Scaricare la presentazione
1
Sistema Endocrino

2
Il Sistema Endocrino è costituito da GHIANDOLE ENDOCRINE che secerno ORMONI
Gli ormoni (dal greco ormao= eccitare) sono dei messaggeri chimici che vengono riversati direttamente nel sangue per essere trasportati ad un organo bersaglio (target) dove esercitano la loro funzione Caratteristiche degli ormoni: Sono immessi direttamente nel circolo sanguigno Sono trasportati dal sangue alle cellule o organi bersaglio Si legano a recettori specifici presenti nelle cellule bersaglio Un ormone può esplicare effetti molteplici sullo stesso tessuto bersaglio
 sono dei messaggeri chimici che vengono riversati direttamente nel sangue per essere trasportati ad un organo bersaglio (target) dove esercitano la loro funzione. Caratteristiche degli ormoni: Sono immessi direttamente nel circolo sanguigno. Sono trasportati dal sangue alle cellule o organi bersaglio. Si legano a recettori specifici presenti nelle cellule bersaglio. Un ormone può esplicare effetti molteplici sullo stesso tessuto bersaglio.")
3
Le funzioni biologiche degli ormoni si svolgono con tre diversi meccanismi:
Endocrino = l’ormone prodotto a livello della ghiandola endocrina raggiunge il tessuto bersaglio tramite il torrente circolatorio Paracrino = l’ormone prodotto a livello della ghiandola endocrina raggiunge il tessuto bersaglio tramite il liquido extra-cellulare Autocrino = l’ormone prodotto a livello della ghiandola endocrina ha effetto sulle stesse cellule che l’hanno prodotto Classificazione degli ormoni: Ormoni peptidici (costituiti da tre o più amminoacidi) Ormoni steroidei (derivanti dal colesterolo) Ormoni amminici (derivanti da un solo amminoacido)
 Ormoni steroidei (derivanti dal colesterolo) Ormoni amminici (derivanti da un solo amminoacido)")
4
Sintesi Gli ormoni peptidici vengono sintetizzati e immagazzinati in vescicole secretorie Preproormone proormone ormone attivo (contenuto in vescicole secretorie) - input esocitosi
 - input esocitosi.")
5
Gli ormoni steroidei sono lipofili e non sono immagazzinati in vescicole secretorie
Stimolo precursore convertito in ormone attivo diffusione semplice

6
Gli ormoni amminici sono sono sintetizzati a partire da un solo amminoacido, sono lipofili e sono immagazzinati in vescicole secretorie

7
MECCANISMO D’AZIONE 1) ORMONI PEPTIDICI
Sintesi e immagazzinamento in vescicole secretorie fino a quando la cellula non riceve un messaggio per la secrezione. Gli ormoni peptidici sono idrofili, viaggiano nel plasma in forma libera e si legano ad un recettore specifico. Il complesso ormone-recettore innesca la risposta cellulare.

8
I RECETTORI PER GLI ORMONI PEPTIDICI
Sono situati sulla membrana cellulare Determinano la sintesi di secondi messaggeri (cAMP, cGMP, DAG, Ca++) I secondi messaggeri determinano modificazioni enzimatiche dentro la cellula
 I secondi messaggeri determinano modificazioni enzimatiche dentro la cellula.")
9
2) ORMONI STEROIDEI Il colesterolo è il precursore di tutti gli ormoni steroidei. Sono lipofili e attraversano facilmente la membrana cellulare. Non sono solubili nel plasma e per questo si legano a proteine trasportatrici I RECETTORI PER GLI ORMONI STEROIDEI Si trovano nel compartimento intracellulare Il complesso ormone-recettore si lega al DNA attivando uno o più geni L’mRNA dirige la sintesi di nuove proteine

10
3) ORMONI AMMINICI I principali ormoni amminici sono: CATECOLAMINE e ORMONI TIROIDEI che derivano dall’amminoacido Tirosina MELATONINA che deriva dall’amminoacido Triptofano Le Catecolamine si comportano come gli ormoni peptidici Gli ormoni tiroidei e la melatonina si comportano come gli ormoni steroidei

11
INTERAZIONE SINERGISMO
Due ormoni hanno lo stesso effetto anche se agiscono con meccanismi diversi. La loro interazione produce un effetto maggiore della somma dei loro singoli effetti Es. Glucagone > livelli di glicemia nel sangue 10mg/ml Adrenalina > livelli di glicemia nel sangue 5mg/ml Glucagone + Adrenalina > livelli di glicemia nel sangue 22mg/ml PERMISSIVISMO Un ormone non può espletare completamente il suo effetto se non è presente un secondo ormone Es. ormoni tiroidei e fattore di crescita insulino-simile IGF-1: l’IGF-1 stimola la crescita scheletrica in presenza degli ormoni tiroidei, mentre questi, direttamente, stimolano poco la crescita ossea. ANTAGONISMO Due ormoni hanno azione fisiologica opposta Es. Glucagone > livelli di glicemia nel sangue Insulina < livelli di glicemia nel sangue

12
Regolazione della secrezione ormonale
L’attività secretoria dei tessuti endocrini è controllata con un meccanismo a feedback negativo. Esistono due circuiti di retroazione: 1. Breve 2. Lungo Nel feedback a circuito breve, la risposta del tessuto bersaglio principale viene a ricadere sulla ghiandola endocrina. Nel feedback a circuito lungo, un segnale proveniente da tessuti bersaglio secondari controlla l’attività secretrice dei tessuti endocrini.

13
IPOFISI È stata definita la ghiandola principale dell’organismo perché i suoi ormoni controllano la maggior parte delle funzioni vitali Sezione trasversa del cranio che mostra l’ipofisi. L’ipofisi è un organo impari e mediano ancorato alla faccia inferiore del cervello mediante un peduncolo di tessuto nervoso (peduncolo ipofisario o infundibulum), allocata nella sella turcica. È costituita da due parti: l’ipofisi anteriore è vero tessuto endocrino di origine epiteliale; l’ipofisi posteriore è un’estensione del tessuto nervoso cerebrale.
, allocata nella sella turcica. È costituita da due parti: l’ipofisi anteriore è vero tessuto endocrino di origine epiteliale; l’ipofisi posteriore è un’estensione del tessuto nervoso cerebrale.")
14
Sviluppo embrionale della parte anteriore e posteriore dell’ipofisi
L’ipofisi anteriore deriva da una estroflessione dell’ectoderma della cavità buccale primitiva (tasca di Rathke). L’ipofisi posteriore deriva da una estroflessione del primitivo pavimento del terzo ventricolo; essa è, pertanto, un’estensione del sistema nervoso facente parte del diencefalo. La neuroipofisi comprende: - una parte superiore più espansa del processo nervoso peduncolare, corrispondente all’eminenza mediana; - il peduncolo infundibulare, avvolto dalla parte tuberale dell’adenoipofisi; - il processo infundibulare che contiene gli ormoni
. L’ipofisi posteriore deriva da una estroflessione del primitivo pavimento del terzo ventricolo; essa è, pertanto, un’estensione del sistema nervoso facente parte del diencefalo. La neuroipofisi comprende: - una parte superiore più espansa del processo nervoso peduncolare, corrispondente all’eminenza mediana; - il peduncolo infundibulare, avvolto dalla parte tuberale dell’adenoipofisi; - il processo infundibulare che contiene gli ormoni.")
15
IPOFISI ANTERIORE O ADENOIPOFISI
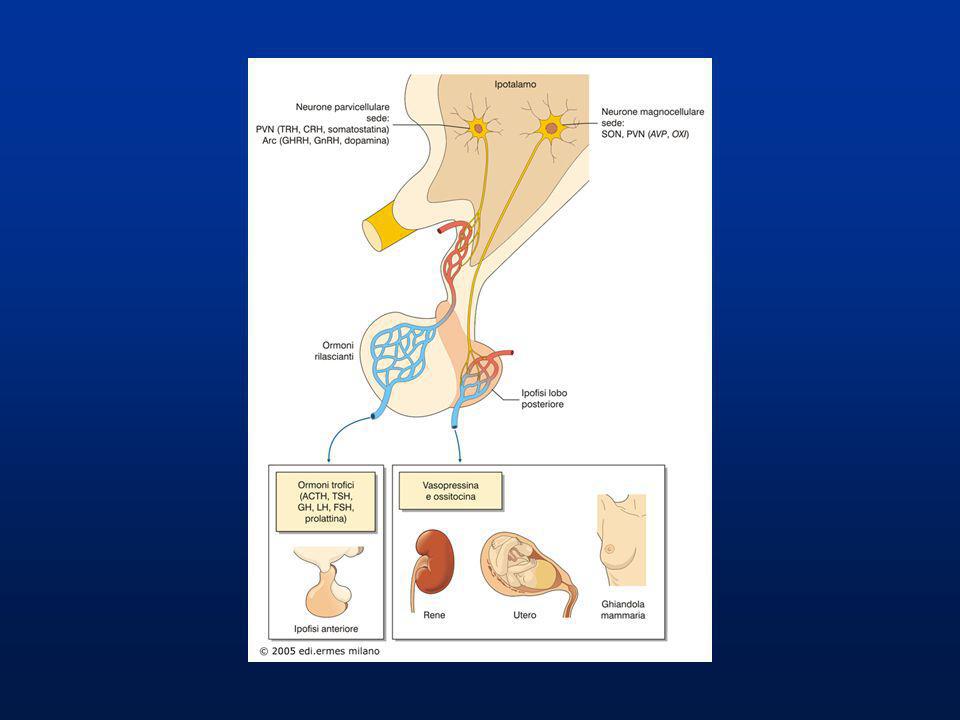
È una vera ghiandola endocrina in rapporto con l’ipotalamo mediante uno speciale gruppo di vasi sanguigni che costituisce i SISTEMA PORTALE IPOTALAMO-IPOFISARIO IPOFISI POSTERIORE O NEUROIPOFISI Sito di accumulo e rilascio di due neuroormoni sintetizzati nell’ipotalamo: OSSITOCINA ADH (Ormone AntiDiuretico) o VASOPRESSINA
 o VASOPRESSINA.")
18
Trasporto, accumulo e rilascio degli ormoni dell’ipofisi posteriore
1) Gli ormoni peptidici vengono sintetizzati nel reticolo endoplasmatico rugoso e impacchettati nell’apparato del Golgi. 2) Le vescicole secretorie contenenti l’ormone vengono trasportate per via neuronale dall’ipotalamo all’ipofisi posteriore. 3) Le vescicole vengono accumulate nella terminazione distale nell’ipofisi posteriore. 4) Gli ormoni vengono rilasciati in circolo per esocitosi. 5) Il sangue contenente gli ormoni si porta nella circolazione generale.
 Gli ormoni peptidici vengono sintetizzati nel reticolo endoplasmatico rugoso e impacchettati nell’apparato del Golgi. 2) Le vescicole secretorie contenenti l’ormone vengono trasportate per via neuronale dall’ipotalamo all’ipofisi posteriore. 3) Le vescicole vengono accumulate nella terminazione distale nell’ipofisi posteriore. 4) Gli ormoni vengono rilasciati in circolo per esocitosi. 5) Il sangue contenente gli ormoni si porta nella circolazione generale.")
19
ORMONI DELL’IPOFISI POSTERIORE
AZIONE PRINCIPALE: STIMOLA …… NATURA CHIMICA SIGLA ORMONE Il riassorbimento di acqua nei tubuli renali (azione antidiuretica); Partecipa quindi alla regolazione idro-salina ed osmotica dei liquidi extracellulari Polipeptide (9 aminoacidi) VP o ADH Vasopressina (Ormone antidiuretico) La contrazione della muscolatura liscia uterina durante il parto; Eiezione del latte dalla mammella nella poppata OXT Ossitocina
; Partecipa quindi alla regolazione idro-salina ed osmotica dei liquidi extracellulari. Polipeptide (9 aminoacidi) VP o ADH. Vasopressina (Ormone antidiuretico) La contrazione della muscolatura liscia uterina durante il parto; Eiezione del latte dalla mammella nella poppata. OXT. Ossitocina.")
20
ADH L’ADH agisce: - sulla muscolatura liscia (recettori V1) delle arteriole periferiche, provocando, in risposta a una condizione di ipovolemia, come durante una grave emorragia, vasocostrizione e aumento della pressione arteriosa sulle cellule del dotto collettore del nefrone (recettori V2), controllandone la permeabilità della membrana basale all’acqua, tramite l’inserzione dell’acquaporina-2 nella membrana apicale delle cellule principali. Se l’osmolarità plasmatica aumenta, o la pressione ematica o la volemia diminuiscono aumenta la frequenza dei potenziali d’azione negli osmocettori (centri della sete) e nelle cellule neurosecernenti, aumentando, di conseguenza, la sintesi e la liberazione nel sangue di ADH. L’insufficiente secrezione di ADH o l’insufficiente risposta dei recettori V2 all’ADH causano il diabete insipido caratterizzato da abbondante diuresi.
 delle arteriole periferiche, provocando, in risposta a una condizione di ipovolemia, come durante una grave emorragia, vasocostrizione e aumento della pressione arteriosa. sulle cellule del dotto collettore del nefrone (recettori V2), controllandone la permeabilità della membrana basale all’acqua, tramite l’inserzione dell’acquaporina-2 nella membrana apicale delle cellule principali. Se l’osmolarità plasmatica aumenta, o la pressione ematica o la volemia diminuiscono aumenta la frequenza dei potenziali d’azione negli osmocettori (centri della sete) e nelle cellule neurosecernenti, aumentando, di conseguenza, la sintesi e la liberazione nel sangue di ADH. L’insufficiente secrezione di ADH o l’insufficiente risposta dei recettori V2 all’ADH causano il diabete insipido caratterizzato da abbondante diuresi.")
21
Ossitocina L’OXT agisce sulla muscolatura liscia uterina (miometrio) e sulle cellule mioepiteliali dei dotti galattofori mammari, inducendo, con la loro contrazione, rispettivamente il travaglio del parto e l’eiezione del latte. L’azione dell’OXT sulla muscolatura liscia uterina riduce, inoltre, l’emorragia post partum. La sintesi e la liberazione dell’ormone sono indotte per via riflessa della distensione meccanica della vagina e dell’utero durante il parto e dalla suzione del capezzolo. Nella donna la secrezione di ossitocina è stimolata anche da stimoli psicologici, come il pianto del lattante. Lo stress emotivo, la paura inibiscono la secrezione di OXT. L’azione dell’ormone nel maschio non è nota.
 e sulle cellule mioepiteliali dei dotti galattofori mammari, inducendo, con la loro contrazione, rispettivamente il travaglio del parto e l’eiezione del latte. L’azione dell’OXT sulla muscolatura liscia uterina riduce, inoltre, l’emorragia post partum. La sintesi e la liberazione dell’ormone sono indotte per via riflessa della distensione meccanica della vagina e dell’utero durante il parto e dalla suzione del capezzolo. Nella donna la secrezione di ossitocina è stimolata anche da stimoli psicologici, come il pianto del lattante. Lo stress emotivo, la paura inibiscono la secrezione di OXT. L’azione dell’ormone nel maschio non è nota.")
22
ADENOIPOFISI

23
IPOTALAMO ADENOIPOFISI
CRH o Corticoliberina + ACTH o Corticotropina Glucocorticoidi (N ParaVentricolare) Deriva dalla pro-opiomelanocortina. Secrezione pulsatile 8 o 9 episodi intensificata dagli stress TRH o Tireoliberina + TSH o Tireotropina T3 e T4 SS o Somatostatina - TSH DA o Dopamina - TSH (N Arcuato) Gn-RH o Gonadoliberina + Gonadotropine (FSH o LH) Testicoli e Ovaie GHRH o Somatoliberina + GH o STH o Somatotropina Fegato (IGF-1 o (N-Aarcuato) Somatomedina C) GHIH o SS o Somatostatina - GH PRL-RH o PRH + PRL o Prolattina Mammelle e Ovaie PRL-IH PIH - PRL o Prolattina
 Deriva dalla pro-opiomelanocortina. Secrezione pulsatile 8 o 9 episodi. intensificata dagli stress. TRH o Tireoliberina + TSH o Tireotropina T3 e T4. SS o Somatostatina - TSH. DA o Dopamina - TSH. (N Arcuato) Gn-RH o Gonadoliberina + Gonadotropine (FSH o LH) Testicoli e Ovaie. GHRH o Somatoliberina + GH o STH o Somatotropina Fegato (IGF-1 o (N-Aarcuato) Somatomedina C) GHIH o SS o Somatostatina - GH. PRL-RH o PRH + PRL o Prolattina Mammelle e Ovaie. PRL-IH PIH - PRL o Prolattina.")
24
ORMONI DELL’IPOFISI ANTERIORE
Lo sviluppo dei follicoli e la secrezione degli ormoni estrogeni nell’ovaio; la spermatogenesi nel testicolo Glicoproteina FSH Follicolo-stimolante AZIONE PRINCIPALE: STIMOLA …… NATURA CHIMICA SIGLA ORMONE La crescita corporea e la sintesi proteica nelle cellule Proteina GH Somatotropo L’ovulazione e la formazione del corpo luteo; la secrezione di androgeni dalle cellule di Leydig del testicolo LH Luteinizzante La secrezione degli ormoni tiroidei dalla tiroide TSH Tireotropo La secrezione di corticosteroidi dalla corticale del surrene Polipeptide ACTH Adreno-corticotropo Lo sviluppo e la secrezione lattea della ghiandola mammaria PRL Prolattina

27
GH La secrezione di GH o somatotropina è regolata dall’azione contrapposta di 2 ormoni ipotalamici: il GHRH o somatoliberina che ne stimola il rilascio, e il GHIH o somatostatina che ne impedisce il rilascio. La secrezione di GH è influenzata da diversi fattori. È stimolata dal sistema simpatico, dallo stress, dall’attività fisica, dall’ipoglicemia. IL GH viene secreto soprattutto durante l‘infanzia e la gioventù. Dopo i 20 anni, la sua sintesi diminuisce velocemente al punto che di solito la concentrazione di tale ormone in una persona di 50 anni è circa la metà di una di 20. È utilizzato, in modo dannoso, dagli atleti che vogliono aumentare la propria massa muscolare. Tale pratica è molto pericolosa in quanto la somministrazione esogena determina atrofia delle cellule dell'adenoipofisi preposte alla sua secrezione. Tessuti bersaglio: sono quelli principalmente implicati nell'accrescimento: tessuto osseo, tessuto muscolare scheletrico e tessuto adiposo

28
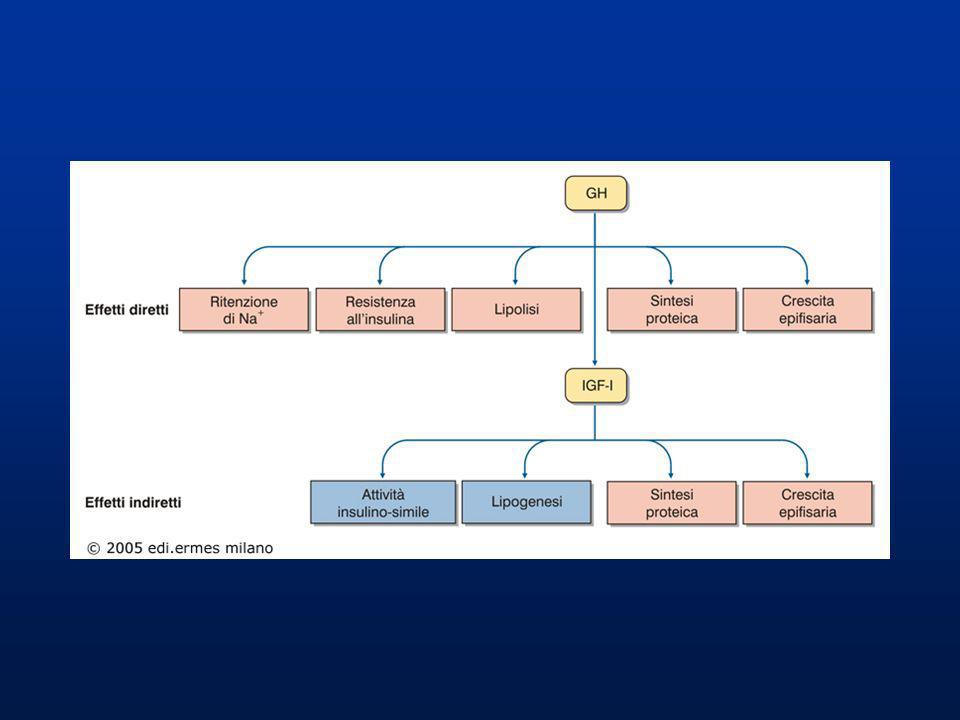
GH Il GH stimola la sintesi epatica del fattore di crescita insulino-simile (più conosciuto come IGF-I o come somatomedina C). L’IGF-I viaggia nel plasma legato a proteine specifiche (IGFBP), soprattutto di origine epatica, che ne regolano la biodisponibilità. Il GH riduce l’assunzione di glucosio del muscolo tendendo ad aumentare la glicemia. Il GH e l’IGF-I stimolano: 1) l’ingresso di amminoacidi e la sintesi proteica nelle cellule epatiche, muscolari scheletriche e, particolarmente, delle zone di accrescimento osseo-cartilagineo, regolando l’accrescimento in altezza; 2) la lipolisi e il catabolismo lipidico per utilizzare gli acidi grassi liberi a scopo energetico e risparmiare le proteine; 3) la gluconeogenesi nel fegato e la glicogenosintesi nei tessuti.
, soprattutto di origine epatica, che ne regolano la biodisponibilità. Il GH riduce l’assunzione di glucosio del muscolo tendendo ad aumentare la glicemia. Il GH e l’IGF-I stimolano: 1) l’ingresso di amminoacidi e la sintesi proteica nelle cellule epatiche, muscolari scheletriche e, particolarmente, delle zone di accrescimento osseo-cartilagineo, regolando l’accrescimento in altezza; 2) la lipolisi e il catabolismo lipidico per utilizzare gli acidi grassi liberi a scopo energetico e risparmiare le proteine; 3) la gluconeogenesi nel fegato e la glicogenosintesi nei tessuti.")
29
CARTILAGINE DI ACCRESCIMENTO
Condrogenesi Ossificazione Altezza scheletro Dimens. organi OSTEOBLASTI Attivazione Accresc. corporeo Somatomedine ALTRE CELLULE numero dimensione GH FEGATO MUSCOLO Sintesi proteica Ossidazione ac. gr. Glicolisi Aminoacidi plasm. Glicemia TESS: ADIPOSO Mobilizz. lipidi Adiposità corporea NEFA plasm. Effetti metabolici

31
Patologie causate da un’alterazione della secrezione di GH
Nanismo ipofisario: sviluppo ridotto del corpo, causato da un’insufficiente secrezione di ormone della crescita durante l’età infantile e l’adolescenza Gigantismo: dimensioni e statura eccessive causate da una ipersecrezione di ormone della crescita durante l’età infantile (prima della pubertà) Acromegalia: ingrossamento delle ossa del capo e delle estremità causato da una ipersecrezione di ormone della crescita durante l’età adulta
 Acromegalia: ingrossamento delle ossa del capo e delle estremità causato da una ipersecrezione di ormone della crescita durante l’età adulta.")
32
Prolattina La secrezione della prolattina è indotto dall’ormone ipotalamico specifico per il suo rilascio il PRH. I recettori della PRL sono presenti nelle mammelle e nelle gonadi femminili. Non è chiaro il significato della prolattina nel maschio. L’aumento degli estrogeni durante la gravidanza stimola l’accrescimento e la proliferazione delle cellule lattotrope ipofisarie a tale punto che l’ipofisi arriva a raddoppiare le sue dimensioni durante la normale gravidanza. FUNZIONE: insieme agli estrogeni, al progesterone, al GH e al cortisolo, stimola la proliferazione e la ramificazione dei dotti mammari. Dopo il parto stimola la sintesi del latte Picco di produzione: dopo il parto Controllo +: Fattore di rilascio ipotalamico PRH Controllo -: dopamina, stress, ipoglicemia

33
TIROIDE La tiroide è un organo impari e mediano, costituita da due lobi e un istmo che li unisce, ed è situata anteriormente alla porzione superiore della trachea. La tiroide è avvolta da una capsula fibrosa con dei sepimenti interni che la suddividono in lobuli. Ciascun lobulo è costituito da numerose vescicole chiuse dette follicoli tiroidei. Il follicolo è l’unità anatomo-funzionale della tiroide ed è una vescicola irregolarmente sferoidale, chiusa da un epitelio monostratificato di cellule secernenti dette “follicolari” o tireociti, che poggiano esternamente alla membrana basale e delimitano internamente una cavità o lume follicolare contenente una sostanza vischiosa detta colloide. colloide La colloide è una soluzione di una glicoproteina, la tireoglobulina (Tg) elaborata dalle cellule follicolari, che costituisce la forma di deposito degli ormoni tiroidei. Tra i follicoli, a ridosso della membrana basale, sono presenti rare cellule parafollicolari o cellule C secernenti calcitonina.
 elaborata dalle cellule follicolari, che costituisce la forma di deposito degli ormoni tiroidei. Tra i follicoli, a ridosso della membrana basale, sono presenti rare cellule parafollicolari o cellule C secernenti calcitonina.")
34
Gli ormoni tiroidei vengono secreti dalla tiroide in risposta all'ormone ipofisario TSH, la cui produzione è regolata dall'ormone ipotalamico TRH. Gli ormoni tiroidei: sono due e vengono chiamati rispettivamente triiodotironina (T3) e tiroxina (T4) Sono critici per lo sviluppo cerebrale e somatico del bambino e dell’attività metabolica dell’adulto Influenzano la funzione di ogni organo e tessuto Devono sempre essere disponibili Vi sono grandi depositi disponibili (colloide dei follicoli tiroidei e proteine plasmatiche di trasporto) di ormoni tiroidei, la cui sintesi e secrezione sono strettamente regolate da meccanismi molto sensibili.
 e tiroxina (T4) Sono critici per lo sviluppo cerebrale e somatico del bambino e dell’attività metabolica dell’adulto. Influenzano la funzione di ogni organo e tessuto. Devono sempre essere disponibili. Vi sono grandi depositi disponibili (colloide dei follicoli tiroidei e proteine plasmatiche di trasporto) di ormoni tiroidei, la cui sintesi e secrezione sono strettamente regolate da meccanismi molto sensibili.")
35
Schema della sintesi degli ormoni tiroidei
La sintesi degli ormoni tirodei è possibile in presenza di una sufficiente quantità di iodio, che deve essere introdotto con la dieta. A livello gastro-intestinale si forma ioduro, che viene assorbito e passa in circolo e, 1) a livello tiroideo, viene captato dalle cellule follicolari per dare origine allo iodio molecolare per azione della perossidasi tiroidea (tireoperossidasi TPO). 2) La tireoglobulina, lunga catena proteica presente nella colloide, lega lo iodio a livello dei residui di tirosina, presenti nella sua struttura. 1 2 3 4 La secrezione degli ormoni tirodei avviene quando 3) il tireocita ingloba, per pinocitosi, la colloide. Sulla vescicola convergono i lisosomi attivati e la tireoglobulina viene scissa per dare origine 4) agli ormoni T3 e T4, liberati nel sangue capillare, MIT, DIT e aminoacidi.
 a livello tiroideo, viene captato dalle cellule follicolari per dare origine allo iodio molecolare per azione della perossidasi tiroidea (tireoperossidasi TPO). 2) La tireoglobulina, lunga catena proteica presente nella colloide, lega lo iodio a livello dei residui di tirosina, presenti nella sua struttura La secrezione degli ormoni tirodei avviene quando 3) il tireocita ingloba, per pinocitosi, la colloide. Sulla vescicola convergono i lisosomi attivati e la tireoglobulina viene scissa per dare origine 4) agli ormoni T3 e T4, liberati nel sangue capillare, MIT, DIT e aminoacidi.")
36
Il TSH stimola la sintesi di ormoni tiroidei promuovendo la sintesi di tireoglobulina (Tg) da parte dei tireociti. La Tg è una proteina ad alto contenuto di residui tirosinici, prodotta nell’apparato di Golgi e riversata sul versante apicale del tireocita. Il TSH stimola l’espressione del NIS (sodium/iodide symporter): pompa che trasporta lo IODIO all’interno del tireocita contro gradiente. Sfrutta il gradiente del sodio, che viene espulso dalla pompa Na/K ATPasi. Lo IODIO viene quindi ORGANICATO nei residui tirosinici della Tg ad opera della tireoperossidasi (TPO) sulla membrana apicale del tireocita. TPO catalizza la formazione di T4 da due molecole di DIT (dioidiotirosina) e la formazione di T3 da una molecola di DIT ed una di MIT (monoiodiotirosina).
: pompa che trasporta lo IODIO all’interno del tireocita contro gradiente. Sfrutta il gradiente del sodio, che viene espulso dalla pompa Na/K ATPasi. Lo IODIO viene quindi ORGANICATO nei residui tirosinici della Tg ad opera della tireoperossidasi (TPO) sulla membrana apicale del tireocita. TPO catalizza la formazione di T4 da due molecole di DIT (dioidiotirosina) e la formazione di T3 da una molecola di DIT ed una di MIT (monoiodiotirosina).")
37
Struttura degli ormoni tiroidei e dei composti correlati
Nel processo di sintesi degli ormoni tiroidei si possono formare anche molecole diverse da T3 e T4, che vengono però metabolizzate all’interno del tireocita, recuperando lo IODIO ed il residuo tirosinico.

38
Gli ormoni tiroidei in circolo sono rappresentati soprattutto dal T4
Gli ormoni tiroidei in circolo sono rappresentati soprattutto dal T4. Il T3 circolante deriva per l’80% dalla deiodazione di T4 in periferia. T4 e T3 circolano legati a proteine plasmatiche: TBG = thyroxine-binding globulin, ogni molecola lega una molecola di T4 o T3. Lega il 70% della T4 circolante e l’80% della T3 circolante. TTR = transthyretin. Lega l’ 11% della T4 circolante e il 9% della T3 circolante. Albumina = Lega il 20% della T4 circolante e l’ 11% della T3 circolante. La forma attiva a livello cellulare è rappresentata dal T3, che deriva per metabolizzazione del T4, cui viene tolto un atomo di iodio sull’anello esterno da parte delle DEIODINASI. Gran parte del T3 si forma all’interno delle cellule bersaglio. Il T3 ha affinità per i recettori nucleari specifici per gli ormoni tiroidei 15 volte superiore rispetto a T4.

39
AZIONI DEGLI ORMONI TIROIDEI
Sono necessari per lo sviluppo del sistema nervoso centrale nel feto e nelle fasi postnatali Importanti effetti sui processi di differenziazione cerebrale nelle prime settimane di vita. Sono necessari per lo sviluppo dello scheletro fetale Sono indispensabili per il normale accrescimento corporeo nel bambino, e la maturazione dei vari apparati, soprattutto quello scheletrico. Azione termogenetica Effetti sul metabolismo glucidico Lipolisi e lipogenesi Sintesi proteica Effetti sul sistema nervoso centrale Effetti sul sistema cardiovascolare

40
Effetti sul metabolismo glucidico
Gli ormoni tiroidei: inducono la produzione epatica di glucosio, aumentano la glicogenolisi e la gluconeogenesi; promuovono l’utilizzazione del glucosio aumentando l'attività di enzimi coinvolti nell’ossidazione del glucosio Lipolisi e lipogenesi stimolano l’attività della lipasi ormono-sensibile → lipolisi stimolano la sintesi e l’ossidazione del colesterolo e la sua conversione in acidi bilari lipogenesi: favorita la sintesi di acidi grassi effetto prevalente su lipolisi = aumenta la disponibilità di ac. grassi, che possono essere ossidati e formare ATP, utilizzato per la termogenesi Sintesi proteica Aumento delle sintesi proteiche (proteine strutturali, enzimi, ormoni); effetto trofico sul muscolo Stimolano l’ossificazione endocondrale, la crescita lineare e la maturazione dei centri epifisari. Gli effetti sulla crescita lineare sono in buona parte mediati dalla loro azione sulla secrezione di GH e di IGF-1 Hanno azione sulla matrice proteica e sulla mineralizzazione dell’osso.
; effetto trofico sul muscolo. Stimolano l’ossificazione endocondrale, la crescita lineare e la maturazione dei centri epifisari. Gli effetti sulla crescita lineare sono in buona parte mediati dalla loro azione sulla secrezione di GH e di IGF-1. Hanno azione sulla matrice proteica e sulla mineralizzazione dell’osso.")
41
Azione termogenetica Gli ormoni tiroidei contribuiscono in modo fondamentale alla spesa energetica ed alla produzione di calore, regolando direttamente il metabolismo basale. Tale azione dipende da: aumento dell’attività metabolica di tutti i tessuti (aumento del consumo di O2, della produzione di calore e della velocità di utilizzazione delle sostanze energetiche) Normale consumo di O2 = 250 ml/min Ipotiroidismo » 150 ml/min. Ipertiroidismo » 400 ml/min
 Normale consumo di O2 = 250 ml/min. Ipotiroidismo » 150 ml/min. Ipertiroidismo » 400 ml/min.")
42
Effetti sul sistema nervoso centrale
Gli ormoni tiroidei regolano lo sviluppo e la differenziazione del sistema nervoso centrale durante la vita fetale e nelle prime settimane di vita, quando assicurano una corretta mielinizzazione delle strutture nervose Deficit della funzionalità tiroidea in epoca precoce comportano gravi ripercussioni sul SNC e possono compromettere il quoziente intellettivo del soggetto. Effetti sul sistema cardiovascolare aumento del numero dei recettori β-adrenergici aumenta la contrattilità cardiaca aumenta frequenza cardiaca aumenta l’eccitabilità della miocellula aumenta il consumo tissutale di O2

43
CALCITONINA Le cellule C sono stimolate a sintetizzare e secernere l’ormone Calcitonina (CAL) in risposta all’aumento della concentrazione plasmatica di Ca2+ (ipercalcemia). La CAL non è immagazzinata in depositi cellulari. La CAL è un peptide che agisce sull’osso, dove inibisce l’attività degli osteoclasti, i soli in cui sono stati trovati i recettori, ma stimolerebbe anche quella degli osteoblasti, determinando la deposizione nell’osso di calcio e fosforo e la riduzione della concentrazione plasmatica del Ca2+. Il ruolo fisiologico della CAL non è ancora del tutto chiaro. A differenza del paratormone, non si verificano nell’uomo modificazioni del metabolismo del calcio e del fosforo quando la disponibilità dell’ormone subisce variazioni, anche grandi, e non risultani in bibliografia effetti sicuri attribuibili alla deficienza di CAL (es. pazienti tiroidectomizzati). Si pensa, tuttavia, che la Cal prevenga l’ipercalcemia dopo un pasto che comporti l’immissione di calcio in circolo. La sintesi di CAL diminuisce con l’età. Questa diminuzione potrebbe contribuire in parte alla progressiva decalcificazione ossea nella senescenza.
 in risposta all’aumento della concentrazione plasmatica di Ca2+ (ipercalcemia). La CAL non è immagazzinata in depositi cellulari. La CAL è un peptide che agisce sull’osso, dove inibisce l’attività degli osteoclasti, i soli in cui sono stati trovati i recettori, ma stimolerebbe anche quella degli osteoblasti, determinando la deposizione nell’osso di calcio e fosforo e la riduzione della concentrazione plasmatica del Ca2+. Il ruolo fisiologico della CAL non è ancora del tutto chiaro. A differenza del paratormone, non si verificano nell’uomo modificazioni del metabolismo del calcio e del fosforo quando la disponibilità dell’ormone subisce variazioni, anche grandi, e non risultani in bibliografia effetti sicuri attribuibili alla deficienza di CAL (es. pazienti tiroidectomizzati). Si pensa, tuttavia, che la Cal prevenga l’ipercalcemia dopo un pasto che comporti l’immissione di calcio in circolo. La sintesi di CAL diminuisce con l’età. Questa diminuzione potrebbe contribuire in parte alla progressiva decalcificazione ossea nella senescenza.")
44
PARATIROIDI Le paratiroidi sono piccole masse endocrine addossate alla superficie dorsale della tiroide. Le paratiroidi e l’ormone da loro prodotto sono essenziali per la vita. Generalmente le paratiroidi sono quattro, una coppia per ciascun lobo della tiroide, di cui una superiore e una inferiore. Esse secernono il paratormone (PTH) ossia l’ormone che regola il metabolismo del calcio e del fosforo, controllando, di concerto con la forma attiva della vitamina D (1,25-diidrossivitamina D3), l’equilibrio fosfo-calcico. La funzione del PTH è di elevare la concentrazione plasmatica di calcio e di diminuire quella del fosforo. Nell’adulto sano i valori normali della calcemia totale sono intorno a 10 mg/dl (intervallo di 9 – 10,5). La secrezione del PTH da parte delle paratiroidi è regolata direttamente dalla concentrazione plasmatica di calcio: l’aumento della calcemia provoca una riduzione della secrezione e deposizione di calcio nelle ossa; la diminuzione della calcemia, al contrario, determina un incremento della sua secrezione e, di conseguenza, del riassorbimento di calcio dalle ossa. Il PTH esplica la sua azione principalmente su tre organi: osso, rene e intestino.
 ossia l’ormone che regola il metabolismo del calcio e del fosforo, controllando, di concerto con la forma attiva della vitamina D (1,25-diidrossivitamina D3), l’equilibrio fosfo-calcico. La funzione del PTH è di elevare la concentrazione plasmatica di calcio e di diminuire quella del fosforo. Nell’adulto sano i valori normali della calcemia totale sono intorno a 10 mg/dl (intervallo di 9 – 10,5). La secrezione del PTH da parte delle paratiroidi è regolata direttamente dalla concentrazione plasmatica di calcio: l’aumento della calcemia provoca una riduzione della secrezione e deposizione di calcio nelle ossa; la diminuzione della calcemia, al contrario, determina un incremento della sua secrezione e, di conseguenza, del riassorbimento di calcio dalle ossa. Il PTH esplica la sua azione principalmente su tre organi: osso, rene e intestino.")
45
Nell’osso stimola il rilascio di calcio e di fosforo, inibendo gli osteoblasti e attivando, indirettamente, gli osteoclasti attraverso il rilascio di IL-1 da parte degli osteoblasti, i soli ad avere il recettore per l’ormone e per la vitamina D3. Nel tubulo renale (tubulo contorto distale e porzione ascendente spessa dell’ansa di Henle) stimola il riassorbimento di calcio e inibisce (tubulo prossimale) quello del fosforo (HPO42-, azione fosfaturica) e del bicarbonato (HCO3-). L’azione del PTH ha sull’intestino è indiretta: l’ormone stimola direttamente il tubulo prossimale renale a produrre la forma attiva della vitamina D3 (colecalciferolo), attivando l’enzima renale 1 α-idrossilasi. L’aumentata sintesi di 1,25-diidrossicolecalciferolo o calcitriolo o ormone D a sua volta si traduce in un incremento dell’assorbimento intestinale di calcio e del riassorbimento osseo; ne deriva un ulteriore incremento del carico di calcio filtrato dal rene ed una minor ritenzione renale del fosforo. In definitiva le azioni del PTH si traducono in un aumento della concentrazione sierica del calcio e una diminuzione di quella del fosforo.

46
Paratiroidi e malattie correlate
Iperparatiroidismo E’ una condizione caratterizzata da un’aumentata secrezione di paratormone da parte di una o di tutte le paratiroidi. Ipoparatiroidismo Una ridotta funzione del tessuto paratiroideo determina un quadro di deficit ormonale, noto come ipoparatiroidismo. La forma più frequente è di certo l'ipoparatiroidismo secondario a chirurgia del collo (tiroidectomia e paratiroidectomia), che può comportare l’ablazione o la devascolarizzazione delle ghiandole paratiroidee determinando carenza di calcio ematico (Ca2+), causa di ipereccitabilità, insorgenza spontanea di potenziali d’azione nei motoneuroni e conseguente tetania muscolare. La carenza del minerale, senza interventi, porta in breve tempo a morte. Di grande interesse clinico è anche il meno comune ipoparatiroidismo primitivo. Malattie ossee da alterazione del metabolismo del cacio. - Osteoporosi - Osteomalacia - Rachitismo
, che può comportare l’ablazione o la devascolarizzazione delle ghiandole paratiroidee determinando carenza di calcio ematico (Ca2+), causa di ipereccitabilità, insorgenza spontanea di potenziali d’azione nei motoneuroni e conseguente tetania muscolare. La carenza del minerale, senza interventi, porta in breve tempo a morte. Di grande interesse clinico è anche il meno comune ipoparatiroidismo primitivo. Malattie ossee da alterazione del metabolismo del cacio. - Osteoporosi. - Osteomalacia. - Rachitismo.")
48
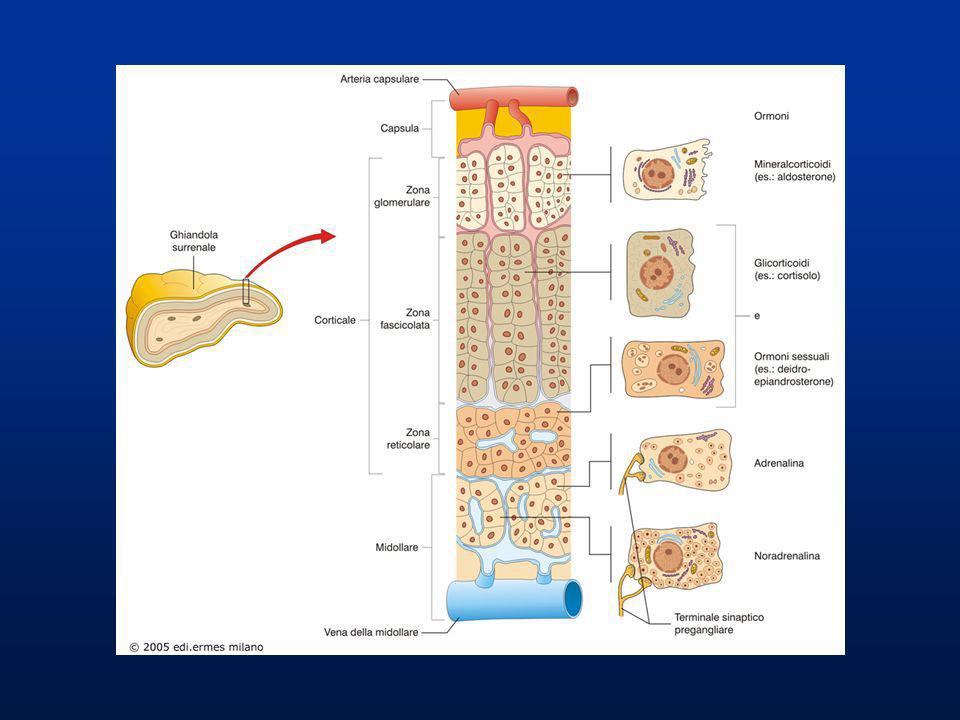
GHIANDOLE SURRENALI Le ghiandole surrenali sono organi pari di forma piramidale che poggiano sul polo posteriore dei reni. Il parenchima surrenale è costituito da una parte periferica più compatta detta CORTICALE (80%) e una parte centrale detta MIDOLLARE (20%). La parte corticale dei surreni è caratterizzate da tre zone: - zona glomerulare (ZG) (15%) periferica - zona fascicolata (ZF) (78%) intermedia - zona reticolare (ZR) (7%) più interna la corticale del surrene o corticosurrene secerne gli ormoni corticosteroidi o corticoidi: nella ZG i mineralcorticoidi (aldosterone) nella ZF i glucocorticodi (cortisolo, corticosterone) nella ZR gli androgeni deidroepiandrosterone (DHEA) e il duo estere deidroepiandrosterone solfato (DHEAS)
 e una parte centrale detta MIDOLLARE (20%). La parte corticale dei surreni è caratterizzate da tre zone: - zona glomerulare (ZG) (15%) periferica. - zona fascicolata (ZF) (78%) intermedia. - zona reticolare (ZR) (7%) più interna. la corticale del surrene o corticosurrene secerne gli ormoni corticosteroidi o corticoidi: nella ZG i mineralcorticoidi (aldosterone) nella ZF i glucocorticodi (cortisolo, corticosterone) nella ZR gli androgeni deidroepiandrosterone (DHEA) e il duo estere deidroepiandrosterone solfato (DHEAS)")
50
Il colesterolo, proveniente dalle LDL o sintetizzato ex novo dall’acetato, è il precursore degli ormoni corticoidi. Le cellule corticali contengono depositi intracellulari di colesterolo, prevalentemente sottoforma di estere, che costituiscono la forma più immediata di colesterolo per la steroidogenesi.

51
I MINERALCORICOIDI Il principale mineralcorticoide prodotto dalla ZG è l’ALDOSTERONE. La secrezione dell’aldosterone è stimolata poco dall’ACTH, ma è stimolata soprattutto dal sistema renina-angiotensina II e dal tasso ematico dello ione K+.

52
L’aldosterone nel rene promuove la ritenzione degli ioni Na+ e la secrezione degli ioni K+ e H+. I suoi siti d’azione sono la porzione terminale del tubulo contorto distale e il dotto collettore. L’effetto sodioritentivo dell’aldosterone è presente anche in altre sedi, come nelle ghiandole salivari, sudoripare e nella mucosa gastrointestinale. L’effetto netto dell’aldosterone è di aumentare il volume del liquido extracellulare e del sangue e di conseguenza la pressione sanguigna e la gittata cardiaca. Il peptide natriuretico atriale, prodotto dal cuore, inibisce il rilascio di aldosterone determinando riduzione del volume ematico e, di conseguenza, della pressione arteriosa.

53
I GLUCOCORTICOIDI Il principale glucocorticoide prodotto dalla ZF è il CORTISOLO. La secrezione del cortisolo è stimolata dall’ACTH, indotto dall’ormone ipotalamico specifico per il suo rilascio il CRH. I glucocorticoidi stimolano la secrezione acida gastrica, aumentano il riassorbimento osseo e l’escrezione urinaria di calcio, la responsività degli adrenorecettori alle catacolamine, la velocità di filtrazione glomerulare, la percezione e l’integrazione di stimoli sensoriali (acustici, tattili, olfattivi e gustativi). Inibiscono i processi immunologici e infiammatori Le azioni principali riguardano la regolazione del metabolismo glucidico (da cui deriva il loro nome) e l’adattamento dell’organismo agli stati di stress di lunga durata, tra i quali anche il digiuno prolungato. Il cortisolo favorisce in diversi tessuti il catabolismo proteico e a livello del fegato e, in minior misura, del rene, la gluconeogenesi Ha effetto diabetogeno, in quanto antagonista dell’insulina, e lipolitico
. Inibiscono i processi immunologici e infiammatori. Le azioni principali riguardano la regolazione del metabolismo glucidico (da cui deriva il loro nome) e l’adattamento dell’organismo agli stati di stress di lunga durata, tra i quali anche il digiuno prolungato. Il cortisolo favorisce in diversi tessuti il catabolismo proteico e a livello del fegato e, in minior misura, del rene, la gluconeogenesi. Ha effetto diabetogeno, in quanto antagonista dell’insulina, e lipolitico.")
54
ANDROGENI Il DHEA e il DHEAS sono androgeni deboli. Sono destinati ad essere trasformati perifericamente nel più potente androgeno che è il TESTOSTERONE. Gli androgeni surrenalici hanno, nel maschio, un significato biologico marginale, perché il testosterone ottenuto da essi è secondario a quello prodotto ex novo nelle gonadi maschili, mentre, nella femmina, sono i soli presenti. Nella donna, gli androgeni sono una fonte di estrogeni in cui sono convertiti nel tessuto adiposo dalle aromatasi. Un eccesso di androgeni surrenalici può causare virilizzazione della donna.

55
ORMONI DELLA MIDOLLARE DEL SURRENE.
I neurormoni della midollare del surrene sono le catecolamine adrenalina o epinifrina (80%) e noradrenalina o norepinefrina (20%), sintetizzate a partire dall’aminoacido tirosina. La sintesi è regolata dall’innervazione simpatica, dal cortisolo, dall’ACTH. L’adrenalina e, in minore quantità, la noradrenalina sono veicolate dal sangue e si legano ai recettori adrenergici presenti sulla membrana plasmatica delle cellule bersaglio. L’adrenalina interagisce con i recettori a- e b-adrenergici, mentre la noradrenalina interagisce soprattutto con i recettori a-adrenergici. Dal punto di vista metabolico l’adrenalina stimola nelle cellule muscolari la glicogenolisi e nelle cellule epatiche, oltre alla glicogenolisi, anche la gluconeogenesi con effetto iperglicemizzante. Negli adipociti stimola la lipolisi, liberando acidi grassi e glicerolo nel sangue. L’adrenalina rinforza i suoi effetti sul metabolismo agendo anche sul pancreas endocrino, stimolando al secrezione di glucagone e inibendo quella dell’insulina.
 e noradrenalina o norepinefrina (20%), sintetizzate a partire dall’aminoacido tirosina. La sintesi è regolata dall’innervazione simpatica, dal cortisolo, dall’ACTH. L’adrenalina e, in minore quantità, la noradrenalina sono veicolate dal sangue e si legano ai recettori adrenergici presenti sulla membrana plasmatica delle cellule bersaglio. L’adrenalina interagisce con i recettori a- e b-adrenergici, mentre la noradrenalina interagisce soprattutto con i recettori a-adrenergici. Dal punto di vista metabolico l’adrenalina stimola nelle cellule muscolari la glicogenolisi e nelle cellule epatiche, oltre alla glicogenolisi, anche la gluconeogenesi con effetto iperglicemizzante. Negli adipociti stimola la lipolisi, liberando acidi grassi e glicerolo nel sangue. L’adrenalina rinforza i suoi effetti sul metabolismo agendo anche sul pancreas endocrino, stimolando al secrezione di glucagone e inibendo quella dell’insulina.")
56
GONADI Le gonadi dell’adulto sono organi pari destinati ad una duplice funzione: esocrina, consistente nella produzione di gameti e fluidi e molecole nutritive facilitanti la fusione dei gameti di sesso opposto endocrina consistente nella produzione degli ormoni sessuali, rispettivamente, per i maschi, il testosterone e il diidrotestosterone, per le femmine l’estradiolo, l’estrone, l’estriolo e il progesterone. L’ipotalamo induce la liberazione delle gonadotropine ipofisarie tramite il GnRH. FSH e LH agiscono direttamente sulle gonadi, dove avviano e promuovono la gametogenesi e controllano la sintesi e il rilascio degli ormoni sessuali steroidei. Gli ormoni steroidei gonadici inducono lo sviluppo e il normale funzionamento degli organi riproduttivi ed esercitano numerosi effetti in sedi e organi bersaglio diversi. Nella regolazione delle gonadi sono direttamente coinvolti anche altri ormoni come il GH, la PRL e il TSH.

57
Il testosterone inibisce il rilascio di LH in entrambi i sessi, nel maschio anche di FSH.
Attivina (fattore gonadico peptidico) stimola il rilascio delle gonadotropine (particolarmente dell’FSH) Inibina e follistatina (fattori gonadici peptidici) inibiscono il rilascio delle gonadotropine (particolarmente dell’FSH) La secrezione delle gonadotropine presenta nel maschio dei picchi con intervalli di minuti, mentre nella femmina varia secondo la fase del ciclo mestruale. Gli androgeni sono precursori obbligati degli estrogeni in cui sono trasformati ad opera delle aromatasi. Gli ormoni sessuali circolano nel sangue legati a proteine plasmatiche come l’albumina e l’SHBG.
 stimola il rilascio delle gonadotropine (particolarmente dell’FSH) Inibina e follistatina (fattori gonadici peptidici) inibiscono il rilascio delle gonadotropine (particolarmente dell’FSH) La secrezione delle gonadotropine presenta nel maschio dei picchi con intervalli di minuti, mentre nella femmina varia secondo la fase del ciclo mestruale. Gli androgeni sono precursori obbligati degli estrogeni in cui sono trasformati ad opera delle aromatasi. Gli ormoni sessuali circolano nel sangue legati a proteine plasmatiche come l’albumina e l’SHBG.")
58
Secrezione degli ormoni testicolari
Le cellule interstiziali di Leydig sintetizzano e secernono il testosterone e il diidrotestosterone. Le cellule di Sertoli non sono capaci di sintetizzare ormoni steroidei, ma convertono il testosterone, prodotto dalle cellule di Leydig, in estradiolo e diidrotestosterone, e secernono l’inibina. Le cellule di Leydig hanno recettori di membrana specifici per l’LH, ma sono sensibili anche all’FSH. L’FSH induce nelle cellule di Leydig l’aumento dei recettori membranari per l’LH, amplificandone conseguentemente gli effetti. Le cellule di Sertoli hanno recettori di membrana specifici per l’FSH. L’inibina che esse secernono ha un ruolo importante nel controllo a feedback negativo dell’FSH ipofisario. La secrezione di FSH è inibita anche anche dal testosterone e dall’estradiolo. L’FSH e il testosterone iniziano e mantengono al spermatogenesi nei tubuli seminiferi: il testosterone controllerebbe la fase iniziale, l’FSH la fase terminale. Il testosterone esercita azioni di tipo anabolico e androgenico su diversi tessuti e apparati come muscolare scheletrico, adiposo, epatico, pilifero, sebaceo.

59
Secrezione degli ormoni ovarici
La maturazione dei follicoli e la normale secrezione degli ormoni ovarici sono regolate dalle gonodotropine adenoipofisarie FSH ed LH, e dal fattore ipotalamico responsabile del loro rilascio GnRH. FSH ed LH stimolano la sintesi e la secrezione degli ormoni steroidei estrogeni, progesterone e androgeni, e degli ormoni peptidici inibina e ralaxina. Le azioni principali degli estrogeni e del progesterone sono: l’induzione e il mantenimento dei caratteri sessuali secondari femminili; la capacità procreativa ciclica della donna il mantenimento della gravidanza La preparazione delle ghiandole mammarie per l’allattamento

60
Correlazioni endocrine durante il ciclo ovarico e dell’endometrio

61
PANCREAS ENDOCRINO Il pancreas è una ghiandola con funzione sia esocrina (97-99%) che endocrina (1-3%). Le “isole pancreatiche”, sparse nel tessuto esocrino, secernono nel sangue ormoni regolatori del metabolismo e sono costituite da quattro tipi di cellule, ciascuno secernente un ormone distinto: le cellule a (10-20%) che secernono glucagone, le cellule b (75-80%) che secernono insulina, le cellule d (3-4%) e le cellule j (1-2%) che secernono rispettivamente la somatostatina (SS) e il polipeptide pancreatico (PP). Gli ormoni pancreatici sono riversati nelle vene pancreatiche e, quindi, nel circolo portale, pertanto giungono al fegato in concentrazione più elevata che agli altri tessuti. Gli ormoni pancreatici più importanti sono il glucagone e l’insulina coinvolti nell’omeostasi glicemica (valori normali: ) in modo da contrapporsi.
 che secernono glucagone, le cellule b (75-80%) che secernono insulina, le cellule d (3-4%) e le cellule j (1-2%) che secernono rispettivamente la somatostatina (SS) e il polipeptide pancreatico (PP). Gli ormoni pancreatici sono riversati nelle vene pancreatiche e, quindi, nel circolo portale, pertanto giungono al fegato in concentrazione più elevata che agli altri tessuti. Gli ormoni pancreatici più importanti sono il glucagone e l’insulina coinvolti nell’omeostasi glicemica (valori normali: ) in modo da contrapporsi.")
62
Glucagone Il glucagone è un ormone peptidico trasportato in forma libera nel plasma, la cui secrezione è stimolata dall’ipoglicemia, mentre è inibita dall’iperglicemia. Le cellule a sono stimolate a secernere glucagone anche dagli amminoacidi ma se è deficitaria la disponibilità del glucosio. Il glucagone agisce soprattutto tramite il fegato, dove stimola le cellule epatiche a rilasciare glucosio attivando gli enzimi preposti alla glicogenolisi e alla gluconeogenesi. Contemporaneamente impedisce che le cellule epatiche utilizzino per se stesse il glucosio, attraverso la glicolisi e la glicogenosintesi. L’azione iperglicemizzante del glucagone si sovrappone a quella dell’adrenalina, del GH, degli ormoni tiroidei e dei glucocorticoidi. Nelle cellule epatiche, il glucagone stimola l’ingresso di aminoacidi glucogenetici dal plasma perché li trasformino in glucosio. Nel fegato il glucagone stimola, inoltre il metabolismo dei lipidi attivando la lipolisi, da cui sono sono ottenuti glicerolo per la gluconeogenesi, e NEFA per la b –ossidazione mentre è inibito il loro utilizzo nella sintesi dei trigliceridi. Per questi effetti lipolitici, il glucagone ha azione chetogenica. La funzione del glucagone è importante nel digiuno, nell’attività fisica, nelle situazioni di stress.

63
Insulina L’insulina è una proteina costituita da due catene lineari (catena A e catena B) unite da due ponti disolfuro. La liberazione in circolo dell’insulina è stimolata primariamente dalla concentrazione plasmatica del glucosio (iperglicemia) alla quale le cellule b sono specificatamente sensibili. L’azione dell’insulina è potenziata dagli aminoacidi, dai NEFA, dai corpi chetonici. L’efficacia di questi stimolatori, definiti secondari, dipende tuttavia dalla concentrazione ematica di glucosio, nel senso che la loro azione è bassa se la glicemia è bassa e viceversa. L’effetto ipoglicemizzante dell’insulina è ottenuto attraverso la la stimolazione della captazione del glucosio ematico all’interno dei tessuti insulino dipendenti (tessuto epatico, muscolare scheletrico e adiposo), la sua utilizzazione a scopo energetico (glicolisi) o nella sintesi e nel deposito del glicogeno nei tessuti epatico e muscolare , mentre negli stessi è inibita la glicogenolisi. Nel tessuto epatico è inibita anche la gluconeogenesi. L’insulina favorisce l’ingresso degli aminoacidi e dei NEFA rispettivamente nel muscolo scheletrico e nel tessuto adiposo, dove promuove la sintesi delle proteina e dei lipidi (trigliceridi) e inibisce la proteolisi e la lipolisi. L’insulina stimola nel fegato la sintesi dei trigliceridi, del colesterolo, delle proteine e delle lipoproteine a densità molto bassa (VLDL).
 unite da due ponti disolfuro. La liberazione in circolo dell’insulina è stimolata primariamente dalla concentrazione plasmatica del glucosio (iperglicemia) alla quale le cellule b sono specificatamente sensibili. L’azione dell’insulina è potenziata dagli aminoacidi, dai NEFA, dai corpi chetonici. L’efficacia di questi stimolatori, definiti secondari, dipende tuttavia dalla concentrazione ematica di glucosio, nel senso che la loro azione è bassa se la glicemia è bassa e viceversa. L’effetto ipoglicemizzante dell’insulina è ottenuto attraverso la la stimolazione della captazione del glucosio ematico all’interno dei tessuti insulino dipendenti (tessuto epatico, muscolare scheletrico e adiposo), la sua utilizzazione a scopo energetico (glicolisi) o nella sintesi e nel deposito del glicogeno nei tessuti epatico e muscolare , mentre negli stessi è inibita la glicogenolisi. Nel tessuto epatico è inibita anche la gluconeogenesi. L’insulina favorisce l’ingresso degli aminoacidi e dei NEFA rispettivamente nel muscolo scheletrico e nel tessuto adiposo, dove promuove la sintesi delle proteina e dei lipidi (trigliceridi) e inibisce la proteolisi e la lipolisi. L’insulina stimola nel fegato la sintesi dei trigliceridi, del colesterolo, delle proteine e delle lipoproteine a densità molto bassa (VLDL).")
64
Alterazioni funzionali
Diabete mellito insulino dipendente o di tipo I o diabete giovanile Diabete non insulino dipendente o di tipo II