Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Il Cavallo Rosso E. Corti (Ed. ARES)
Biotecnologie vegetali per l’industria e l’ambiente Biochimica classica - atto I Il Cavallo Rosso E. Corti (Ed. ARES)
")
2
Come manipolare il metabolismo?
È necessaria la comprensione del sistema metabolico e di come venga modulato il flusso Metà corso dedicato alla comprensione dei sistemi multienzimatici Metà corso a discutere degli interventi di ingegneria metabolica

3
Il metabolismo: questo sconosciuto
Un centinaio di anni di studio degli enzimi e dei metaboliti ci ha permesso di costruire accurate mappe metaboliche: E. Buchner: cell-free fermentation "Alcoholic Fermentation Without Yeast Cells". (1897) Ber. Dt. Chem. Ges. 30: 117–124.
 Ber. Dt. Chem. Ges. 30: 117–")
4
Flux is phenotype! (H. Kacser)
E’ possibile descrivere il metabolismo a partire dalle proprietà (studiate di solito in vitro) dei suoi componenti? vie metaboliche enzimologia Flux is phenotype! (H. Kacser) Teoria generale capace di predire i flussi?
 dei suoi componenti vie metaboliche. enzimologia. Flux is phenotype! (H. Kacser) Teoria generale capace di predire i flussi")
5
Cinetica di MM irreversibile
Breve ripasso di enzimologia Colmate le lacune con attenzione (ad es. Voet & Voet, Biochemistry) Cinetica di MM irreversibile E + S = ES E + P Derivazione della formula reperibile su tutti i testi (decenti) di biochimica Formula e curva sono la stessa cosa detta in due modi diversi!!!
 Cinetica di MM irreversibile. E + S = ES E + P. Derivazione della formula reperibile su tutti i testi (decenti) di biochimica. Formula e curva sono la stessa cosa detta in due modi diversi!!!")
6
Curva v/S Significato di KM e Vm; unità di misura

7
Molte rappresentazioni della cinetica sono sballate
Quando v raggiunge Vmax? A [S] infinito, cioè mai!

8
Linearizzazione y = 0 x = 0 Esistono tipi diversi di linearizzazione (vedi Voet e Voet o Fell)
")
9
Inibizione competitiva
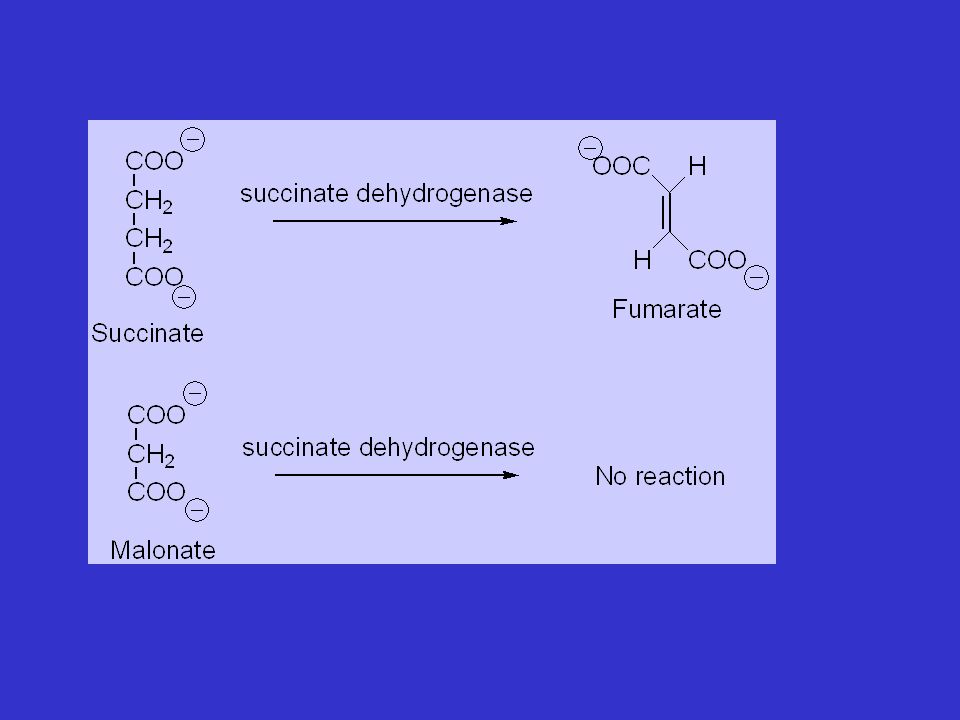
E + S = ES E + P + I Ki es. Succinato deidrogenasi: fumarato (S) e malonato (I) EI + S = EIS no reaction Il valore di Km viene aumentato: KMapp = KM(1+I/Ki)= KMα L’inibizione si annulla aumentando la concentrazione del substrato Che faccià avrà la linearizzazione in presenza di inibitore?
 e malonato (I) EI + S = EIS no reaction. Il valore di Km viene aumentato: KMapp = KM(1+I/Ki)= KMα. L’inibizione si annulla aumentando la concentrazione del substrato. Che faccià avrà la linearizzazione in presenza di inibitore")
11
Inibizione incompetitiva
E + S = ES E + P + I Ki es. EPSP sintasi (con glifosate) EIS no reaction Il valore di Vm viene diminuito: Vmapp= Vm/(1+I/Ki)= Vm/α’ L’inibizione, inefficace a basse [S], non si annulla ad alte [S] Che faccià avrà la linearizzazione in presenza di inibitore?
 EIS no reaction. Il valore di Vm viene diminuito: Vmapp= Vm/(1+I/Ki)= Vm/α’ L’inibizione, inefficace a basse [S], non si annulla ad alte [S] Che faccià avrà la linearizzazione in presenza di inibitore")
12
Fosfoenolpiruvato (PEP) 3-enolpiruvil-3-shikimato -fostato (EPSP)
COO - -2 3 PO C 2 OOC N 2- Shikimato-3-fosfato Fosfoenolpiruvato (PEP) Glifosate 3-enolpiruvil-3-shikimato -fostato (EPSP) EPSP sintasi -2 O3 P Reazione della EPSP sintasi e sua inibizione da parte del glifosate Nonostante assomigli molto a parte dello shikimato, il glifosate è un inibitore competitivo del PEP. La reazione è sequenziale, con lo shikimato che entra per primo, per cui il glifosate risulta essere un inibitore incompetitivo di questo primo substrato. Questo causa l’accumulo di Shikimato e la riduzione in Arogenato (che è un inibitore della DHAP sintasi che quindi esacerba l’accumulo di Shikimato) (vedi note)
 Glifosate. 3-enolpiruvil-3-shikimato -fostato (EPSP) EPSP sintasi. -2 O3 P. Reazione della EPSP sintasi e sua inibizione da parte del glifosate. Nonostante assomigli molto a parte dello shikimato, il glifosate è un inibitore competitivo del PEP. La reazione è sequenziale, con lo shikimato che entra per primo, per cui il glifosate risulta essere un inibitore incompetitivo di questo primo substrato. Questo causa l’accumulo di Shikimato e la riduzione in Arogenato (che è un inibitore della DHAP sintasi che quindi esacerba l’accumulo di Shikimato) (vedi note) idinformationmodule= &topicorder=5&maxto=7&minto=1.")
13
Inibizione mista E + S = ES E + P + EI EIS no reaction
Ki K’i EIS no reaction EI Un caso particolare della Inibiz. Mista è quella non-competitiva dove alfa = alfa’ Quindi non c’è effetto sulla Km Sia Vm che Km cambiano: Vmapp = Vm/α′ e KM′ = α KM/α′ L’inibizione si sente sia a bassa che ad alta concentrazione di S Che faccia avrà la linearizzazione in presenza di inibitore?

14
Le inibizioni dal punto di vista grafico

15
Equazione MM reversibile
Per una dimostrazione elegante:

16
Due conseguenze importanti:
La v della reazione reversibile sarà minore di quella irreversibile per due motivi: L’equazione per la MM reversibile è derivabile come quella irreversibile La presenza di prodotto rende più piccolo il numeratore La presenza di prodotto rende più grande il denominatore

17
La relazione di Haldane
All’equilibrio v = 0 per cui il numeratore sarà = 0 = Keq

18
Le costanti cinetiche non sono indipendenti!
Ricordatevi che il valore della Keq è indipendente dalla presenza dell’enzima Una formulazione alternativa dell’equazione si ottiene sfruttando la relazione di Haldane (sostituisco Vr) Prendete l’equazione della MM rev e sostituite a Vr l’espressione sulla sinistra
 Prendete l’equazione della MM rev e sostituite a Vr l’espressione sulla sinistra.")
19
Quando la Keq >>1 )
(cioè una reazione molto spostata verso i prodotti) ) L’effetto di P si fa sentire solo sul denominatore (si parla di inibizione da prodotto)
 ) L’effetto di P si fa sentire solo sul denominatore. (si parla di inibizione da prodotto)")
20
Inoltre se [P]/KP è bassa ( 0 o comunque <<1)
Che altro non è se non l’equazione per la reazione irreveversibile
![Inoltre se [P]/KP è bassa ( 0 o comunque <<1)](http://slideplayer.it/slide/193293/1/images/20/Inoltre+se+%5BP%5D%2FKP+%C3%A8+bassa+%28%EF%81%80+0+o+comunque+%3C%3C1%29.jpg "Che altro non è se non l’equazione per la reazione irreveversibile.")
21
Importantissimo L’equazione di MM irreversibile non descrive il comportamento in vivo perché la maggior parte degli enzimi catalizzano reazioni reversibili. Quella reversibile è cruciale per descrivere i sistemi reali Inoltre la maggior parte delle reazioni enzimatiche hanno più di un substrato o di un prodotto per cui si richiedono equazioni più complesse.

22
Enzimi a più substrati/prodotti
A + B C + D Equaz. generale (irrev.) Classificati secondo il meccanismo: *Random order * Compulsory order * Ping Pong Sequenziali
 Classificati secondo il meccanismo: *Random order. * Compulsory order. * Ping Pong. Sequenziali.")
23
http://employees. csbsju
Enzimi allosterici: Molti enzimi non hanno curve iperboliche, ma sigmoidi, per cui sono stati introdotte teorie per spiegare questi comportamente (Hill, MWC e Koshland) v =Vm*(x*(1+x)n-1+L*c*x*(1+c*x)n-1)/((1+x)n + L*(1+c*x)n) [MWC model] Andate a rivederli sul testo di biochimica !
 v =Vm*(x*(1+x)n-1+L*c*x*(1+c*x)n-1)/((1+x)n + L*(1+c*x)n) [MWC model] Andate a rivederli sul testo di biochimica !")
24
atto II (ovverosia la termodinamica)
Dopo le nozioni essenziali di enzimologia, ripassiamo alcuni concetti di termodinamica che sono rilevanti per le vie metaboliche

25
All’eq. G= 0 e G°' = –RT lnKeq
Si raggiunge sempre uno stato stazionario (ss)? Che conseguenze avrebbe non raggiungere uno ss… La presenza di un flusso in una via metabolica cosa implica dal punto di vista termodinamico? Un G ≠ 0 (<0) G = G°' + RT lnQ Q = quoziente di reaz. Q (chiamato anche Γ) è il rapporto conc. dei prodotti/conc. reagenti All’eq. G= 0 e G°' = –RT lnKeq G°' = –RT lnKeq = –2.3RT Log10(Keq) G°' è la variazione di energia libera in condizioni standard
 Che conseguenze avrebbe non raggiungere uno ss… La presenza di un flusso in una via metabolica cosa implica dal punto di vista termodinamico Un G ≠ 0 (<0) G = G° + RT lnQ. Q = quoziente di reaz. Q (chiamato anche Γ) è il rapporto conc. dei prodotti/conc. reagenti. All’eq. G= 0 e G° = –RT lnKeq. G° = –RT lnKeq = –2.3RT Log10(Keq) G° è la variazione di energia libera in condizioni standard.")
26
Ordine di grandezza Da Lehninger
ATP + H2O ADP + Pi Go' = -31 kJoules/mol Pi + glucose glucose-6-P + H2O Go' = +14 kJoules/mol

27
Values of Go and Keq for common reactions at 25oC
SO3(g) 2 SO2(g) + O2(g) 141.7 1.4 x 10-25 H2O(l) H+(aq) + OH-(aq) 79.9 1.0 x 10-14 AgCl(s) + H2O Ag+(aq) + Cl-(aq) 55.6 1.8 x 10-10 HOAc(aq) + H2O H+(aq) + OAc-(aq) 27.1 1.8 x 10-5 N2(g) + 3 H2(g) 2 NH3(g) -32.9 5.8 x 105 HCl(aq) + H2O H+(aq) + Cl-(aq) -34.2 1 x 106 Cu2+(aq) + 4 NH3(aq) Cu(NH3)42+(aq) -76.0 2.1 x 1013 Zn(s) + Cu2+(aq) Zn2+(aq) + Cu(s) -211.8 1.4 x 1037 Reaction Go(KJ/mol) Keq R = [15] J · K-1 · mol Go (kJ/mol)
 2 SO2(g) + O2(g) x H2O(l) H+(aq) + OH-(aq) x AgCl(s) + H2O. Ag+(aq) + Cl-(aq) x HOAc(aq) + H2O. H+(aq) + OAc-(aq) x N2(g) + 3 H2(g) 2 NH3(g) x 105. HCl(aq) + H2O. H+(aq) + Cl-(aq) x 106. Cu2+(aq) + 4 NH3(aq) Cu(NH3)42+(aq) x Zn(s) + Cu2+(aq) Zn2+(aq) + Cu(s) x Reaction Go(KJ/mol) Keq. R = [15] J · K-1 · mol-1 Go (kJ/mol)")
28
Quoziente di reazione e Keq

29
G = G°' + RT lnQ = –RT lnKeq+ RT lnQ =
= RT ln(Q/Keq) = RT ln ρ (con ρ Disequilibrium ratio) ρ Che valori assume ρ? Ovviamente tra 0 e 1 per G <0 Quanto più ρ è vicino a 0 tanto più G < 0 (reaz. spontanea) Dovete capire bene la differenza tra G e G° ' ρ (o G, che concettualmente è la stessa cosa) è considerato da molti un indice potenziale di regolazione: *se ρ è molto piccolo (0) si ritiene che l’enzima sia un potenziale sito di regolazione (se ρ 1 c’è poca possibilità di regolazione)
 = RT ln ρ. (con ρ Disequilibrium ratio) ρ. Che valori assume ρ Ovviamente tra 0 e 1 per G <0. Quanto più ρ è vicino a 0 tanto più G < 0 (reaz. spontanea) Dovete capire bene la differenza tra G e G° ρ (o G, che concettualmente è la stessa cosa) è considerato da molti un indice potenziale di regolazione: *se ρ è molto piccolo (0) si ritiene che l’enzima sia un potenziale sito di regolazione (se ρ 1 c’è poca possibilità di regolazione)")
30
Il loro rapporto vale (=Q), da cui posso calcolarmi G

31
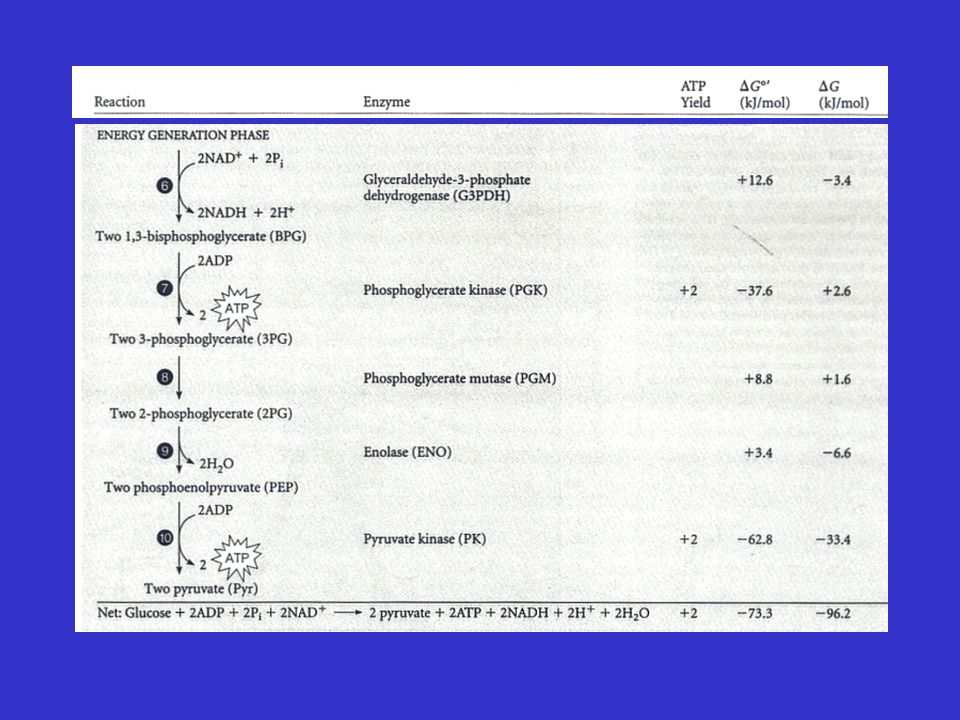
ΔG°' e ΔG in vivo nella glicolisi

33
Perché ρ è considerato un indice di potenziale regolazione?
I metaboliti devono essere a concentrazioni nel range μM- mM A) Non possono essere a concentrazioni molto alte (es. 1M) * perché ci sarebbero effetti osmotici pesanti * perché le variazioni nel metabolismo sarebbero lente (ci sarebbe molto intermedio da smaltire) * perché si investirebbe molto in intermedi (e non prodotti)
 Non possono essere a concentrazioni molto alte (es. 1M) * perché ci sarebbero effetti osmotici pesanti. * perché le variazioni nel metabolismo sarebbero lente (ci sarebbe molto intermedio da smaltire) * perché si investirebbe molto in intermedi (e non prodotti)")
34
Kcat = Vmax/Etot = turnover number
B) Non possono essere a concentrazioni molto basse (es. nM) * perché gli enzimi avrebbero Km basse e quindi una bassa efficacia catalitica: Kcat / Km vale al max * perché gli intermedi verrebbero esauriti velocemente (non ci sarebbero riserve efficaci di metaboliti) kcat is called the turnover number kcat = moli di substrate trasformato / mole di enzima/secondo Kcat = Vmax/Etot = turnover number n. di reazioni catalizzate da ogni sito attivo ogni secondo; per un enzima MM vale K2. Non può essere maggiore della frequenza con cui substrato ed enzima si incontrano. Se ogni incontro è produttivo, si avrà la perfezione catalitica
 Non possono essere a concentrazioni molto basse (es. nM) * perché gli enzimi avrebbero Km basse e quindi una bassa efficacia catalitica: Kcat / Km vale al max * perché gli intermedi verrebbero esauriti velocemente (non ci sarebbero riserve efficaci di metaboliti) kcat is called the turnover number. kcat = moli di substrate trasformato / mole di enzima/secondo. Kcat = Vmax/Etot = turnover number. n. di reazioni catalizzate da ogni sito attivo ogni secondo; per un enzima MM vale K2. Non può essere maggiore della frequenza con cui substrato ed enzima si incontrano. Se ogni incontro è produttivo, si avrà la perfezione catalitica.")
35
Quando [S] << KM (soluzioni diluite) ricaviamo
Pendenza della retta ammettendo E= Etot : = (k2/KM)[E][S] ~ (kcat/KM)[E][S] Rate constant per reaz. Bimolecolare Non può essere maggiore della velocità d’incontro
![Quando [S] << KM (soluzioni diluite) ricaviamo](http://slideplayer.it/slide/193293/1/images/35/Quando+%5BS%5D+%3C%3C+KM+%28soluzioni+diluite%29+ricaviamo.jpg "Pendenza della retta. ammettendo E= Etot : = (k2/KM)[E][S] ~ (kcat/KM)[E][S] Rate constant per reaz. Bimolecolare. Non può essere maggiore della velocità d’incontro.")
36
(kcat/KM) vale al massimo 108-109 cioè il valore massimo compatibile con la diffusione
Diffusion theory predicts that kcat/KM will attain a value of about (mol/L)-1s-1. Carbonic anhydrase, Catalase, Fumarase and Triose phosphate isomerase actually approach this limit. Sulla TIM e in generale sulla Glicolisi TIM accelerates isomerization by a factor of 1010 compared with the rate obtained with a simple base catalyst such as acetate ion. Indeed, the kcat/KM ratio for isomerization of glyceraldehyde 3-phosphate is 2 × 108 M-1 s-1.
-1s-1. Carbonic anhydrase, Catalase, Fumarase and Triose phosphate isomerase actually approach this limit. Sulla TIM e in generale sulla Glicolisi. TIM accelerates isomerization by a factor of 1010 compared with the rate obtained with a simple base catalyst such as acetate ion. Indeed, the kcat/KM ratio for isomerization of glyceraldehyde 3-phosphate is 2 × 108 M-1 s-1.")
37
Da Lehninger The kcat and activation energy of Rubisco from a variety of C3 and C4 plants Sage et al. J. Exp. Bot. (2002) 53: Sage et al.

38
Se questi 2 ragionamenti sono veri, ne consegue che :
* quando G° ' <<0 la reazione deve rimanere lontana dell’eq. * quando G° ' 0 (Keq 1) la reazione deve rimanere vicina all’eq. G <<0 G 0 Cosa significa (approssimativamente)? Cosa succederebbe infatti se le reazioni con G° ' <<0 andassero all’equilibrio?
 la reazione deve rimanere vicina all’eq. G <<0. G 0. Cosa significa (approssimativamente) Cosa succederebbe infatti se le reazioni con G° <<0 andassero all’equilibrio")
39
[P] >> [S] Con due reazioni di questo tipo in fila (come ad es. nella glicolisi GG6PF6PF1,6BP*), allora: [S] << [P1] [P2] << [P3] e più precisamente: ([F1,6BP][ADP]2 / [G][ATP]2) = 106 * La prima e terza reazione hanno un G grande e negativo. E cosa succederebbe se le reazioni con G° ' 0 fossero lontane dall’equilibrio… [P] << [S]
![[P] >> [S] Con due reazioni di questo tipo in fila (come ad es. nella glicolisi GG6PF6PF1,6BP*), allora:](http://slideplayer.it/slide/193293/1/images/39/%5BP%5D+%3E%3E+%5BS%5D+Con+due+reazioni+di+questo+tipo+in+fila+%28come+ad+es.+nella+glicolisi+G%EF%83%A0G6P%EF%83%A0F6P%EF%83%A0F1%2C6BP%2A%29%2C+allora%3A.jpg "[S] << [P1] [P2] << [P3] e più precisamente: ([F1,6BP][ADP]2 / [G][ATP]2) = 106. * La prima e terza reazione hanno un G grande e negativo. E cosa succederebbe se le reazioni con G° 0 fossero lontane dall’equilibrio… [P] << [S]")
40
Riassumendo: Le concentrazioni dei metaboliti possono rimanere dentro ambiti sensati (μM-mM) se le reazioni con * G° ' <<0 rimangono lontane dall’equilibrio (G <<0, vale a dire ρ 0). Tali reazioni quindi devono essere ben regolate! * G° ' 0 rimangono vicine all’equilibrio (G 0) Queste reazioni devono essere catalizzate da enzimi in eccesso (altrimenti rallentando rimarrebbero lontane dall’equilibrio)
. Tali reazioni quindi devono essere ben regolate! * G° 0 rimangono vicine all’equilibrio (G 0) Queste reazioni devono essere catalizzate da enzimi in eccesso (altrimenti rallentando rimarrebbero lontane dall’equilibrio)")
41
TPI aumenta la velocità della reazione 109 volte!!
Da Lehninger TPI aumenta la velocità della reazione 109 volte!!

42
Referenze Per la cinetica enzimatica vedere
* Testo di biochimica (ad es. Voet and Voet, Biochemistry) * Fell “understanding the control of Metabolism”, 1997 cap. 3 Per la parte di cinetica chimica vedere: * Fell “understanding the control of Metabolism”, 1997 cap e 1.4.3
 * Fell understanding the control of Metabolism , 1997 cap. 3. Per la parte di cinetica chimica vedere: * Fell understanding the control of Metabolism , 1997 cap e")
43
Effect of reversible inhibitors on the reaction rate: � no inhib. a)� mixed inhib. ([I] = Ki = 0.5 Ki'); lower Vmaxapp (= 0.67 Vmax), higher Kmapp (= 2 Km). b)� competitive inhib. ([I] = Ki); Vmaxapp unchanged (= Vmax), higher Kmapp (= 2 Km). c)� uncompetitive inhib. ([I] = Ki'); lower Vmaxapp (= 0.5 Vmax) and Kmapp (= 0.5 Km). d)� noncompetitive inhib. ([I] = Ki = Ki'); lower Vmaxapp (= 0.5 Vmax), unchanged Kmapp (= Km).
� mixed inhib. ([I] = Ki = 0.5 Ki ); lower Vmaxapp (= 0.67 Vmax), higher Kmapp (= 2 Km). b)� competitive inhib. ([I] = Ki); Vmaxapp unchanged (= Vmax), higher Kmapp (= 2 Km). c)� uncompetitive inhib. ([I] = Ki ); lower Vmaxapp (= 0.5 Vmax) and Kmapp (= 0.5 Km). d)� noncompetitive inhib. ([I] = Ki = Ki ); lower Vmaxapp (= 0.5 Vmax), unchanged Kmapp (= Km).")
44
A special case of uncompetitive inhibition is substrate inhibition which occurs at high substrate concentrations in about 20% of all known enzymes (e.g. invertase is inhibited by sucrose). It is primarily caused by more than one substrate molecule binding to an active site meant for just one, often by different parts of the substrate molecules binding to different subsites within the substrate binding site. If the resultant complex is inactive this type of inhibition causes a reduction in the rate of reaction, at high substrate concentrations. It may be modelled by the following scheme:

45
Effect of substrate inhibition the reaction rate
Effect of substrate inhibition the reaction rate. A comparison is made between the inhibition caused by increasing KS relative to Km. � no inhibition, KS/Km >> 100; � KS/Km = 100; � KS/Km = 10; � KS/Km = 1. By the nature of the binding causing this inhibition, it is unlikely that KS/Km < 1.

46
Competitiva

47
Incompetitiva

48
Quando α = α‘ si parla di in. non-competitiva (la Km non cambia)
Mista Quando α = α‘ si parla di in. non-competitiva (la Km non cambia)
")
49
Effetto degli inibitori sui parametri cinetici

Presentazioni simili















 Acidi poliprotici>")



