nm.")
8
Il diametro si mantiene costante perché le purine si appaiano alle pirimidine
2nm G ≡ C: 3 legami H A = T: 2 legami H

9
Il modello prefigurava genialmente una modalità di replicazione semiconservativa che verrà poi sperimentalmente dimostrata da Meselson e Stahl nel 1958. Ogni filamento dell’elica funziona da stampo per la sintesi di un nuovo filamento, sfruttando la complementarietà delle basi.

10
Modelli ipotetici di replicazione del DNA
Filamento parentale Filamento nuovo Alla fine del processo di sintesi potremmo ottenere: due molecole di DNA ciascuna costituita a un filamento di nuova sintesi e da un filamento originario (semiconservativa) una molecola di DNA costituita da due filamenti parentali e l’altra da due filamenti di nuova sintesi (conservativa) due molecole di DNA costituite entrambe da un’alternanza di filamenti nuovi e parentali (dispersiva)
 una molecola di DNA costituita da due filamenti parentali e l’altra da due filamenti di nuova sintesi (conservativa) due molecole di DNA costituite entrambe da un’alternanza di filamenti nuovi e parentali (dispersiva)")
11
La replicazione del DNA è semiconservativa
Ciascuno dei due filamenti parentali serve da stampo per la sintesi di un nuovo filamento e le due nuove doppie eliche sono costituite da ognuna da un filamento vecchio e da un filamento nuovo

12
Tutte le DNA polimerasi in tutti gli organismi sono capaci di sintetizzare DNA soltanto in direzione 5’-3’ DNA polimerasi DNA dipendente 5’ 3’ Direzione di sintesi

13
La DNA polimerasi allunga il filamento nascente aggiungendo un nucleotide all’estremità 3’OH
5’ 5’ 3’ 3’

14
Polimerizzazione del DNA

15
Direzione di sintesi dei due filamenti
5’ 3’ Poiché le polimerasi possono estendere solo l’estremità 3’, mentre entrambi i filamenti servono da stampo per la sintesi di nuovi filamenti, la sintesi deve procedere sui due filamenti stampo in direzione opposta 5’ 3’ 5’ 3’ 5’ 3’

16
Replicazione del DNA 1 In corrispondenza delle origini di replicazione l’elicasi svolge i filamenti parentali della doppia elica rompendo i legami H tra le basi e generando due filamenti singoli Le ssbp (single strand binding proteins) legano il singolo filamento e lo stabilizzano Origin of replication Denaturazione del DNA helicase Questo schema (con questi enzimi) vale per i procarioti
 legano il singolo filamento e lo stabilizzano. Origin of replication. Denaturazione del DNA. helicase. Questo schema (con questi enzimi) vale per i procarioti.")
17
Replicazione del DNA 2 innesco
La primasi (RNA polimerasi-DNA dipendente) sintetizza un breve innesco (primer) a RNA (le DNA pol non sono in grado di iniziare ex novo la sintesi, ma solo di allungare un 3’ preesistente) La DNA polimerasi III estende l’innesco a RNA sintetizzando DNA innesco 3’ helicase helicase
 sintetizza un breve innesco (primer) a RNA (le DNA pol non sono in grado di iniziare ex novo la sintesi, ma solo di allungare un 3’ preesistente) La DNA polimerasi III estende l’innesco a RNA sintetizzando DNA. innesco. 3’ helicase. helicase.")
18
La DNA polimerasi procede, sul filamento guida (leading strand), nella stessa direzione di avanzamento della elicasi e quindi in maniera continua … Ma cosa succede sul filamento complementare? Leading strand helicase

19
Il filamento ritardato (lagging strand) viene sintetizzato in maniera discontinua (la DNApol III va in direzione opposta rispetto all’elicasi), attraverso la sintesi di una serie di frammenti (fr. di Okazaki) 5’ 3’ 5’ helicase primase

20
Replicazione del DNA 3 Rimozione del primer a RNA e “gap filling” da parte della DNA pol I che attacca nucleotidi al 3’ libero del frammento di Okazaki precedente mentre degrada il primer a RNA DNApolI ha attività RNasica? Si nei procarioti ( Negli eucarioti c’è RNase H 5’ helicase

21
Azione della DNA pol I Primer RNA 5’ 5’ 3’ 3’ 5’ 3’ 5’ 5’ 3’ 3’
DNApolI ha attività RNasica? Si nei procarioti ( Negli eucarioti c’è RNase H 5’ 5’ 3’ 3’ DNA pol I 5’ 3’

22
Replicazione del DNA 4 Saldatura dei frammenti (ligasi) 5’ ligase
helicase ligase

23
Replicazione del DNA È la DNA pol I, non la II a rimuovere i primers

24
La separazione dei filamenti di una struttura elicoidale genera dei superavvolgimenti
Topoisomerasi I Rompono transitoriamente una sola delle catene del DNA, la ruotano attorno a quella integra e infine riuniscono le estremità interrotte Le Topoisomerasi hanno il compito di rilassare (eliminare) le tensioni che si creano nella doppia elica del DNA in seguito ai processi cellulari che implichino un cambiamento di topologia del DNA (trascrizione, replicazione del DNA)
 le tensioni che si creano nella doppia elica del DNA in seguito ai processi cellulari che implichino un cambiamento di topologia del DNA (trascrizione, replicazione del DNA)")
25
Replicazione del DNA circolare
ORI è sequenza ricca in A-T Nei procarioti l’origine di replicazione è singola Direzione di replicazione bidirezionale a partire da ORI singola Origins tend to be "AT-rich" (rich in adenine and thymine bases) to assist this process, because A-T base pairs have two hydrogen bonds (rather than the three formed in a C-G pair)—in general, strands rich in these nucleotides are easier to separate because a greater number of hydrogen bonds requires more energy to break them
 to assist this process, because A-T base pairs have two hydrogen bonds (rather than the three formed in a C-G pair)—in general, strands rich in these nucleotides are easier to separate because a greater number of hydrogen bonds requires more energy to break them.")
26
Replicazione del DNA lineare
Le lunghe molecole di DNA lineare dei cromosomi eucarioti hanno origini di replicazione multiple Direzione di replicazione bidirezionale a partire da ORI multiple DNA pol delta e epsilon (l’analogo di DNA pol III negli eucarioti)
")
27
Necessità di ORI multiple negli eucarioti
Le lunghe molecole di DNA lineare dei cromosomi eucarioti hanno origini di replicazioni multiple Le DNA polimerasi eucariotiche hanno una velocità di incorporazione di 75 nucleotidi al secondo La lunghezza media di un cromosoma umano è 1.4 x 108 bp (3.2 x 109 bp genoma aploide/ 23 cromosomi) Se ci fosse una singola ORI, per duplicare un singolo cromosoma medio occorrerebbero più di 10 giorni In realtà la fase S dura circa 8 ore 10 giorni considerando un ORI interno al cromosoma, da cui si dipartono due forche replicative
 Se ci fosse una singola ORI, per duplicare un singolo cromosoma medio occorrerebbero più di 10 giorni. In realtà la fase S dura circa 8 ore giorni considerando un ORI interno al cromosoma, da cui si dipartono due forche replicative.")
28
Il cromosoma batterico ha un’unica ORI
Il genoma di E. coli è lungo 4.6x10 6 bp La velocità dellla Pol III arriva fino a 1000 nt/ sec Per replicare il cromosoma occorrono: 4.6x10 6 bp = 70 min c.a 1000nt/sec su ogni lato della forca replicativa quindi 35 min c.a

29
Caratteristiche delle DNA polimerasi
Hanno attività polimerasica SOLO in direzione 5’-3’ Necessità di innesco a RNA (primer) (non possono iniziare dal nulla la sintesi di una nuova catena) Attività esonucleasica 3’-5’ correzione di bozze
 (non possono iniziare dal nulla la sintesi di una nuova catena) Attività esonucleasica 3’-5’ correzione di bozze.")
30
Correzione di bozze Alla DNA polimerasi è associata un’attività esonucleasica che consente la “correzione di bozze”= rimozione di nucleotidi errati. Con questo sistema vengono corretti gli errori di appaiamento commessi dalla DNA pol mentre la replicazione è ancora in corso. -tasso di errore durante la sintesi 1 su nt -dopo correzione 1su dieci miliardi nt È sufficientemente basso?

31
Tasso di mutazione e correzione degli errori
Nel genoma umano (aploide) ci sono c.a. 3 x109 nt di questi solo 6 x 107 nt (il 2%) sono sequenze codificanti (esoniche) Se non ci fosse correzione di bozze (tasso di errore durante la sintesi 1x10-4 nt): 6000 mutazioni cadrebbero in sequenze codificanti a ogni replicazione del DNA Dopo correzione di bozze (tasso di errore 1x10-10 nt ): 0.006 mutazioni cadono in sequenze codificanti a ogni replicazione del DNA (1 mutazione ogni 166 replicazioni) Sono comunque maggiori di quelle osservate nella cellula, grazie all’intervento di processi di riparazione del DNA post-replicativi
 ci sono c.a. 3 x109 nt. di questi solo 6 x 107 nt (il 2%) sono sequenze codificanti (esoniche) Se non ci fosse correzione di bozze (tasso di errore durante la sintesi 1x10-4 nt): 6000 mutazioni cadrebbero in sequenze codificanti a ogni replicazione del DNA. Dopo correzione di bozze (tasso di errore 1x10-10 nt ): mutazioni cadono in sequenze codificanti a ogni replicazione del DNA (1 mutazione ogni 166 replicazioni) Sono comunque maggiori di quelle osservate nella cellula, grazie all’intervento di processi di riparazione del DNA post-replicativi.")
32
Riparazione degli appaiamenti errati (mismatch repair)
Provvede ad effettuare una scansione del DNA dopo la replicazione alla ricerca di appaiamenti errati Il sistema di riparazione è in grado di riconoscere e riparare il filamento di nuova sintesi Mismatch repair is strand-specific. During DNA synthesis only the newly synthesised (daughter) strand will include errors, and replacing a base in the parental strand would actually introduce an error. The mismatch repair machinery has a number of cues which distinguish the newly synthesised strand from the template (parental). In gram-negative bacteria transient hemimethylation distinguishes the strands (the parental is methylated and daughter is not). In other prokaryotes and eukaryotes the exact mechanism is not clear. Mismatch repair (MMR), which corrects errors of DNA replication and recombination that result in mispaired (but undamaged) nucleotides.
 strand will include errors, and replacing a base in the parental strand would actually introduce an error. The mismatch repair machinery has a number of cues which distinguish the newly synthesised strand from the template (parental). In gram-negative bacteria transient hemimethylation distinguishes the strands (the parental is methylated and daughter is not). In other prokaryotes and eukaryotes the exact mechanism is not clear. Mismatch repair (MMR), which corrects errors of DNA replication and recombination that result in mispaired (but undamaged) nucleotides.")
33
Difetti nel “mismatch repair”
tasso di errore durante la sintesi 1x10-4 nt dopo correzione 1x10-10 nt …è sufficientemente basso? Difetti nel “mismatch repair” Mutazioni nei geni che codificano per gli enzimi coinvolti nel mismatch repair sono associate al cancro. La cellula non riesce a riparare le mutazioni che si accumulano in tutto il genoma e che, quando colpiscono geni che regolano la proliferazione cellulare, inducono la trasformazione cancerosa (es: cancro del colon non poliposico ereditario HNPCC, autosomica dominante)
")
34
Un errore non corretto viene poi perpetuato nei cicli di replicazione successivi

35
Cromosomi

36
Cromatina All’interno del nucleo della cellula eucariotica si trova la cromatina (DNA genomico + proteine) La cromatina appare organizzata in entità separate (cromosomi) quando la cellula è in mitosi interfase tarda profase inizio anafase
 quando la cellula è in mitosi. interfase. tarda profase. inizio anafase.")
37
Cromosomi Cromosomi metafasici altamente condensati braccio p
centromero braccio q P petit, q è semplicemente la lettera che segue la p According to the NCBI, "q" refers to the French word "queue" meaning 'tail' Ciascun cromosoma, dopo la fase S, è costituito da due cromatidi fratelli, tenuti insieme dal centromero

38
Telomeri 1 Sequenze ripetute all’estremità dei cromosomi costituiti da alcune migliaia di ripetizioni di sequenze brevi [nell’uomo: (TTAGGG)n] Proteggono il cromosoma dalla degradazione ad opera di nucleasi (sono ripiegati su se stessi) ed impediscono che le estremità dei cromosomi si saldino tra di loro I telomeri sono ripiegati su se stessi per proteggerli dall’azione delle esonucleasi
![Telomeri 1 Sequenze ripetute all’estremità dei cromosomi costituiti da alcune migliaia di ripetizioni di sequenze brevi [nell’uomo: (TTAGGG)n]](http://slideplayer.it/slide/12319625/73/images/38/Telomeri+1+Sequenze+ripetute+all%E2%80%99estremit%C3%A0+dei+cromosomi+costituiti+da+alcune+migliaia+di+ripetizioni+di+sequenze+brevi+%5Bnell%E2%80%99uomo%3A+%28TTAGGG%29n%5D.jpg "Proteggono il cromosoma dalla degradazione ad opera di nucleasi (sono ripiegati su se stessi) ed impediscono che le estremità dei cromosomi si saldino tra di loro. I telomeri sono ripiegati su se stessi per proteggerli dall’azione delle esonucleasi.")
39
Telomeri 2 Nelle cellule germinali e nelle cellule staminali la lunghezza dei telomeri rimane costante ad ogni divisione cellulare grazie all’attività della telomerasi Nelle cellule somatiche differenziate ad ogni replicazione del DNA il telomero subisce un accorciamento La riduzione dei telomeri dopo n divisioni provoca arresto della crescita cellulare e apoptosi Effetti «ambientali» sulla lunghezza del telomero: lo stress, il fumo accorciano i telomeri, fare sport li mantiene lunghi (

40
Telomeri e senescenza

42
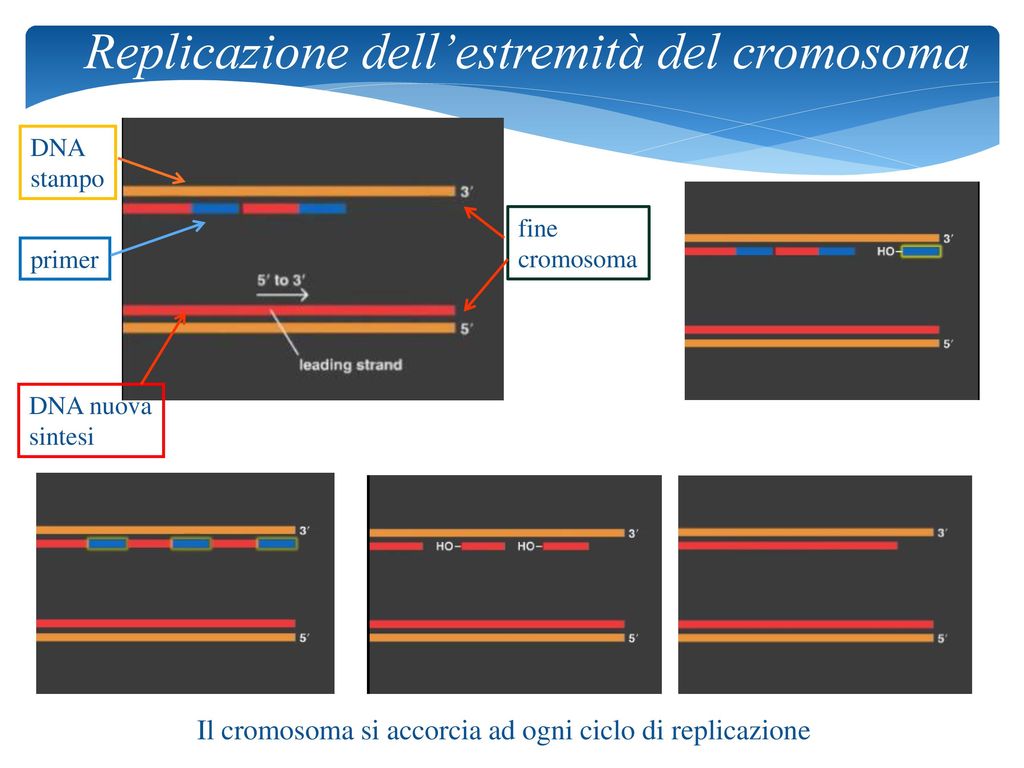
Replicazione dell’estremità del cromosoma
DNA stampo fine cromosoma primer DNA nuova sintesi Il cromosoma si accorcia ad ogni ciclo di replicazione

43
Centromero È la regione del cromosoma in cui i cromatidi fratelli sono uniti Sequenza ripetute In corrispondenza del centromero ci sono due cinetocori (uno per cromatidio) che servono ad agganciare il cromosoma alle fibre del fuso
 che servono ad agganciare il cromosoma alle fibre del fuso.")
44
Il DNA genomico degli eucarioti è lunghissimo!
Distanza tra due coppie di basi 0.34nm 3x109 bp (numero di basi di un genoma aploide umano) x 0.34nm (distanza tra due coppie di basi) = 1 metro (lunghezza di un genoma umano aploide) Deve essere compattato in un nucleo di pochi m!!!
 x 0.34nm (distanza tra due coppie di basi) = 1 metro (lunghezza di un genoma umano aploide) Deve essere compattato in un nucleo di pochi m!!!")
45
Nucleosoma L’unità fondamentale della cromatina è il nucleosoma: tratto di DNA (146bp) avvolto con c.a. due giri attorno a un core proteico formato da un ottamero istonico Istoni: proteine cariche positivamente (arg, lys) che interagiscono coi gruppi fosfato del DNA Linker c.a. 60 bp 11 nm 2 nm
 avvolto con c.a. due giri attorno a un core proteico formato da un ottamero istonico. Istoni: proteine cariche positivamente (arg, lys) che interagiscono coi gruppi fosfato del DNA. Linker c.a. 60 bp. 11 nm. 2 nm.")
46
Nucleosoma (1.65 giri,146 pb) H2A H2B H4 H3 11 nm H3 H2A Dimensioni istone: 6 x 11 nm DNA linker + DNA avvolto sull’ottamero = 200bp c.a. 0.34 nm x 200bp = 70nm quindi il livello di compattamento nella fibra da 10nm è di 7X Nella fibra da 30nm ci sono 6 nucleosomi, quindi l’impaccamento è di 40X Le anse sono lunghe nt H4 Ottamero istoni: 2 molecole di H2A, H2B, H3 e H4

47
Istone H1 e fibra da 30nm L’istone H1 (di giunzione) ha la funzione di compattare ulteriormente i nucleosomi Promuove la formazione della fibra solenoide da 30nm 11 nm H1

48
Il cromosoma metafasico presenta un alto grado di compattazione della cromatina
Le anse sono lunghe nt

49
Eucromatina-eterocromatina 1
Eterocromatina è più compatta e trascrizionalmente inattiva Eucromatina interfasica trascrizionalmente attiva; si trova allo stato di cromatina distesa ed è accessibile agli enzimi della trascrizione

50
Eucromatina-eterocromatina 2
Eucromatina: frazione nucleare trascrizionalmente attiva e a bassa densità Eterocromatina: frazione nucleare trascrizionalmente inattiva e densa

51
Cromosoma X e compensazione del dosaggio genico nei mammiferi
maschio livello di espressione femmina (quantità mRNA) gene1 gene1 Nella femmina un cromosoma X viene inattivato per equiparare il dosaggio genico (livello di espressione dei geni che mappano sull’X) a quello del maschio inattivo
 gene1. gene1. Nella femmina un cromosoma X viene inattivato per equiparare il dosaggio genico (livello di espressione dei geni che mappano sull’X) a quello del maschio. inattivo.")
52
Inattivazione cromosoma X
Nelle femmine di mammifero uno dei due cromosomi X è inattivato ed è sempre condensato. Il cromosoma X inattivo è visibile nel nucleo interfasico di cellule ♀ come una masserella di cromatina condensata (eterocromatina), addossata alla membrana nucleare. Viene chiamata cromatina sessuale o corpo di Barr Nuclei di cellule della mucosa orale di ♀ (sinistra) e ♂ (destra)
, addossata alla membrana nucleare. Viene chiamata cromatina sessuale o corpo di Barr. Nuclei di cellule della mucosa orale di ♀ (sinistra) e ♂ (destra)")
53
Cariotipo Il cariotipo di una cellula eucariota è costituito dal numero e dalla morfologia (posizione del centromero e dimensioni) dei suoi cromosomi metafasici
 dei suoi cromosomi metafasici.")
54
Il cariotipo umano Nel 1955 Tjio e Levan (analizzando centinaia di metafasi in 5 tessuti provenienti da 7 individui) dimostrano che il numero esatto di cromosomi umani è 46
 dimostrano che il numero esatto di cromosomi umani è 46.")
55
Cromosomi umani 1→ 22 sono uguali nel maschio e nella femmina (autosomi) La 23esima coppia è quella dei cromosomi sessuali: XX nella femmina (sesso omogametico) XY nel maschio (sesso eterogametico) Determinano il sesso dell’individuo Negli uccelli il sesso omogametico è quello maschile (ZZ), mentre quello femminile è eterogametico (ZW)
 XY nel maschio. (sesso eterogametico) Determinano il sesso dell’individuo. Negli uccelli il sesso omogametico è quello maschile (ZZ), mentre quello femminile è eterogametico (ZW)")
56
Sistema di determinazione del sesso sistema biologico che determina lo sviluppo delle caratteristiche sessuali in un individuo ambientale es. temperatura (Testudo), CO2, O2, umidità, fotoperiodo (Gammarus) genica (asparago: mm Mm ) Cromosomica (cromosoma X e Y, W e Z negli uccelli) Anellide Batteri parassiti di Wolbachia cambiano il sesso del loro ospite quando si tratta di un maschio perché hanno bisogno di svilupparsi nelle uova delle femmine
, CO2, O2, umidità, fotoperiodo (Gammarus) genica (asparago: mm Mm ) Cromosomica (cromosoma X e Y, W e Z negli uccelli) Anellide. Batteri parassiti di Wolbachia cambiano il sesso del loro ospite quando si tratta di un maschio perché hanno bisogno di svilupparsi nelle uova delle femmine")
57
Denaturazione e rinaturazione in vitro del DNA

58
Rinaturazione in presenza di una sonda
rinaturazione sonda denaturazione La sonda è una sequenza di DNA complementare a una determinata sequenza di interesse, che viene marcata con radioattivo o con fluorocromi

59
Ibridazione del cromosoma con sonde fluorescenti
Fluorescenza: emissione luce dopo irraggiamento con UV o anche luce visibile Cromosomi metafasici su vetrino; Denaturazione del DNA, rinaturazione in presenza di sonda fluorescente specifica per una determinata regione del cromosoma

60
telomero …AATCCCAATCC sonda TTAGGG*

61
Fluorescent In Situ Hybridization

62
Origine evolutiva del cromosoma 2 umano

63
Origine evolutiva del cromosoma 2 umano: fusione di cr 12 e cr 13 di pan troglodites
Sequenze telomeriche Sequenze telomeriche il cromosoma 2 deriva dalla fusione dei due cromosomi di scimpanzè o questi ultimi derivano dalla rottura del cromosoma 2? Dato che nei gorilla e negli oranghi il numero di cromosomi è uguale a quello degli scimpanzé, e cioè 24 coppie, la prima opzione risulta più probabile.

Presentazioni simili





>")













