Scaricare la presentazione
1
DNA: riparazione e ricombinazione

2
Causa delle mutazioni Errori di replicazione
Danni chimici. Il DNA è una molecola fragile: perdita o alterazioni di basi, rottura dello scheletro Inserimento di trasposoni .

3
Mutazioni Puntiformi Delezioni /inserzioni
Esposti alla mutazioni sono i microsatelliti: difficoltà a copiare sequenza ripetute, slippage Transizioni trasversioni

4
Riparazione del mismatch
Mismatch repair è parte della replicazione. In E. coli il DNA viene analizzato per distorsioni da Mut S (un dimero), lo distorce ulteriormente Recluta MutL, che recluta MutH che taglia il filamento vicino al mismatch Poi intervengono: elicasi, esonucleasi, DNA Pol (III) e ligasi.
, lo distorce ulteriormente. Recluta MutL, che recluta MutH che taglia il filamento vicino al mismatch. Poi intervengono: elicasi, esonucleasi, DNA Pol (III) e ligasi.")
5
Mismatch repair in E. coli
Come fa a identificare il filamento nuovo su cui operare il taglio ed escissione? Dalla metilazione della A nella sequenza CTAG ad opera della Dam metilasi MutS si lega al filamento non metilato

6
Mismatch repair in Eucarioti
Gli eucarioti hanno omologhi di Mut(s) con più alta specificità (per mismatch, indels etc) Non hanno Dam Metilasi, riconoscono il filamento stampo dalle incisioni dei frammenti di Okasaki. Sono associate al sliding clamp Mutazioni dei geni predispongono ai tumori
 con più alta specificità (per mismatch, indels etc) Non hanno Dam Metilasi, riconoscono il filamento stampo dalle incisioni dei frammenti di Okasaki. Sono associate al sliding clamp. Mutazioni dei geni predispongono ai tumori.")
7
Chimica delle mutazioni
Idrolisi: deaminazione di Citosina (anche adenosina e guanina) depurinazione
 depurinazione.")
8
La chimica delle basi del DNA facilita il riconoscimento dei danni al DNA
La deaminazione produce basi non-naturali che possono essere riconosciute, con una eccezione: la 5-metil citosina (la sequenza CG TG

9
Danni al DNA Sono indotti da radiazioni e sostanze mutagene
Alchilazione Attacco da forme reattive dell’ossigeno (OH.) UV: dimerizzazione delle pirimidine Raggi X: rompono la doppia elica Agenti intercalanti: inserzioni o delezioni
 UV: dimerizzazione delle pirimidine. Raggi X: rompono la doppia elica. Agenti intercalanti: inserzioni o delezioni.")
10
Analoghi ed agenti intercalanti
5-bromouracile nella forma enolica si lega a G Gli agenti intercalanti alterano lo spaziamento tra le basi e inducono inserzioni o delezioni

11
Se non corretti gli errori sono trasmessi

12
Sitemi di riparazione umani
Riparazione di mismatch Correzione diretta Excisione di basi (specifica) Excisione di nucleotidi
 Excisione di nucleotidi.")
13
Riparazione del DNA I danni causano una distorsione strutturale: cambio di forma di una molecola. Fotoriattivazione ripara da pyrimidine dimer che si formano per irradiazione con ultravioletto che genera un legame covalente tra due pirimidine adiacenti nel DNA. Blocca la replicazione. Excision repair: un sistema di riparazione in cui uno strand è exciso direttamente e poi sostituito usando lo stand complementare come templato. Recombination-repair: un modo di riempire un gap in un strand usando un single strand omologo di un altro duplex sintesi Error-prone quando il DNA incorpora basi non-complementari nello strand figlio.

14
Fotoriattivazione La DNA fotoliasi è attivata dalla luce e rompe i legami covalenti che uniscono le pirimidine

15
Meccanismo di escissione delle basi
Riconoscimento da base-flipping

16
Reazione dell’uracil glicosilasi
Prima interviene la glicosilasi e poi endo- e eso-nucleasi La glicosilasi di sicurezza sostituisce la A con C per riparare oxoG

17
Base flipping l’uracile e la basi alchilate sono riconosciute dalle glicosilasi e tolte direttamente dal DNA. I dimeri delle pirimidine sono tolti rompendo i legami covalenti tra di loro. Le metilasi aggiungono un gruppo metile alle citosine Tutti questi tipi di enzimi agiscono girando la base fuori dalla doppia elica dove può essere eliminata o modificata e riportata nella alfa elica

18
Meccanismo di escissione di nucleotidi
In E. coli proteine UvrA, -B, -C e –D UvrA-UvrB cercano distorsioni del DNA. UvrB fonde il DNA e recluta UvrC che taglia il DNA 8 nt a monte e 4 nt a valle. L’elicasi UvrD toglie il frammento tagliato Poi: DNA Pol I, ligasi Nell’uomo le proteine coinvolte causano lo Xeroderma pigmentoso, XP A-G

19
Trascrizione accoppiata alla riparazione del DNA.
Quando la RNA polimerasi trova un mismatch, si ferma. Recluta il sistema di riparazione per scissione nucleotidica e si distacca. Al sistema partecipa FTIIH che fa parte del fattore generale di trascrizione XPG eucariotica taglia un frammento di nt

20
Danni ai sistemi di riparazione causano tumori

21
Sintesi translesione La replicazione del DNA si blocca se trova una lesione. La Pol III si distacca e al suo posto entra La DNA pol translesione (Pol IV o Pol V). Copia il DNA con poca fedeltà, e poi si stacca per far posto alla Pol III. In E. coli queste polimerasi sono indotte dalla risposta SOS
. Copia il DNA con poca fedeltà, e poi si stacca per far posto alla Pol III. In E. coli queste polimerasi sono indotte dalla risposta SOS.")
22
Sintesi del DNA translesione

23
Risposta SOS Danni al DNA inducono RecA a scatenare la risposta SOS: l’espressione di geni che codificano molti enzimi di riparazione RecA attiva l’attività di auto-taglio di LexA. LexA reprime il sistema SOS; la sua degradazione induce la sintesi dei geni.

24
Le rotture alla doppia elica del DNA sono riparate efficientemente
Le cellule producono gli enzimi di riparazione in risposta al danno al DNA I danni la DNA rallentano la progressione del ciclo cellulare

25
Rotture di double strand
Non-homologous end-joining (NHEJ) lega terminazioni piatte. Lo si trova in meccanismi di riparazione e di ricombinazione (come la ricombinazione delle immunoglobuline). The NHEJ pathway può ligare le estremità piatte del DNAduplex. Mutazioni del NHEJ pathway causano malattie
 lega terminazioni piatte. Lo si trova in meccanismi di riparazione e di ricombinazione (come la ricombinazione delle immunoglobuline). The NHEJ pathway può ligare le estremità piatte del DNAduplex. Mutazioni del NHEJ pathway causano malattie.")
26
Tipi di ricombinazioni
Ricombinazione tra sequenze omologhe: (generalized o Homologous recombination) avviene prevalentemente nella meiosi. Avviene allo stadio di “four strand”. Ricombinazione tra paia di sequenze specifiche: site specific ricombination. Es. nell’integrazione dei fagi. Gli enzimi sono specifici per la sequenza Trasposizione: sequenze vengono inserite senza biosogno di omologia di sequenza Copy choice: usata dai virus RNA, la polimerasi passa dal templato ad un’altra sequenza
 avviene prevalentemente nella meiosi. Avviene allo stadio di four strand . Ricombinazione tra paia di sequenze specifiche: site specific ricombination. Es. nell’integrazione dei fagi. Gli enzimi sono specifici per la sequenza. Trasposizione: sequenze vengono inserite senza biosogno di omologia di sequenza. Copy choice: usata dai virus RNA, la polimerasi passa dal templato ad un’altra sequenza.")
27
La ricombinazione omologa
La ricombinazione omologa è coinvolta: Nel crossing over durante la meiosi Recuperare sequenze perse per danni al DNA Per far ripartire forche replicative danneggiate Può regolare l’espressione di alcuni geni. Produzione di animali Knock-out

28
Ricombinazione omologa
La ricombinazione permette di separare i vari geni nel genoma e di separare le mutazioni favorevoli da quelle sfavorevoli Le ricombinazioni avvengono tra sequenze che corrispondono perfettamente, così da non perdere nessuna base dal cromosoma ricombinante La frequenza non è costante ma varia con effetti globali e locali. Avviene 2 volte più frequentemente nella femmina che nel maschio Dipende dalla struttura del cromosoma ed il crossing-over non avviene in vicinanza di regioni condensate delle cromatina

29
La ricombinazione generale

30
Passaggi delle ric. omologa
Allineamento di due molecole di DNA omologhe Introduzione di rotture del DNA Formazione di una corta regione di appaiamento: invasione del filamento e formazione della Holliday junction Movimento della Holliday Junction - branch migration Taglio della giunzione: risoluzione

31
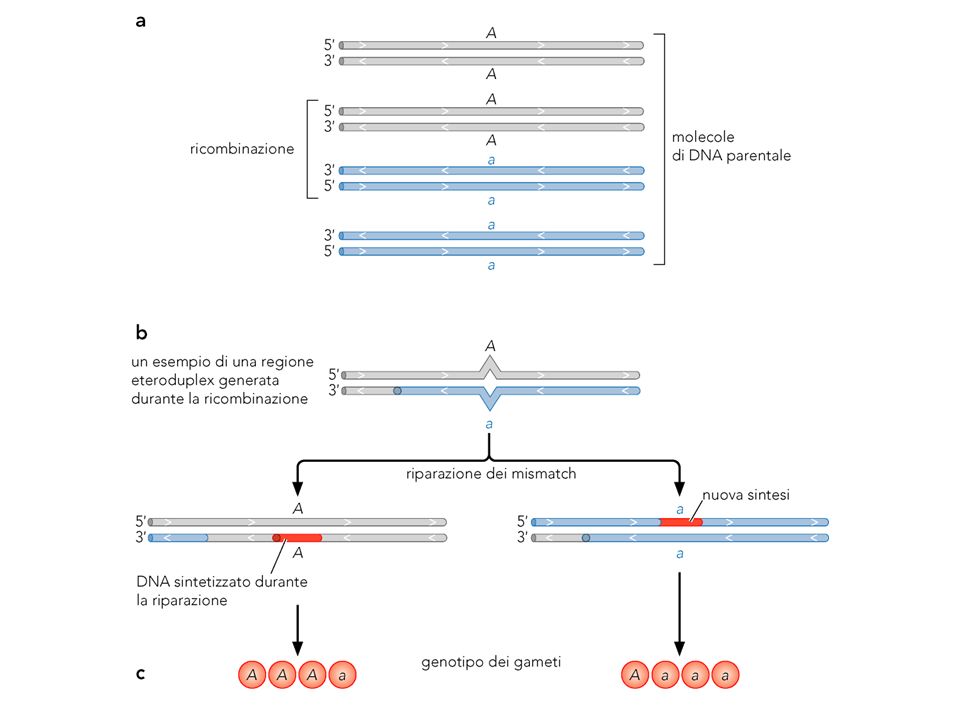
Modello di Holliday I DNA si appaiano, si ha un taglio ai siti corrispondenti, le terminazioni libere si complementano all’altro duplex: Joint molecule, e recombinant joint Il sito di ricombinazione: DNA ibrido o heteroduplex. Esso migra e deve essere catalizzato da enzimi La joint molecule deve essere risolta da due tagli. Se esso è sullo stesso strand si ottiene una zona heteroduplex: patch recombinant Se è sull’altro strand, tutti gli strand sono stati tagliati e si ha ricombinazione: splice recombinant. In in caso si ha una regione di heteroduplex

32
La complementarietà è a livello dei single strand
La complementarietà è a livello dei single strand. Essi sono accessibili dopo rottura del DNA. Uno strand spiazza (invade) quello corrispondente dell’altro duplex e i due duplex rimangono fisicamente uniti
 quello corrispondente dell’altro duplex e i due duplex rimangono fisicamente uniti.")
33
Holliday junction La giunzione può migrare in entrambe le direzioni

35
Risoluzione di una Holliday Junction
La risoluzione della giunzione di Holliday dà luogo a ricombinanti convenzionali o recombinant patches in base allo strand che è tagliato

36
Modello della Rottura del double strand (DSB)
Lo scambio genetico solitamente inizia con un taglio del double strand (double strand brake) Il DNA ricevente si taglia. Esonucleasi agiscono e liberano un 3’. Questo invade il DNA omologo e si estende. Si forma un D-loop. Il D loop si estende e si annila al DNA ricevente, e viene ricopiato Ci sono due regioni unite Il DNA del donatore ha sostituito quello dell’accettore
 Il DNA ricevente si taglia. Esonucleasi agiscono e liberano un 3’. Questo invade il DNA omologo e si estende. Si forma un D-loop. Il D loop si estende e si annila al DNA ricevente, e viene ricopiato. Ci sono due regioni unite. Il DNA del donatore ha sostituito quello dell’accettore.")
37
Risoluzione della riparazione del DSB
Se i tagli avvengono simmetricamente si risolve in un patch Se asimmetrici nella ricombinazione

38
Correzione degli errori di replicazione
A replication fork may stall when it encounters a damaged site or a nick in DNA. A stalled fork may reverse by pairing between the two newly synthesized strands. The structure of the stalled fork is the same as a Holliday junction and may be converted to a duplex and DSB by resolvases.

39
Meccanismi molecolari della RO
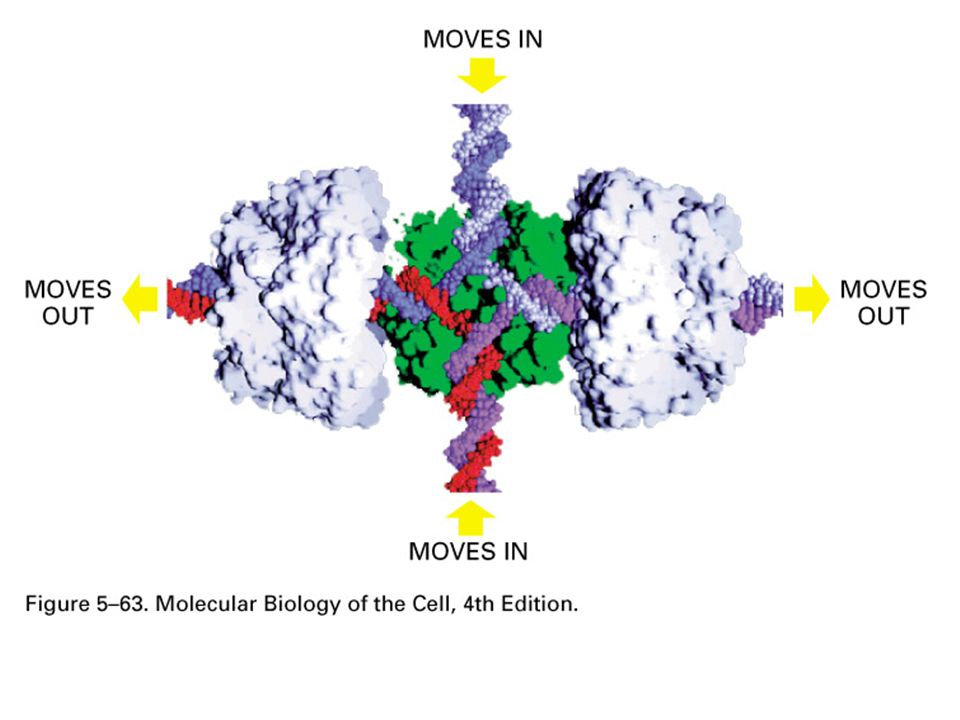
In E. coli il sistema di riparazione dei double stand breaks (DSB) è il sistema RecBCD Il complesso si lega a DSB, ha attività elicasica e nucleasica e taglia entrambi i filamenti. Quando raggiunge un sito chi (Cross-over Hotspot Instigator, CHI) digerisce solo con polarità 5’-3’ producendo una coda 3’ Il DNA senza siti viene digerito completamente RecA si lega al 3’- la proteina che scambia filamento RecA copre il 3’ in maniera cooperativa e catalizza l’appaiamento con il DNA omologo
 è il sistema RecBCD. Il complesso si lega a DSB, ha attività elicasica e nucleasica e taglia entrambi i filamenti. Quando raggiunge un sito chi (Cross-over Hotspot Instigator, CHI) digerisce solo con polarità 5’-3’ producendo una coda 3’ Il DNA senza siti viene digerito completamente. RecA si lega al 3’- la proteina che scambia filamento. RecA copre il 3’ in maniera cooperativa e catalizza l’appaiamento con il DNA omologo.")
40
I substrati di recA per lo scambio
Requisiti: DNA complementari tra i partner Una regione a singolo filamento La possibilità alla nuova elica di avvolgersi

41
RecA

42
RecA Modello per la ricerca del filamento omologo. Formazione della molecola giuntata Negli eucarioti Rad51 e Dmc1 Polarità dell’assemblaggio di RecA

43
RuvA, RuvB RuvAB riconosce la giunzione di Halliday e ne promuove la migrazione RuvA è un tetramero, RuvB un esamero con attività ATPasica. RuvC è una resolvasi che risolve il complesso tagliando in una delle due direzioni

44
RuvAB si lega alla Holliday J.
RuvA si lega specificamente alla Holliday J. Recluta RuvB che è un’ATPasi simile a elicasi, che muove la Halliday J. RuvC risolve le Holliday J.

45
Negli eucarioti Nei procarioti la RO serve per riparare le rotture del dsDNA, permettere alle forche bloccate di ripartire e ricombinare con DNA fagico o di coniugazione Negli eucarioti è anche necessaria per la meiosi per l’appaiamento dei cromosomi omologhi Ricombinazione meiotica

46
Ricombinazione meiotica: Spo11
Spo11 inizia la ricombinazione meiotica producendo dei tagli del dsDNA Taglia in zone non associate a nucleosomi ed attive

47
Nella meiosi Inizia quando una endonucleasi taglia il doppio strand. Una esonucleasi produce i 3’ protundenti. Questi trovano la regione omologa di un secondo cromosoma per formare la sinapsi

48
MRX processa le estremità tagliate
MRX è un complesso con attività esonucleasica. Degrada i filamenti con estremità 5’, attaccati as Spo11. Produce filamenti con estremità 3’. Dmc1 è l’omologo di RecA, specifico per la meiosi. La ricombinazione avviene tra cromosomi non-fratelli

49
RO negli eucarioti È richiesta per l’appaiamento dei cromosomi omologhi nella meiosi Durante la meiosi viene espresso SPO11 che introduce rotture del DNA a doppio filamento MRX degrada il filamento dal 5’ al 3’. Le estremità 3’ legano Rad51 o Dmc1 (omologhi di RecA)
")
50
L’appaiamento dei cromosomi è necessario per la ricombinazione
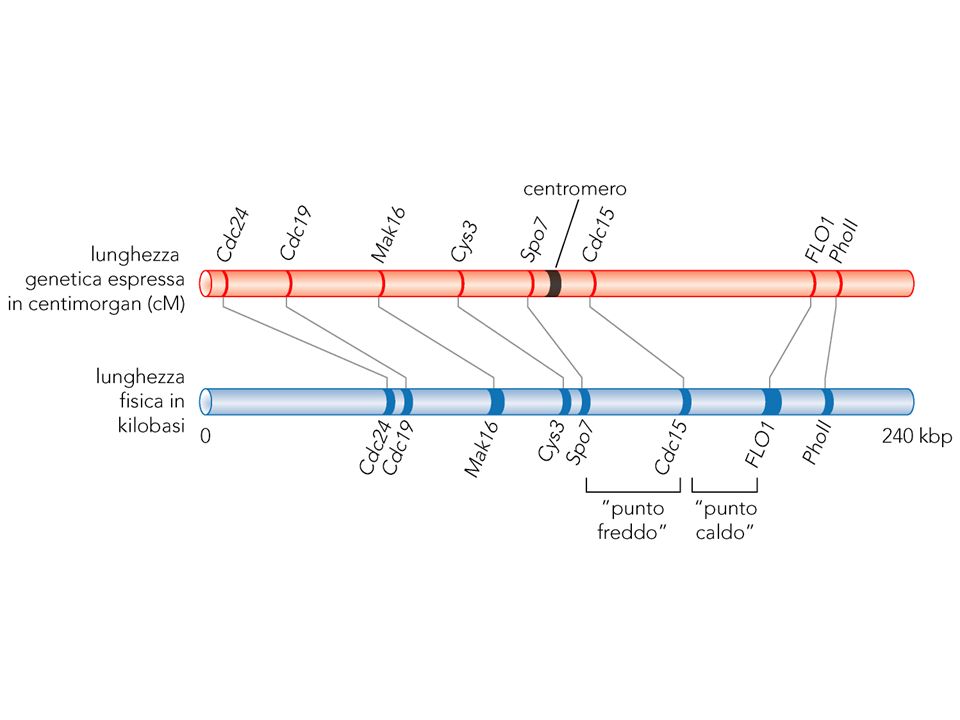
Rotture di doppio strand accadono presto nella meiosi in zone calde (hot spots) per la ricombinazione, spesso promotori o zone accessibili della cromatina La frequenza della ricombinazioni declina in entrambi i lati in un gradiente
 per la ricombinazione, spesso promotori o zone accessibili della cromatina. La frequenza della ricombinazioni declina in entrambi i lati in un gradiente.")
51
La conversione genetica nella meiosi

52
Conversione genetica per una correzione