Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
ANTIBIOTICI DELLA PARETE

2
STRUTTURA DELLA PARETE DEI BATTERI

3
Sintesi dei monomeri di mureina
Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan.")
4
L-Ala D-Glu L-Lys Gly D-Ala UDP-Acido N-acetilmuramico

5
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina

6
a | Assembly of the cell-wall subunit takes place on the cytosolic side of the bacterial plasma membrane. UDP-activated precursor sugars are assembled on a polyisoprenoid carrier, the coupling of which produces Lipid II, which is then transported across the membrane by an unknown mechanism. Next, the peptidoglycan subunit is transferred to the growing peptidoglycan chain and the polyisoprenoid carrier is recycled back to the cytoplasmic side to complete the cycle. b | Structure of Lipid II. The polyisoprenoid anchor consists of eight isoprene units in the cis-conformation followed by two units in the trans-conformation and the terminal isoprene unit. Depicted here is a Lipid II carrying a pentapeptide with a lysine at the third position. Together with diaminopimelic acid, this is the most common residue at this position. The third amino acid of the pentapeptide is coupled to the glutamate at position two via the side chain carboxylate. The red bars indicate the minimal binding sites in Lipid II of glycopeptide antibiotics (1), nisin (2), ramoplanin (3) and mersacidin (4). GlcNAc, N-acetylglucosamine; MurNAc, N-acetylmuramic acid.
, nisin (2), ramoplanin (3) and mersacidin (4). GlcNAc, N-acetylglucosamine; MurNAc, N-acetylmuramic acid..")
7
Sintesi dei monomeri di mureina
BP-NAM-NAG Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan.")
8
Traslocazione dei monomeri di mureina
Fig. 1. Enzymatic steps in cell wall biosynthesis, with antibacterial sites of action indicated. (a) MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3'-hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come?, Pages Steven J. Projan
 MurA–F perform the cytoplasmic steps in peptidoglycan biosynthesis. The process of bacteria cell wall biosynthesis starts with the MurA transferase, which transfers the addition of an enolpyruvyl moiety to 3 -hydroxyl-UDP-NAG. Subsequently, the MurB reductase reduces the enol ether to a lactyl ether, utilizing one equivalent of NADPH and a solvent proton to form UDP-NAM. Next, a series of ATP-dependent amino acid ligases (MurC, MurD, MurE and MurF) catalyze the stepwise synthesis of the pentapeptide side-chain using the newly synthesized carboxylate as the first acceptor site. Each enzyme is responsible for the addition of one residue except for MurF, which catalyzes the addition of the dipeptide d-Ala-d-Ala. MurE in Gram-negative bacteria catalyzes the meso-2,6-diaminopimelate (DAP) addition, whereas in Gram-positive bacteria MurE catalyzes l-lysine addition. (b) The product of MurF, UDP-NAM-pentapeptide (NAM now abbreviated to M), is then catalyzed at the plasma membrane by the translocase MraY to transfer the phospho-UDP-NAM-pentapeptide moiety to the membrane acceptor undecaprenyl phosphate to form Lipid I. Then the MurG transferase catalyzes the addition of UDP-NAG (NAG now abbreviated to G) yielding UDP-NAM-(pentapeptide)-pyrophosphoryl-undecaprenol (Lipid II). Lipid II is then moved across the membrane into periplasm and catalyzed by the transglycosylase (TG) and the transpeptidase (TP) to form the extensively cross-linked polymer peptidoglycan essential for the integrity of the bacterial cell wall. The large penicillin binding proteins (PBPs) contain both TG and TP domains. New (and not so new) antibacterial targets – from where and when will the novel drugs come , Pages Steven J. Projan.")
9
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina Formazione di polimeri lineari di mureina (transglicosilazione) NAM Gly5 NAG NAM Gly5 NAG NAM Gly5 NAG NAM Gly5 NAG
 NAM. Gly5. NAG. NAM. Gly5. NAG. NAM. Gly5. NAG. NAM. Gly5. NAG.")
10
ANTIBIOTICI GLICOPEPTIDICI
Dhpg, dihydroxyphenylglycine; Glc, glucose; Hpg, hydroxyphenylglycine; Van, vancosamine.

11
The VanA gene cluster that confers vancomycin resistance
The VanA gene cluster that confers vancomycin resistance. A TWO-COMPONENT REGULATORY system VanR–VanS regulates vancomycin resistance in vancomycin-resistant ENTEROCOCCI (VRE) and vancomycin-resistant Staphylococcus aureus (VRSA) strains. VanS is a membrane-associated sensor (of vancomycin) that controls the level of phosphorylation of VanR. VanR is a transcriptional activator of the operon encoding VanH, VanA and VanX. VanH is a dehydrogenase that reduces pyruvate to D-Lac, whereas VanA is a ligase that catalyses the formation of an ester bond between D-Ala and D-Lac. Vancomycin does not bind to D-Ala-D-Lac, which leads to vancomycin resistance. VanX is a dipeptidase that hydrolyses the normal peptidoglycan component D-Ala-D-Ala, which prevents it from causing vancomycin sensitivity. VanY is a D,D-carboxypeptidase that hydrolyses the terminal D-Ala residue of late peptidoglycan precursors that are produced if elimination of D-Ala-D-Ala by VanX is not complete. So, D-Ala-D-Lac replaces the normal dipeptide D-Ala-D-Ala in peptidoglycan synthesis resulting in vancomycin resistance150. VanZ confers resistance to teicoplanin by an unknown mechanism.
 and vancomycin-resistant Staphylococcus aureus (VRSA) strains. VanS is a membrane-associated sensor (of vancomycin) that controls the level of phosphorylation of VanR. VanR is a transcriptional activator of the operon encoding VanH, VanA and VanX. VanH is a dehydrogenase that reduces pyruvate to D-Lac, whereas VanA is a ligase that catalyses the formation of an ester bond between D-Ala and D-Lac. Vancomycin does not bind to D-Ala-D-Lac, which leads to vancomycin resistance. VanX is a dipeptidase that hydrolyses the normal peptidoglycan component D-Ala-D-Ala, which prevents it from causing vancomycin sensitivity. VanY is a D,D-carboxypeptidase that hydrolyses the terminal D-Ala residue of late peptidoglycan precursors that are produced if elimination of D-Ala-D-Ala by VanX is not complete. So, D-Ala-D-Lac replaces the normal dipeptide D-Ala-D-Ala in peptidoglycan synthesis resulting in vancomycin resistance150. VanZ confers resistance to teicoplanin by an unknown mechanism.")
12
ANTIBIOTICI PEPTIDICI

13
SINTESI DELLA PARETE DEI BATTERI
Sintesi dei monomeri di mureina Traslocazione dei monomeri di mureina Formazione di polimeri lineari di mureina (transglicosilazione) Transpeptidazione (formazione di legami crociati tra polimeri lineari)
 Transpeptidazione (formazione di legami crociati tra polimeri lineari)")
14
Gly Antibiotici -lattamici Glicopeptide polimero
L-Ala D-Glu L-Lys Gly D-Ala Acido N-acetilmuramico Glicopeptide polimero Acido N-acetilmuramico Glicopeptide polimero Antibiotici -lattamici Transpeptidasi Acido N-acetilmuramico Glicopeptide polimero

16
Meccanismo d’azione degli antibiotici -lattamici
Interazione con le penicillin-binding proteins (PBPs) Inibizione della reazione di transpeptidazione Inattivazione di un inibitore di enzimi autolitici della parete cellulare (p.es. mureina idrolasi)
 Inibizione della reazione di transpeptidazione. Inattivazione di un inibitore di enzimi autolitici della. parete cellulare (p.es. mureina idrolasi)")
19
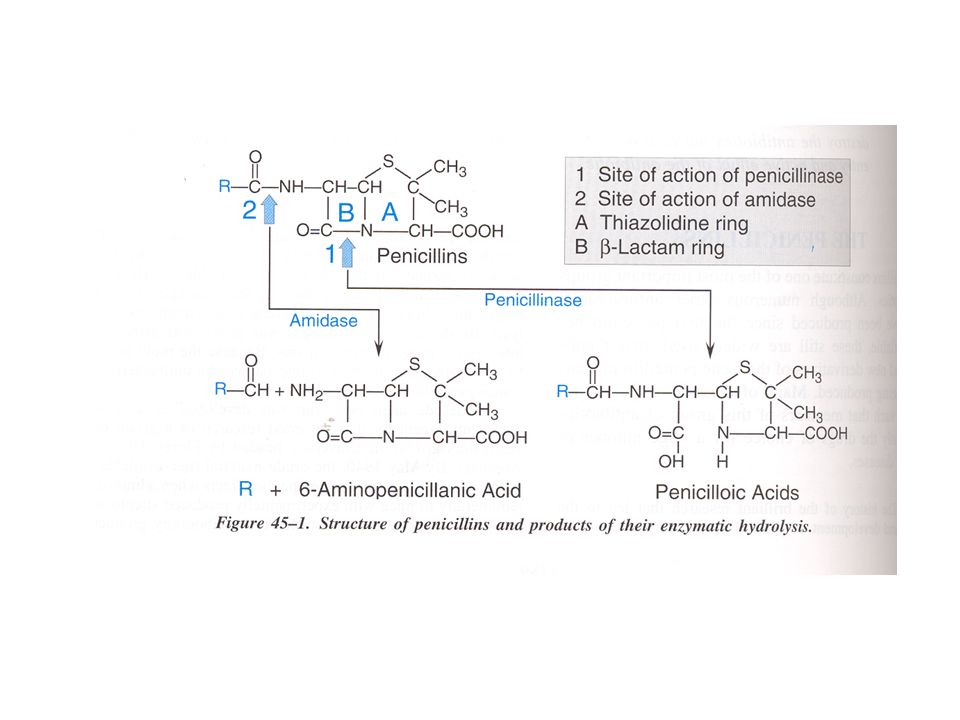
CLASSIFICAZIONE DELLE PENICILLINE: 1 - PENICILLINE NATURALI
ANELLO -LATTAMICO ANELLO TIAZOLIDNICO ACIDO 6-AMINOPENICILLANICO

20
Meccanismi di secrezione degli acidi (A) e delle
basi organiche (B) nel tubulo prossimale
 nel tubulo prossimale.")
21
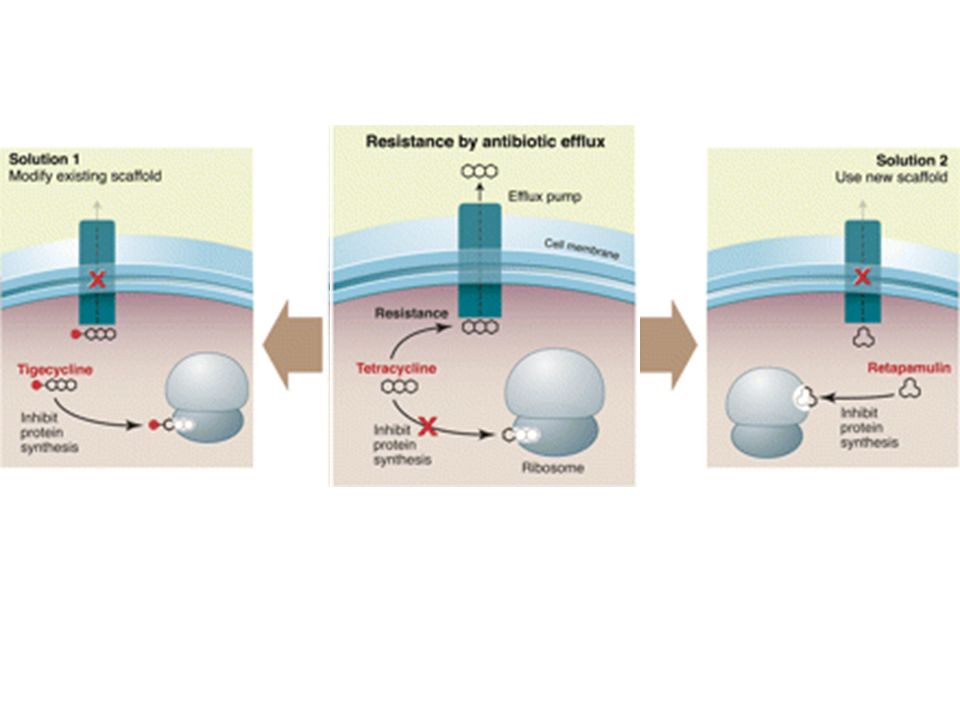
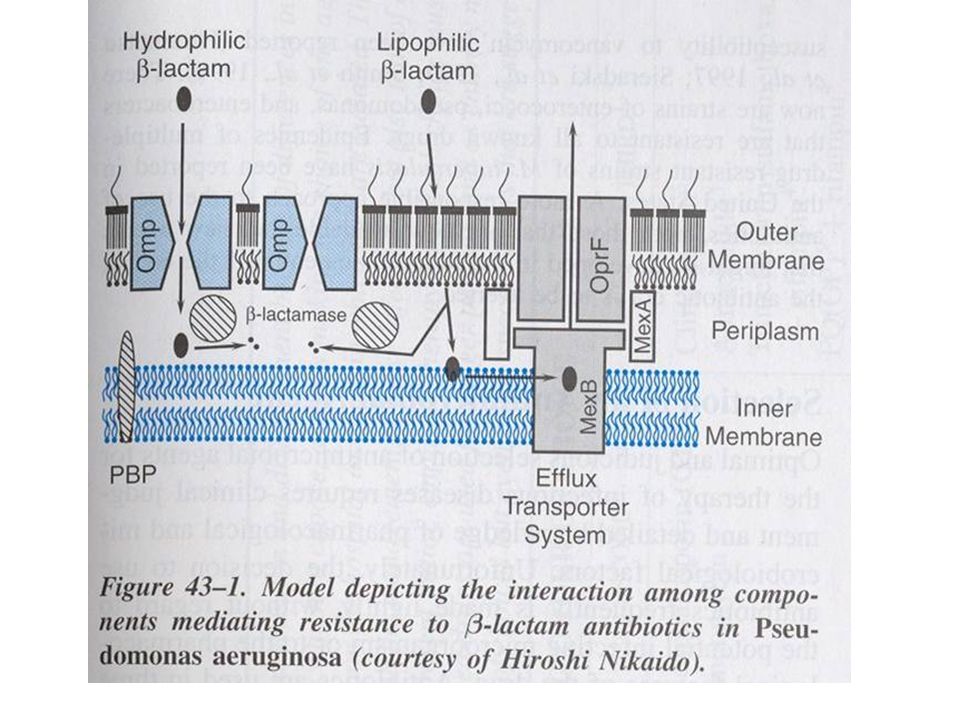
Meccanismi di resistenza agli antibiotici -lattamici
Diminuita espressione di porine (batteri Gram -) Espressione di pompe in grado di determinare l’efflusso del farmaco
 Espressione di pompe in grado di determinare l’efflusso del farmaco.")
22
Pseudomonas aeruginosa: struttura e
fattori di virulenza Colonie su agar

23
Colonization Chronic infection Acute infection
break in front line defenses adhesion thanks to cell-associated virulence factors Chronic infection in cystic fibrosis patients selection of alginate-producing mitants protects from host defense mechanisms low production of extracellular virulence factors tissue damage due to chronic inflammation Acute infection Cell-to-cell signaling systems allow coordinate cell density-dependent production of high amounts of extracellular virulence factors tissue damage by protease and…, leading to blood vessel invasion, diseemination, systemic inflammatory response syndrome, multiple organ failure and death

25
Meccanismi di resistenza agli antibiotici -lattamici
Diminuita espressione di porine (batteri Gram -) Espressione di pompe in grado di determinare l’efflusso del farmaco Alterazioni quali/quantitative dellepenicillin-binding proteins (PBPs) Distruzione enzimatica del farmaco da parte delle -lattamasi
 Espressione di pompe in grado di determinare l’efflusso del farmaco. Alterazioni quali/quantitative dellepenicillin-binding proteins (PBPs) Distruzione enzimatica del farmaco da parte delle -lattamasi.")
26
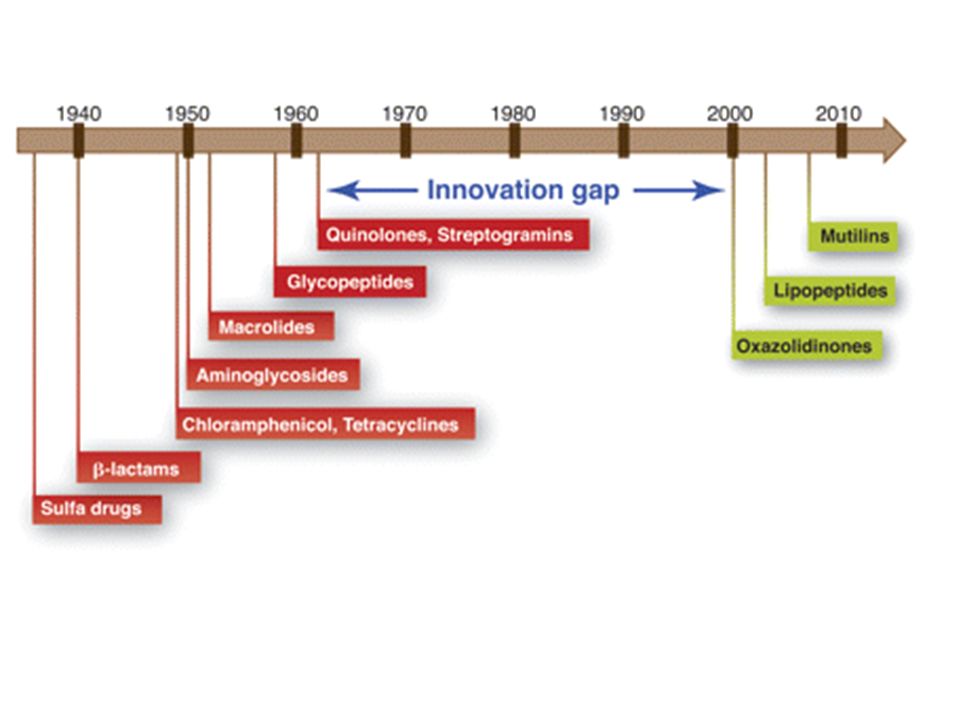
STRUTTURA DEGLI ANTIBIOTICI -LATTAMICI

27
CLASSIFICAZIONE DELLE PENICILLINE: 2 -PENICILLINE ANTI-STAFILOCOCCICHE
(resistenti alla -lattamasi)
")
28
CLASSIFICAZIONE DELLE PENICILLINE: 3 - PENICILLINE AD AMPIO SPETTRO

29
CLASSIFICAZIONE DELLE PENICILLINE: 4 - PENICILLINE A SPETTRO ESTESO
(attive contro Pseudomonas)
")
30
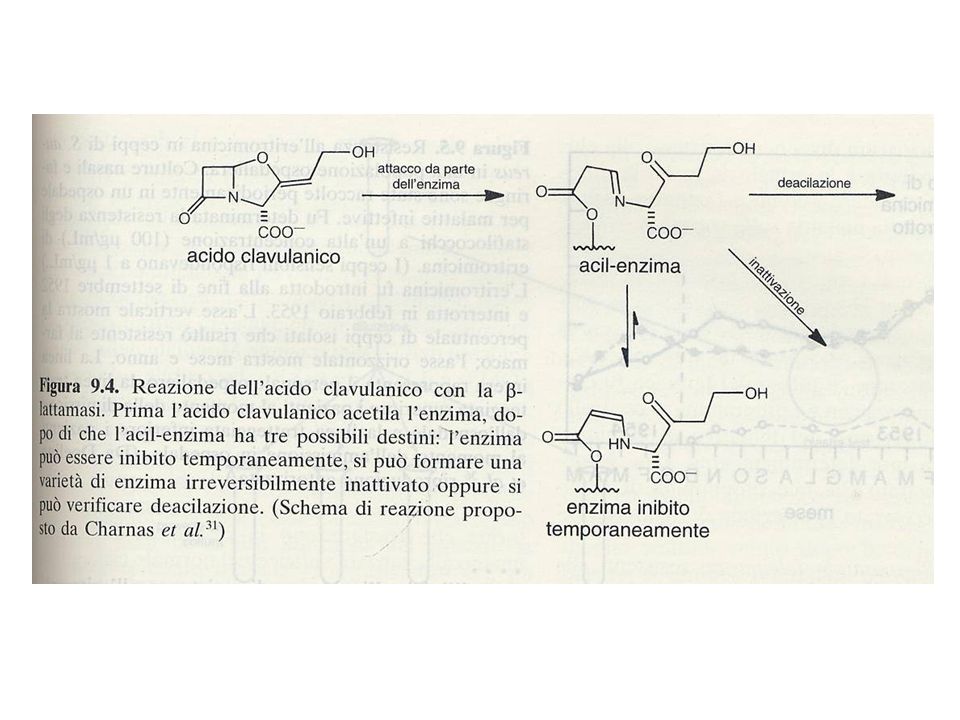
Inibitori delle -lattamasi

32
CEFALOSPORINE COOH ANELLO -LATTAMICO ANELLO DIIDROTIAZINICO
ACIDO 7-AMINOCEFALOSPORANICO

33
CEFALOSPORINE I GENERAZIONE Attività vs Resistenza alle Gram + Gram -
-lattamasi I GENERAZIONE (es. cefalotina, cefazolina, cefalexina) II GENERAZIONE (es. cefamandolo, cefoxitina, cefuroxima) III GENERAZIONE (es. cefotaxime, ceftazidime, cefoperazone) IV GENERAZIONE (es. cefepime, cefpirome)
 II GENERAZIONE. (es. cefamandolo, cefoxitina, cefuroxima) III GENERAZIONE. (es. cefotaxime, ceftazidime, cefoperazone) IV GENERAZIONE. (es. cefepime, cefpirome)")
34
ANTIBIOTICI MONOBATTAMICI
SO3H O

35
ANTIBIOTICI CARBAPENEMICI

36
IMIPENEM DEGRADAZIONE DIPEPTIDASI CILASTATINA

37
Classificazione delle -lattamasi
Classe A: comprende la maggior parte delle forme di rilevanza clinica, generalmente codificate da plasmidi e sensibili agli inibitori attualmente in commercio; contengono un residuo di serina a livello del sito attivo Classe B: sono metalloenzimi, contenenti un atomo di Zn a livello del sito attivo e sono quelle con la più ampia specificità di substrato; sono insensibili agli inibitori attualmente in commercio Classe C: sono generalmente codificate da DNA cromosomiale e attive nei confronti delle cefalosporine, che sono in grado di indurne l’espressione attraverso derepressione genica; contengono un residuo di serina a livello del sito attivo Classe D: sono attive principalmente nei confronti delle oxacilline; contengono un residuo di serina a livello del sito attivo

38
Spettro d’azione delle -lattamasi

39
REAZIONI INDESIDERATE AI FARMACI (Adverse Drug Reactions, ADR)
LIEVI MODERATE GRAVI LETALI LIEVI: non richiedono un antidoto, una terapia o il prolungamento della degenza ospedaliera; MODERATE: richiedono una modificazione del regime di dosaggio del farmaco, ma non necessariamente la sua sospensione e possono prolungare la degenza ospedaliera o richiedere un trattamento specifico; GRAVI: sono potenzialmente pericolose per la vita, richiedendo la sospensione del farmaco e un trattamento specifico; LETALI: concorrono in modo diretto o indiretto alla morte del paziente

40
REAZIONI INDESIDERATE AI FARMACI CATEGORIE
EFFETTI COLLATERALI TOSSICITÀ DA SOVRADOSAGGIO ALLERGIE IDIOSINCRASIE Gli EFFETTI COLLATERALI possono essere definiti come effetti farmacologici prevedibili dose-dipendenti che si verificano nell'ambito dei dosaggi terapeutici e che sono indesiderabili in un determinato contesto terapeutico. Per esempio, la diarrea è un effetto collaterale di molti antibiotici orali; essa compare perché gli antibiotici distruggono non soltanto i batteri che infettano i tessuti ma anche quelli che costituiscono la flora microbica normale del tratto GI. Qualche volta gli effetti collaterali si rivelano utili. Per esempio la sonnolenza prodotta dagli antiistaminici, che è indesiderabile nel trattamento delle allergie durante le ore diurne, può essere utile al momento di coricarsi se il paziente ha un'insonnia associata. La TOSSICITÀ DA SOVRADOSAGGIO è il tipo di tossicità che si verifica in maniera prevedibile con dosi al di sopra del range terapeutico per un determinato paziente. Essa è un fenomeno in parte sovrapposto al tipo di tossicità degli effetti collaterali, specialmente nel caso di farmaci con indice terapeutico ristretto. La gravità della reazione è solitamente dose-dipendente (p. es., le emorragie da anticoagulanti orali, le convulsioni da anestetici locali). La tossicità da sovradosaggio può essere la conseguenza di una diminuzione della clearance del farmaco in pazienti con compromissione della funzionalità epatica o renale. Le ALLERGIE AI FARMACI non sono dose-dipendenti, sono frequenti e causano una grave tossicità. Esse si sviluppano quando la reattività di un paziente è alterata a causa di un precedente contatto con un farmaco che si comporta da antigene o da allergene. I sintomi e i segni sono determinati dalle interazioni antigene-anticorpo e sono ampiamente indipendenti dalle proprietà farmacologiche del farmaco. Le reazioni allergiche non sono completamente imprevedibili; la raccolta di una storia clinica approfondita e l'esecuzione di test cutanei appropriati possono identificare gli individui a rischio. IDIOSINCRASIA è un termine impreciso che viene usato per designare reazioni avverse inaspettate e peculiari che si verificano in una piccola percentuale di individui in occasione dell'assunzione di un farmaco. Tali reazioni non sono legate alle proprietà farmacologiche conosciute del farmaco e non sono di natura inequivocabilmente allergica. Ne sono esempi l'emolisi acuta da sulfamidici, la neuropatia periferica da isoniazide e l'anemia aplastica da cloramfenicolo. L'idiosincrasia è stata definita come una reattività anomala a un farmaco determinata geneticamente, ma non tutte le reazioni idiosincrasiche riconoscono una causa farmacogenetica. Il termine potrebbe diventare obsoleto man mano che diventano noti i meccanismi specifici responsabili delle ADR. Per esempio, l'emolisi da sulfamidici è legata a un deficit genetico dell'enzima G6PD e la neuropatia periferica da isoniazide è legata a una predisposizione genetica all'acetilazione lenta del farmaco.
. La tossicità da sovradosaggio può essere la conseguenza di una diminuzione della clearance del farmaco in pazienti con compromissione della funzionalità epatica o renale. Le ALLERGIE AI FARMACI non sono dose-dipendenti, sono frequenti e causano una grave tossicità. Esse si sviluppano quando la reattività di un paziente è alterata a causa di un precedente contatto con un farmaco che si comporta da antigene o da allergene. I sintomi e i segni sono determinati dalle interazioni antigene-anticorpo e sono ampiamente indipendenti dalle proprietà farmacologiche del farmaco. Le reazioni allergiche non sono completamente imprevedibili; la raccolta di una storia clinica approfondita e l esecuzione di test cutanei appropriati possono identificare gli individui a rischio. IDIOSINCRASIA è un termine impreciso che viene usato per designare reazioni avverse inaspettate e peculiari che si verificano in una piccola percentuale di individui in occasione dell assunzione di un farmaco. Tali reazioni non sono legate alle proprietà farmacologiche conosciute del farmaco e non sono di natura inequivocabilmente allergica. Ne sono esempi l emolisi acuta da sulfamidici, la neuropatia periferica da isoniazide e l anemia aplastica da cloramfenicolo. L idiosincrasia è stata definita come una reattività anomala a un farmaco determinata geneticamente, ma non tutte le reazioni idiosincrasiche riconoscono una causa farmacogenetica. Il termine potrebbe diventare obsoleto man mano che diventano noti i meccanismi specifici responsabili delle ADR. Per esempio, l emolisi da sulfamidici è legata a un deficit genetico dell enzima G6PD e la neuropatia periferica da isoniazide è legata a una predisposizione genetica all acetilazione lenta del farmaco.")
41
REAZIONI DI IPERSENSIBILITÀ Classificazione di Gell & Coombs
TIPO I: reazioni mediate da IgE TIPO II: reazioni citotossiche TIPO III: reazioni da immunocomplessi TIPO IV: reazioni di ipersensibilità cellulare Le reazioni di tipo I sono reazioni in cui gli antigeni (allergeni) si combinano con anticorpi specifici della classe IgE che si trovano legati a recettori di membrana sulle mast-cellule tissutali e sui basofili ematici. La reazione antigene-anticorpo provoca il rapido rilascio di potenti mediatori vasoattivi e infiammatori, i quali possono essere preformati (p. es. istamina, triptasi) o sintetizzati de novo a partire dai lipidi di membrana (p. es. leucotrieni e prostaglandine). Nel volgere di alcune ore, le mast-cellule e i basofili rilasciano anche citochine proinfiammatorie (p. es. interleuchina 4 e interleuchina 13). Questi mediatori provocano vasodilatazione, aumento della permeabilità capillare, ipersecrezione ghiandolare, contrazione della muscolatura liscia e infiltrazione tissutale da parte di eosinofili e altre cellule infiammatorie. Le reazioni di tipo II sono reazioni citotossiche che avvengono quando un anticorpo reagisce con le componenti antigeniche di una cellula o di elementi tissutali, oppure con un antigene o un aptene che si trovi legato a una cellula o un tessuto. La reazione antigene-anticorpo può attivare alcune cellule citotossiche (cellule T killer o macrofagi) per dare luogo alla citotossicità cellulo-mediata anticorpo-dipendente. Essa comprende solitamente l'attivazione del complemento e può provocare l'adesione opsoninica mediante il rivestimento della cellula con l'anticorpo; la reazione procede con l'attivazione dei componenti del complemento per mezzo del C3 (con conseguente fagocitosi della cellula) o con l'attivazione di tutto il sistema complementare con conseguente citolisi o danno tissutale. Le reazioni di tipo III sono reazioni da immunocomplessi (IC) dovute al deposito a livello dei vasi o dei tessuti di IC antigene-anticorpo solubili circolanti. Gli IC attivano il complemento e innescano così una sequenza di eventi che conduce alla migrazione di cellule polimorfonucleate e al rilascio di enzimi proteolitici lisosomiali e di fattori di permeabilità nei tessuti, producendo in questo modo una reazione infiammatoria acuta. Le conseguenze della formazione di IC dipendono in parte dalla proporzione relativa di antigene e di anticorpo nell'IC. In presenza di un eccesso di anticorpo, gli IC precipitano rapidamente nel punto in cui è localizzato l'antigene (p. es. all'interno delle articolazioni nell'AR) oppure vengono fagocitati dai macrofagi evitando così di produrre danno. In presenza di un lieve eccesso di antigene, gli IC tendono a essere più solubili e possono causare reazioni sistemiche in seguito alla deposizione in diversi tessuti. Le reazioni di tipo IV sono reazioni di ipersensibilità cellulare, cellulo-mediata, ritardata o di tipo tubercolinico prodotte da linfociti T sensibilizzati in seguito al contatto con un antigene specifico. Gli anticorpi circolanti non vi sono implicati, né sono necessari perché si sviluppi il danno tissutale. La trasmissione dell'ipersensibilità ritardata dagli individui sensibilizzati a quelli non sensibilizzati può avvenire attraverso i linfociti del sangue periferico, ma non attraverso il siero. I linfociti T sensibilizzati che sono stati innescati o attivati dal contatto con un antigene specifico possono provocare il danno immunologico mediante un effetto tossico diretto o attraverso la liberazione di sostanze solubili (linfochine). Nelle colture tissutali, dopo la sensibilizzazione i linfociti T attivati distruggono le cellule bersaglio per contatto diretto. Le citochine liberate dai linfociti T attivati comprendono diversi fattori che influenzano l'attività dei macrofagi, dei neutrofili e delle cellule killer linfoidi .
 si combinano con anticorpi specifici della classe IgE che si trovano legati a recettori di membrana sulle mast-cellule tissutali e sui basofili ematici. La reazione antigene-anticorpo provoca il rapido rilascio di potenti mediatori vasoattivi e infiammatori, i quali possono essere preformati (p. es. istamina, triptasi) o sintetizzati de novo a partire dai lipidi di membrana (p. es. leucotrieni e prostaglandine). Nel volgere di alcune ore, le mast-cellule e i basofili rilasciano anche citochine proinfiammatorie (p. es. interleuchina 4 e interleuchina 13). Questi mediatori provocano vasodilatazione, aumento della permeabilità capillare, ipersecrezione ghiandolare, contrazione della muscolatura liscia e infiltrazione tissutale da parte di eosinofili e altre cellule infiammatorie. Le reazioni di tipo II sono reazioni citotossiche che avvengono quando un anticorpo reagisce con le componenti antigeniche di una cellula o di elementi tissutali, oppure con un antigene o un aptene che si trovi legato a una cellula o un tessuto. La reazione antigene-anticorpo può attivare alcune cellule citotossiche (cellule T killer o macrofagi) per dare luogo alla citotossicità cellulo-mediata anticorpo-dipendente. Essa comprende solitamente l attivazione del complemento e può provocare l adesione opsoninica mediante il rivestimento della cellula con l anticorpo; la reazione procede con l attivazione dei componenti del complemento per mezzo del C3 (con conseguente fagocitosi della cellula) o con l attivazione di tutto il sistema complementare con conseguente citolisi o danno tissutale. Le reazioni di tipo III sono reazioni da immunocomplessi (IC) dovute al deposito a livello dei vasi o dei tessuti di IC antigene-anticorpo solubili circolanti. Gli IC attivano il complemento e innescano così una sequenza di eventi che conduce alla migrazione di cellule polimorfonucleate e al rilascio di enzimi proteolitici lisosomiali e di fattori di permeabilità nei tessuti, producendo in questo modo una reazione infiammatoria acuta. Le conseguenze della formazione di IC dipendono in parte dalla proporzione relativa di antigene e di anticorpo nell IC. In presenza di un eccesso di anticorpo, gli IC precipitano rapidamente nel punto in cui è localizzato l antigene (p. es. all interno delle articolazioni nell AR) oppure vengono fagocitati dai macrofagi evitando così di produrre danno. In presenza di un lieve eccesso di antigene, gli IC tendono a essere più solubili e possono causare reazioni sistemiche in seguito alla deposizione in diversi tessuti. Le reazioni di tipo IV sono reazioni di ipersensibilità cellulare, cellulo-mediata, ritardata o di tipo tubercolinico prodotte da linfociti T sensibilizzati in seguito al contatto con un antigene specifico. Gli anticorpi circolanti non vi sono implicati, né sono necessari perché si sviluppi il danno tissutale. La trasmissione dell ipersensibilità ritardata dagli individui sensibilizzati a quelli non sensibilizzati può avvenire attraverso i linfociti del sangue periferico, ma non attraverso il siero. I linfociti T sensibilizzati che sono stati innescati o attivati dal contatto con un antigene specifico possono provocare il danno immunologico mediante un effetto tossico diretto o attraverso la liberazione di sostanze solubili (linfochine). Nelle colture tissutali, dopo la sensibilizzazione i linfociti T attivati distruggono le cellule bersaglio per contatto diretto. Le citochine liberate dai linfociti T attivati comprendono diversi fattori che influenzano l attività dei macrofagi, dei neutrofili e delle cellule killer linfoidi .")
42
Beta-lactam toxicity. UPPER PANEL: Beta-lactams can modify amino groups on human proteins, creating an immunogenic beta-lactam hapten. LOWER PANEL: in the absence of modification, human proteins are nonantigenic. Modification of endogenous proteins by the addition of a beta-lactam antibiotic results in the formation of a new antigenic determinant that can be recognized as “foreign” by antibodies of the host immune system La reazione immunologica specifica è stata definita soltanto per la benzilpenicillina. Questo farmaco non si lega con i tessuti o le proteine sieriche in modo sufficientemente energico da formare un complesso antigenico, ma il suo principale prodotto di degradazione, l'acido benzilpenicillanico, può combinarsi con le proteine tissutali per formare il benzilpenicilloile (BPO), il determinante antigenico principale della penicillina. Diversi determinanti antigenici minori vengono formati in quantitativi relativamente piccoli con meccanismi meno ben definiti. Le reazioni di ipersensibilità (di tipo I, II, III, IV) il più delle volte coinvolgono il determinante BPO. Anticorpi IgE diretti contro i determinanti minori possono essere responsabili, in alcuni pazienti, di anafilassi e orticaria. Sono stati individuati anticorpi IgG contro il determinante principale, ma non contro i determinati minori. Essi possono agire come "anticorpi bloccanti" per il BPO, modificando o anche impedendo una reazione contro il BPO stesso, mentre la mancanza di anticorpi IgG bloccanti diretti contro i determinanti minori può spiegare la capacità di questi determinanti di provocare anafilassi. Tutte le penicilline semisintetiche (p. es. l'amoxicillina, la carbenicillina, la ticarcillina) potenzialmente reagiscono in maniera crociata con la penicillina, cosicché i pazienti ipersensibili alla penicillina spesso reagiscono anche contro di esse. Reazioni crociate si verificano in grado minore con le cefalosporine. La terapia con una cefalosporina va cominciata con grande cautela se il paziente ha una storia di reazioni gravi (p. es. anafilassi) alla penicillina. L'anemia emolitica indotta dalla penicillina è una reazione ematologica da anticorpi (citotossica, di tipo II) : l'anticorpo reagisce con l'aptene, che è saldamente legato alla membrana dei GR, provocando agglutinazione e aumento della distruzione dei GR stessi.
, il determinante antigenico principale della penicillina. Diversi determinanti antigenici minori vengono formati in quantitativi relativamente piccoli con meccanismi meno ben definiti. Le reazioni di ipersensibilità (di tipo I, II, III, IV) il più delle volte coinvolgono il determinante BPO. Anticorpi IgE diretti contro i determinanti minori possono essere responsabili, in alcuni pazienti, di anafilassi e orticaria. Sono stati individuati anticorpi IgG contro il determinante principale, ma non contro i determinati minori. Essi possono agire come anticorpi bloccanti per il BPO, modificando o anche impedendo una reazione contro il BPO stesso, mentre la mancanza di anticorpi IgG bloccanti diretti contro i determinanti minori può spiegare la capacità di questi determinanti di provocare anafilassi. Tutte le penicilline semisintetiche (p. es. l amoxicillina, la carbenicillina, la ticarcillina) potenzialmente reagiscono in maniera crociata con la penicillina, cosicché i pazienti ipersensibili alla penicillina spesso reagiscono anche contro di esse. Reazioni crociate si verificano in grado minore con le cefalosporine. La terapia con una cefalosporina va cominciata con grande cautela se il paziente ha una storia di reazioni gravi (p. es. anafilassi) alla penicillina. L anemia emolitica indotta dalla penicillina è una reazione ematologica da anticorpi (citotossica, di tipo II) : l anticorpo reagisce con l aptene, che è saldamente legato alla membrana dei GR, provocando agglutinazione e aumento della distruzione dei GR stessi.")
43
ANTIBIOTICI PEPTIDICI
POLIMIXINA B The polymyxins (Polymixin B and colistin) are basic peptides that act as cationic detergents. Their antimicrobial activity is restricted to Gram-negative bacteria. They are surface active, amphipathic agents that interact strongly with phospholipids and penetrate into and disrupt the structure of cell membranes.
 are basic peptides that act as cationic detergents. Their antimicrobial activity is restricted to Gram-negative bacteria. They are surface active, amphipathic agents that interact strongly with phospholipids and penetrate into and disrupt the structure of cell membranes.")
44
LIPOPEPTIDI CICLICI Daptomycin is an antibacterial agent of a new class of antibiotics, the cyclic lipopeptides. Daptomycin is a natural product which has clinical utility in the treatment of infections caused by aerobic Gram-positive bacteria. The mechanism of action of daptomycin is distinct from any other antibiotic. Daptomycin binds to bacterial membranes and causes a rapid depolarization of membrane potential. The loss of membrane potential leads to inhibition of protein, DNA and RNA synthesis, which results in bacterial cell death. FDA Drug Approval List (cited 29 October 2003)
")
Presentazioni simili













 Regione catalitica (omologa a GyrA) Regione C-terminale (variabile)>")
>")


 PRRs (Patterns Recognition Receptors) Fc Receptors C Receptors IgG.>")

