Scaricare la presentazione
1
Da Cosa Dipende il II Livello Decisionale?
Il II Livello Decisionale deve discriminare le proteine integrali di membrana da quelle che saranno invece liberate nel lume del reticolo. Anche questo livello di decisione è fondato su una breve sequenza amminoacidica posta in una regione della catena, ma a valle del peptide segnale, chiamata SEQUENZA DI ARRESTO DEL TRASFERIMENTO.

2
Sequenza di Arresto del Trasferimento
Essa è composta da: Amminoacidi per la maggior parte idrofobi, quindi adatti a rimanere inseriti nello spessore del foglietto lipidico della membrana del reticolo endoplasmatico rugoso; A questo gruppo di amminoacidi segue un numero di amminoacidi a carica positiva; la concentrazione di tali cariche determina un ostacolo fisico-chimico insormontabile al passaggio della catena polipeptidica attraverso la membrana.

3
Il segmento di catena polipeptidica compreso tra l’estremo N-terminale e la sequenza di arresto si trova entro il reticolo; La sequenza d’arresto è integrata nella membrana ed il tratto rimanente in direzione C-terminale sporge dalla superficie citoplasmatica del reticolo rimanendo a contatto del citoplasma.

4
Da notare che: Il canale di traslocazione, se da una parte mantiene una via di comunicazione che attraversa totalmente la membrana, dall’altra provvede ad aprirsi allo scopo di poter scaricare le proteine di nuova sintesi.

5
Traslocazione post-Traduzionale
Questo processo permette ad alcune proteine, sintetizzate da ribosomi liberi e rilasciate nel citosol, di essere trasferite ad altri organelli come per esempio ai mitocondri che peraltro ricevono attraverso questa via una parte considerevole delle loro proteine funzionali.

6
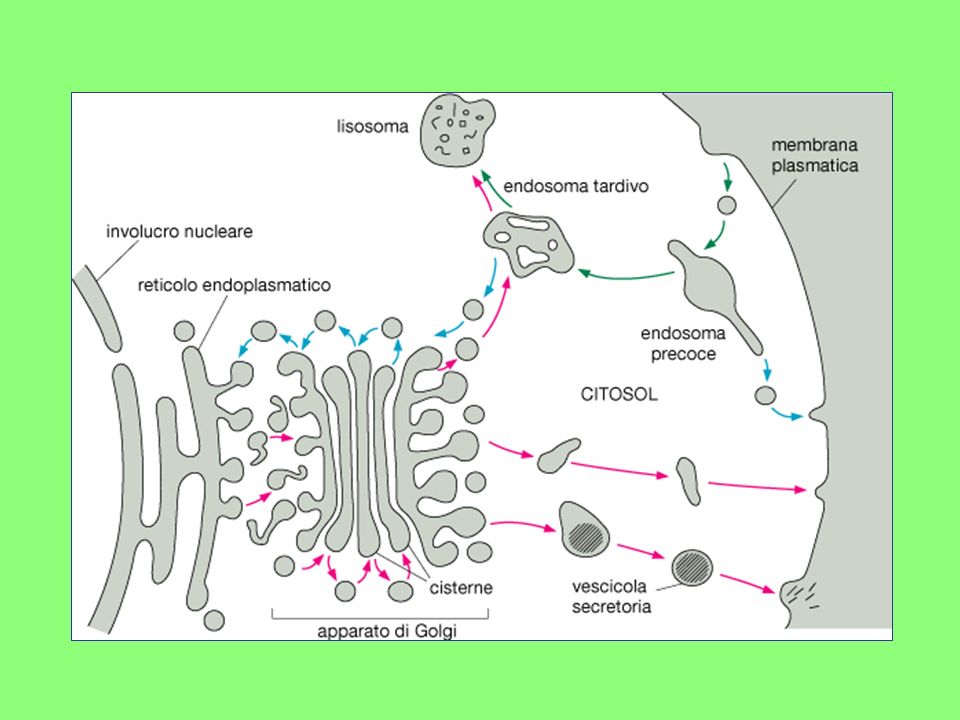
TRASPORTO VESCICOLARE
E’ un processo molto importante che ha il compito di mettere in comunicazione l’interno della cellula con l’esterno ed è sostenuto dalle vescicole di trasporto. Il trasporto vescicolare è reso possibile da un continuo processo di gemmazione cui segue un processo di fusione delle stesse vescicole di trasporto.

7
Traffico Vescicolare: la Via Secretoria
Tale via è diretta verso l’esterno ed i percorsi sono i seguenti: Biosintesi e traslocazione proteica nella membrana del RE; Dall’RE all’apparato del Golgi; Dal Golgi alla membrana plasmatica oppure ai lisosomi, attraverso l’endosoma.

8
Traffico Vescicolare: la Via Endocitica
Tale via è diretta verso l’interno della cellula e le molecole extracellulari vengono inglobate in vescicole derivate dalla membrana plasmatica e trasportate al lisosoma attraverso l’endosoma tardivo.

9
Traffico Vescicolare Affinchè il trasporto vescicolare sia operato correttamente, è necessario che ogni vescicola di trasporto porti con sé solamente le proteine necessarie al destinatario e si fondi in maniera appropriata con la membrana bersaglio. Questo è il motivo per il quale i diversi tipi di vescicole di trasporto sono caratterizzati da un corredo molecolare specifico.

10
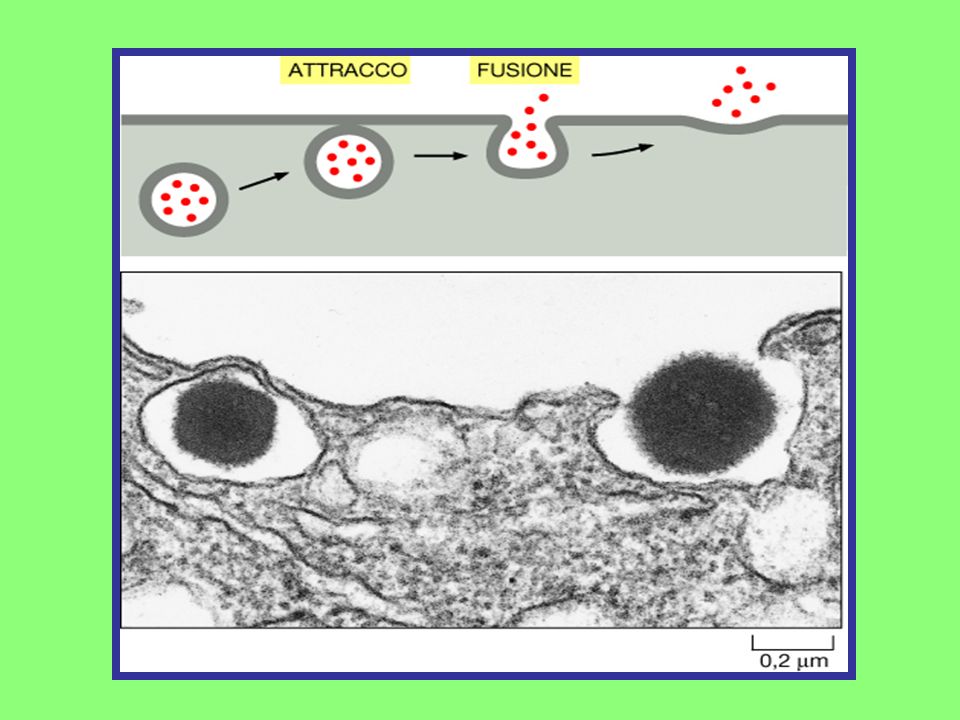
Gemmazione Vescicolare
Le vescicole che gemmano dalla membrana plasmatica mostrano un tipico rivestimento proteico sulla faccia rivolta verso il citosol; Il rivestimento da un lato ha la funzione di conferire la conformazione a gemma e dall’altro permette l’inglobamento delle molecole da trasportare; Il rivestimento proteico meglio conosciuto è rappresentato dalla CLATRINA; Le vescicole rivestite di clatrina possono gemmare dall’apparato del Golgi, via secretoria, oppure dalla membrana plasmatica nella via endocitica. Gemmazione Vescicolare

11
Gemmazione Vescicolare
Immagine al microscopio elettronico che mostra le vescicole che gemmano sulla membrana plasmatica di cellule epiteiali poste in coltura. Gemmazione Vescicolare

12
Assemblaggio di una Vescicola Secretoria Rivestita da Clatrina

13
I recettori di carico hanno il compito di riconoscere e legare le molecole da trasportare;
Le adaptine hanno il compito di legare da un lato i recettori di carico e dall’altro le molecole di clatrina; Le molecole di clatrina si aggregano in una rete a forma di canestro sulla faccia citosolica della membrana stabilizzando così l’ associazione molecolare; Le molecole di dinamina sono capaci di legare GTP e si aggregano in un anello intorno alla radice di ogni fossa nel momento in cui è profondamente invaginata.

14
Clatrina

15
Da notare che: Questo processo è comune alle vescicole di trasporto a carico del reticolo, dell’apparato del Golgi nonchè della membrana plasmatica; Esistono diversi tipi di adaptine ed in particolare quelle che legano i recettori di carico al livello della membrana plasmatica sono diversi da quelli che legano i recettori di carico al livello dell’apparato del Golgi; Nel momento in cui la dinamina idrolizza il GTP, si assiste alla contrazione dell’anello con conseguente distacco dalla membrana; A gemmazione completa le proteine di rivestimento si staccano e la restante parte della vescicola si fonde con la membrana bersaglio.

16
Esistono diversi tipi di proteine di rivestimento dedicate al trasporto vescicolare.

17
La Specificità delle Vescicole
Le vescicole di trasporto devono riconoscere specificamente l’organello bersaglio affinchè avvenga la fusione e la liberazione del contenuto; Riconosciamo pertanto le proteine transmembrana v-SNARE, presenti sulle vescicole, che vengono specificamente riconosciute da sequenze t-SNARE situate al livello della membrana target; La fusione delle membrane può non seguire immediatamente il riconoscimento sostenuto dagli SNARE. La Specificità delle Vescicole

18
La Specificità delle Vescicole

19
La Fusione delle Vescicole
La fusione richiede una distanza fra le due membrane non maggiore di 1,5 nm; Affinchè si stabilisca un rapporto così stretto, l’acqua deve essere necessariamente esclusa dalla superficie idrofilica della membrana, processo energeticamente sfavorito; E’ stata ipotizzata, pertanto, la presenza di proteine specifiche che, associandosi al sito di fusione, formano un complesso che consente il superamento di questa barriera energetica. La Fusione delle Vescicole

20
La maggior parte delle proteine subiscono modificazioni covalenti al livello del reticolo endoplasmatico: Formazione di ponti disolfuro, per ossidazione a coppie di catene laterali di cisteine, che si realizzano nel lume del RE; Glicosilazione a carico delle proteine che entrano nel lume del RE o nella sua membrana ad opera di enzimi glicosilanti. PERCORSO SECRETORIO

21
La glicosilazione avviene quasi subito dopo l’ingresso del peptide nel lume dell’RE grazie all’aggiunta in blocco di catene laterali oligosaccaridiche a particolari Asparagine (asparagina-X-serina oppure treonina). Le sequenze oligosaccaridiche sono fissate al dolicolo, un lipide specializzato della membrana del RE.

22
Il legame si stabilisce in una sola reazione catalizzata da un enzima legato alla membrana, una protein olisaccaride transferasi, e l’asparagina facente parte della sequenza che determina appunto il sito della N-glicosilazione.

23
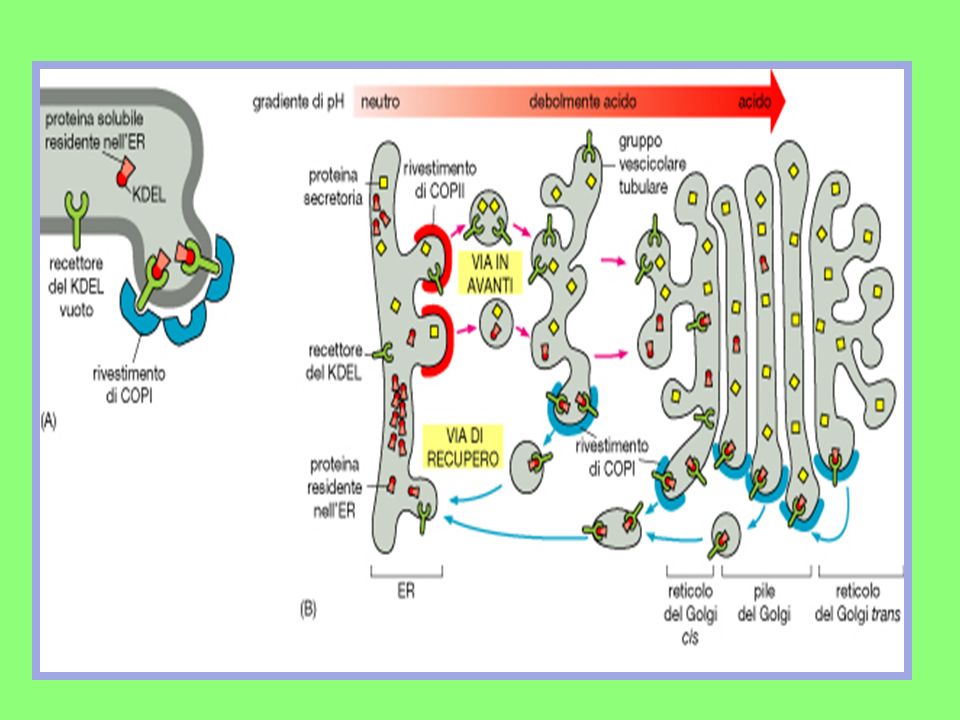
E’ da ricordare che l’uscita dal RE è ben controllato:
Le proteine destinate a funzionare nel RE, sono trattenute da una breve sequenza (4aa) detta segnale di ritenzione in RE, riconosciuta da uno specifico recettore presente sulla membrana del RE stesso; Le proteine destinate al transito in altre sedi, devono superare una vera e propria selezione. SELEZIONE Essa è sostenuta dall’azione di proteine secondatrici residenti (chaperon) che hanno il compito di trattenere le proteine nel RE finchè non assumono la conformazione più idonea al transito; se ciò non accade, le proteine vengono degradate.
 detta segnale di ritenzione in RE, riconosciuta da uno specifico recettore presente sulla membrana del RE stesso; Le proteine destinate al transito in altre sedi, devono superare una vera e propria selezione. SELEZIONE. Essa è sostenuta dall’azione di proteine secondatrici residenti (chaperon) che hanno il compito di trattenere le proteine nel RE finchè non assumono la conformazione più idonea al transito; se ciò non accade, le proteine vengono degradate.")
25
L’apparato di Golgi

26
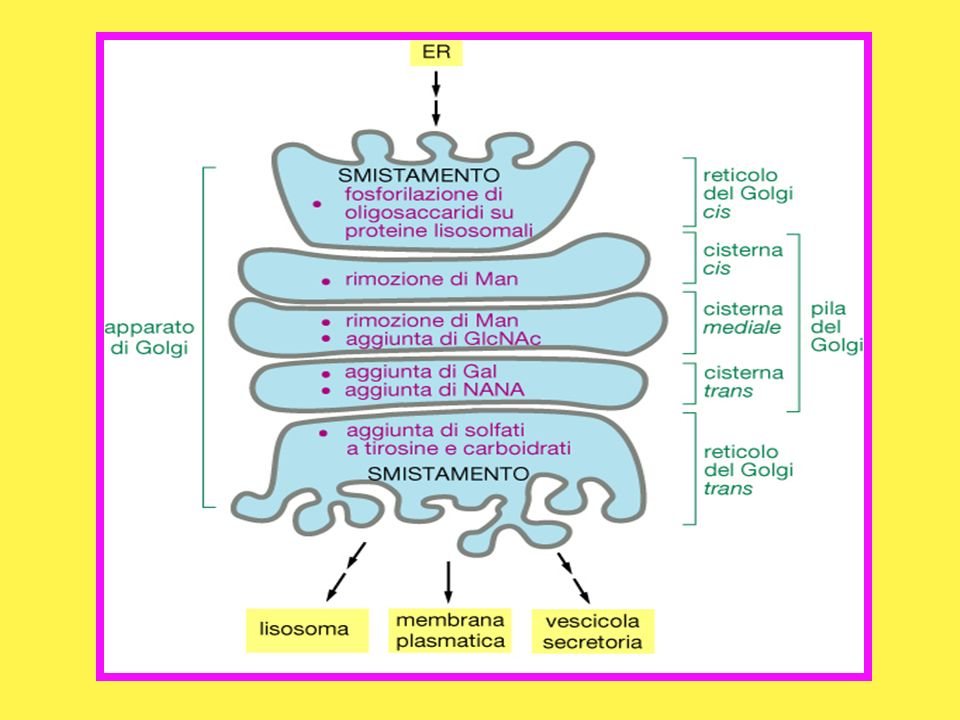
E’ situato vicino al nucleo e consiste di una serie di cisterne appiattite, delimitate da membrane e disposte le une sulle altre ed il loro numero varia a seconda del tipo cellulare in esame; Distinguiamo un lato di ingresso, o cis, ed uno di uscita, ovvero in trans; Nel Golgi cis arrivano le proteine provenienti dal RE via vescicole di trasporto; Attraverso il Golgi trans, invece, transitano le proteine destinate alla superficie cellulare o ad altri comparti cellulari, opportunamente veicolate da vescicole di trasporto.

29
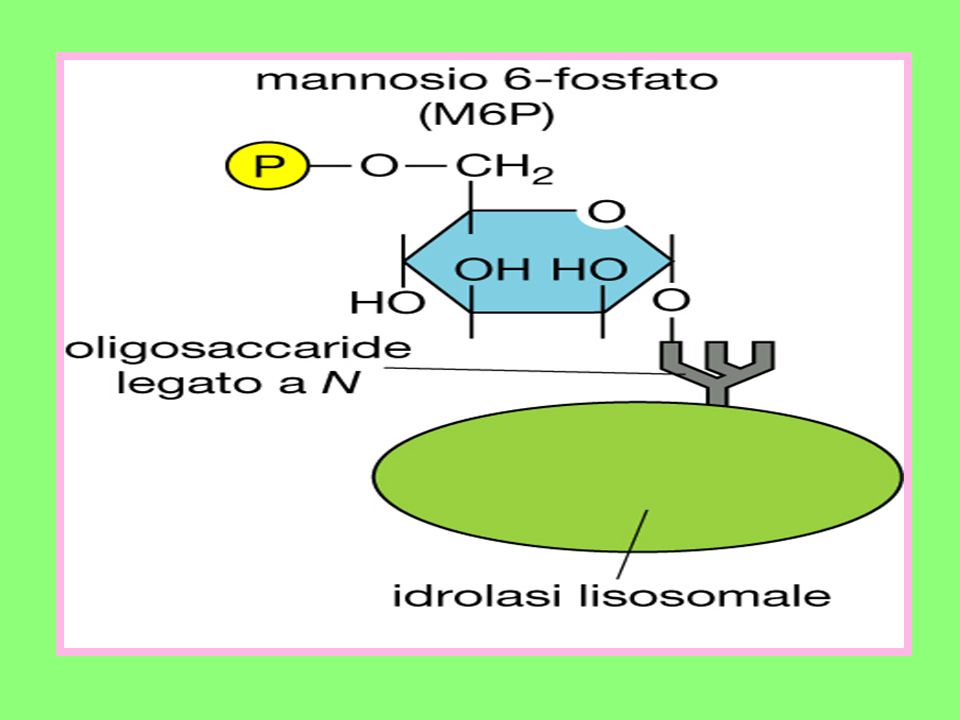
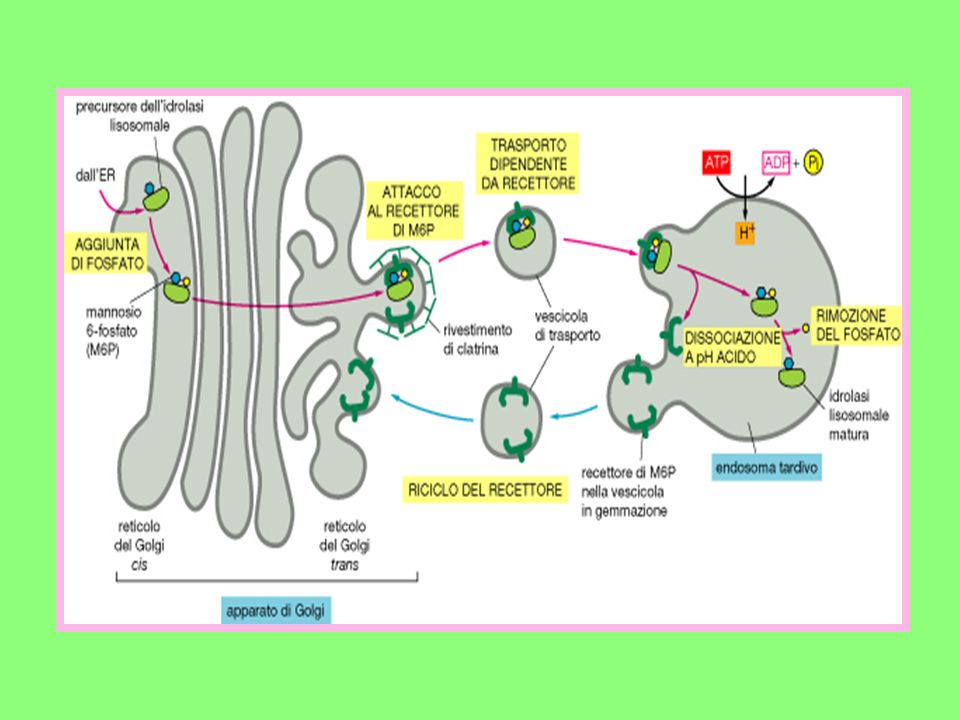
All’Apparato del Golgi giungono:
Componenti di membrana prodotti nel RE; Una miscela di proteine non ancora completamente glicosilate. Nell’Apparato del Golgi deve avvenire: La rifinitura di tutti i prodotti che hanno caratteristiche strutturali non ancora definitive; Il loro avviamento verso la destinazione che a ciascuno compete. Un esempio ci è offerto da alcuni enzimi lisosomiali le cui catene oligosaccaridiche una volta giunte in cis, vengono fosforilate in corrispondenza di due residui di mannosio che sono trasformati in mannosio 6-fosfato.

32
Rielaborazione dell’Oligosaccaride nell’ER e nell’Apparato di Golgi
Generazione di Catene Oligosaccaridiche Complesse

33
Nell’RE, l’elaborazione inizia con la rimozione dei glucosi dall’oligosaccaride trasferito iniziamente alla proteina; Una mannosidasi della membrana dell’RE rimuove un mannosio specifico. Nel Golgi, la mannosidasi I rimuove altri 3 mannosi e la N-acetil glucosammina transferasi I aggiunge N-acetilglucosammina che permette alla Mannosidasi II di rimuovere altri 2 mannosi così che possano essere aggiunte altre 2 molecole di N-acetilglucosamina; La successiva aggiunta di tre molecole di galattosio e tre di acido sialico completa l’oligosaccaride complesso.

34
Qual è lo scopo della glicosilazione?
Poiché le catene degli zuccheri hanno una flessibilità limitata, anche un piccolo oligosaccaride è in grado di sporgere da una glicoproteina limitando così l’avvicinamento di altre macromolecole. In questo modo, per esempio, la presenza di oligosaccaridi tende a rendere una glicoproteina relativamente resistente alla digestione proteolitica. E’ verosimile che gli oligosaccaridi sulle proteine di superficie della cellula fornissero in origine alla cellula eucariotica un rivestimento protettivo che, a differenza della parete cellulare rigida dei batteri, permetteva alla cellula di cambiare forma e di muoversi.Nel corso dell’evoluzione, tali glicoproteine come per esempio le selettine si sono “convertite” a svolgere funzioni di riconoscimento durante il fenomeno dell’adesione cellulare.

35
Si ritiene che il trasferimento ordinato delle proteine da un compartimento a quello immediatamente successivo si compia per mezzo di vescicole di trasporto che si liberano per gemmazione dalle cisterne di ciascun compartimento per fondersi con quelle del compartimento successivo riversandovi il contenuto. Finalmente dall’ ultimo compartimento si liberano vescicole dirette verso destinazioni finali diverse. Infatti, tra le proteine che attraversano l’Apparato del Golgi si possono identificare: Proteine che costituiscono gli enzimi dei lisosomi; Proteine destinate ad essere secrete fuori dalla cellula; Proteine destinate ad essere inserite nella membrana cellulare.

37
Esistono due forme di secrezione: Secrezione Costitutiva;
Secrezione Regolata. Nella Secrezione Costitutiva le proteine costituenti i prodotti di secrezione vengono immediatamente trasferite all’esterno attraverso le vescicole di secrezione che vanno a fondersi con la membrana plasmatica durante il ciclo di produzione. Nella Secrezione Regolata le proteine vengono conservate nelle vescicole di secrezione che la cellula può accumulare finchè un appropriato stimolo non ne induce la secrezione.

38
Forme di Secrezione La Secrezione Costitutiva è continua e fornisce, per esempio, proteine di nuova sintesi alla membrana plasmatica nel caso in cui questa deve crescere di dimensioni per poi dividersi. La Secrezione Regolata è intermittente e le vescicole si accumulano in prossimità della membrana, ma si fondono con essa solo quando la cellula viene stimolata da un segnale extracellulare rappresentato ad esempio da un aumento del livello ematico di glucosio che segnala ad alcune cellule pancreatiche di secernere l’ormone insulina.

39
Le cellule eucariotiche sono in grado di assumere fluidi, molecole piccole e molecole grandi attraverso il processo di endocitosi. Riconosciamo: La Fagocitosi ovvero l’ingestione di solidi e La Pinocitosi ovvero l’ingestione dei liquidi. ENDOCITOSI

40
Negli organismi animali esistono cellule specializzate, chiamate fagociti o macrofagi che esercitano la fagocitosi. Sono cellule di origine ematica e sono in grado di distruggere batteri, virus, cellule morte o gravemente danneggiate. FAGOCITOSI Essa si divide in due fasi consecutive rappresentate dal riconoscimento della particella da ingerire ed il suo internamento.

41
Le particelle virali o batteriche non sono direttamente aggredibili dalle cellule fagocitarie perché sprovvisti di specifici ligandi affini ai loro recettori. In questo caso, le cellule immunocompetenti dell’organismo possono produrre anticorpi diretti contro i determinanti antigenici presenti sulla superficie della particella virale o batterica così che i macrofagi possano riconoscere e legare con i loro recettori gli anticorpi adesi alla superficie della particella stessa. Riconoscimento

42
L’internamento delle particelle segue il riconoscimento e porta al loro trasferimento nel corpo cellulare, entro un involucro di membrana che lo separa dal citoplasma chiamato fagosoma e ad esso si fondono i lisosomi. Nei lisosomi secondari, quindi, il materiale fagocitato può essere digerito, ma i prodotti della digestione tornano nel citoplasma ed i residui inutilizzabili vengono successivamente espulsi. Internamento

43
La pinocitosi consiste nell’introduzione nel corpo cellulare di minuscole goccioline di liquido prelevate dall’ambiente extracellulare attraverso la formazione di vescicole pinocitotiche. Distinguiamo la pinocitosi aspecifica e la pinocitosi selettiva. PINOCITOSI

44
PINOCITOSI ASPECIFICA
La pinocitosi aspecifica provvede all’assunzione di liquidi extracellulari attraverso la formazione di vescicole, ma non si assiste a nessuna modificazione della composizione. In questo modo la cellula introduce i materiali presenti nel liquido extracellulare in quantità proporzionali alla loro concentrazione. PINOCITOSI ASPECIFICA

45
Nella pinocitosi selettiva o endocitosi mediata da recettori, la membrana che costituisce la vescicola è fornita di speciali recettori con i quali raccoglie dall’ambiente esterno speciali tipi di molecole o macromolecole ed introduce quindi una gocciolina che non ha più la composizione originale del liquido extracellulare perché è stata arricchita dei composti di maggiore interesse per l’economia cellulare. PINOCITOSI SELETTIVA

46
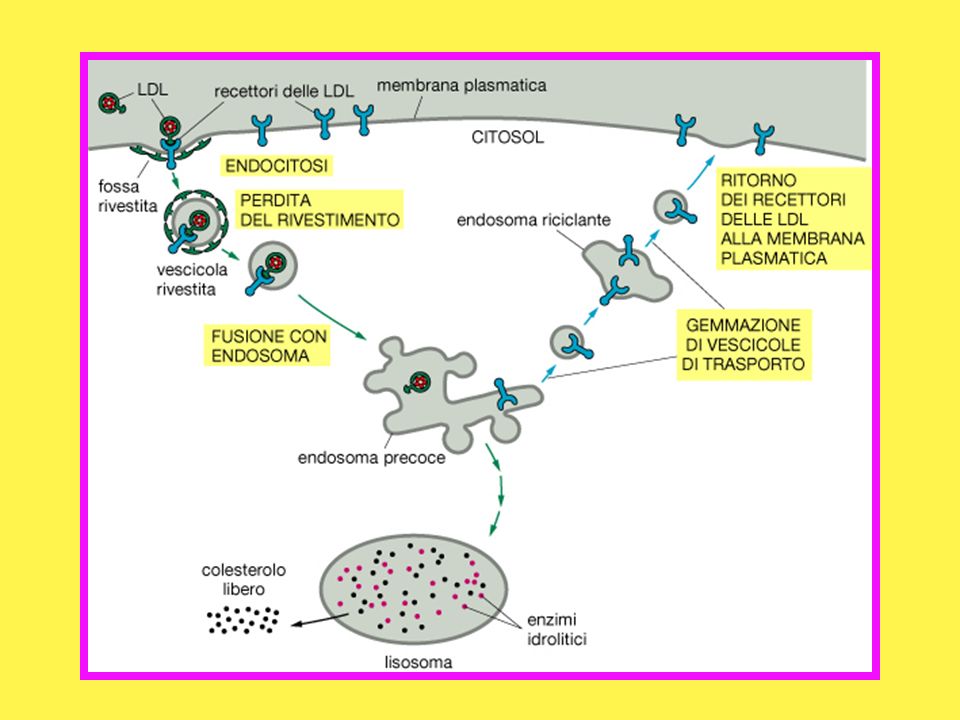
Il colesterolo è insolubile in acqua e viaggia nel circolo sanguigno legato a lipoproteine a bassa densità o LDL; Le LDL si legano ai recettori presenti sulla superficie della cellula; Si formano così le vescicole rivestite di clatrina; Le vescicole perdono il loro rivestimento; Si assiste alla fusione con gli endosomi in cui le LDL vengono dissociate dai loro recettori.

47
Gli ENDOSOMI sono corpi endocellulari costituiti dalla confluenza di vescicole di endocitosi. Sono di norma implicati nell’accumulo di metaboliti assunti selettivamente dalla cellula dall’ambiente extracellulare con il meccanismo della endocitosi mediata da recettori.

49
ORGANIZZAZIONE della FORMA e dei MOVIMENTI
La morfologia e le funzioni di una cellula sono strettamente interdipendenti, nel senso che lo svolgimento delle attività di ogni singola cellula richiede da un lato il mantenimento dell’organizzazione interna e dall’altro quello della forma. ORGANIZZAZIONE della FORMA e dei MOVIMENTI ESEMPI: La conformazione allungata della cellula muscolare rende più efficace la loro attività contrattile; La presenza di lunghi e sottili prolungamenti è essenziale per la funzione di conduzione delle cellule nervose; La presenza dei microvilli sulla superficie delle cellule epiteliali intestinali è una caratteristica importante per la loro attività di assorbimento selettivo di molecole.

50
Il citoscheletro contribuisce:
A conferire la forma; Ad organizzare una disposizione ordinata degli organelli; A determinare sia i movimenti interni richiesti dalle attività cellulari sia gli eventuali spostamenti della cellula nell’ambiente extracellulare. Il Citoscheletro

51
Il Citoscheletro Cellula della pelle (fibroblasto) fissata e colarata con Blue di Coomassie.
 fissata e colarata con Blue di Coomassie.")
52
Le conoscenze attuale è la risultante di studi ultrastrutturali, immunoistochimici e biochimici i cui risultati devono ovviamente essere integrati ed interpretati globalmente affinchè si abbia un quadro generale delle sue proprietà. Il Citoscheletro

53
Il Citoscheletro Sistemi di Fibre Diametro Medio
Componente proteico principale Caratteristiche funzionali Microtubuli 25 nm Tubulina Strutture Dinamiche Microfilamenti 7 nm Actina Filamenti Intermedi 10 nm Variabile Strutture Statiche

54
Il Citoscheletro: Il Sistema dei Microtubuli
Il sistema dei microtubuli è prodotto dall’organizzazione ordinata di 2 tipi di subunità proteiche globulari chiamate Tubulina a e Tubulina b che differiscono nella loro composizione amminoacidica. I microtubuli sono generati dalla seguente associazione progressiva: Ogni subunità a si lega ad una subunità b formando un eterodimero a-b, ovvero particella 6S; I dimeri si legano in file ordinate o Protofilamenti composti dai due tipi di subunità regolarmente alternati; I protofilamenti si associano fra loro in gruppi di 13, in modo che ciascun gruppo di associazione costituisce una struttura cilindrica cava.

55
Il Citoscheletro: Il Sistema dei Microtubuli
D: Microtubulo visto lateralmente al microscopio elettronico; E: Sezione trasversale del microtubulo con il suo anello di 13 subunità.

56
A D Il Citoscheletro: Il Sistema dei Microtubuli
Studiando la polimerizzazione in vitro della tubulina si può constatare che i microtubuli che si ottengono sono funzionalmente polarizzati. I microtubuli possono incorporare e perdere subunità a-b aggiungendone di nuove in corrispondenza di una delle estremità che chiameremo A, ovvero Accettrice, mentre la perdita avviene principalmente a carico dell’estremità opposta che chiameremo D, ovvero Donatrice. A D

57
Il Citoscheletro: Il Sistema dei Microtubuli
I microtubuli acquistano un breve segmento radioattivo inizialmente localizzato in corrispondenza dell’estremità A; Il segmento radioattivo si sposta gradualmente verso l’estremità opposta per ulteriore aggiunta di subunità non marcate denunziando così la contemporanea perdita di subunità a carico dell’estremità D. A D

58
Quali sono i fattori che determinano e dirigono la formazione dell’apparato microtubulare e la sua distribuzione nel citoplasma? In vitro è stato dimostrato che ogni dimero a-b lega saldamente una molecola di GTP che idrolizza a GDP poco dopo avere aggiunto una unità al microtubulo in crescita. Le molecole di tubulina associate a GTP si organizzano bene insieme nella parete del microtubulo, mentre quelle associate a GDP hanno una conformazione diversa e stabiliscono legami più labili tra loro.

59
Il Citoscheletro: Il Sistema dei Microtubuli
La fissazione di GTP e la successiva idrolisi è quindi richiesta non tanto per fornire energia al processo di polimerizzazione, bensì per conferire agli eterodimeri di tubulina la conformazione più adatta a legarsi tra loro ed a rimanere quindi associati, com’è dimostrato dal fatto che il GTP può essere sostituito da analoghi non idrolizzabili. Consideriamo il fuso mitotico che guida i cromosomi durante la mitosi: L’esposizione delle cellule in mitosi alla Colchicina, una sostanza che lega saldamente la tubulina libera, impedisce la polimerizzazione dei microtubuli e di conseguenza il fuso mitotico sparisce rapidamente e le cellule si arrestano in mitosi incapaci di ripartire i propri cromosomi in due gruppi; L’esposizione delle cellule al Taxolo conduce, invece, all’impossibilità di perdere le subunità con il risultato che i microtubuli si allungano, ma ne è impedito l’accorciamento.

60
Il Citoscheletro: Il Sistema dei Microtubuli
La distruzione del fuso mitotico o la sua inattivazione conduce le cellule in divisione a morte. Le cellule tumorali che hanno una spiccata attività proliferativa risultano molto sensibili ai farmaci antimitotici che destabilizzano o stabilizzano i tubuli e proprio per questo motivo si usano nella terapia del cancro sostanze derivate dalla colchicina oppure dal taxolo.

61
Il Citoscheletro: Il Sistema dei Microtubuli
Osservando la riorganizzazione dell’apparato microtubulare dopo la sua disaggregazione sperimentale, si osserva che esso, una volta rimosso l’agente disaggregante, tende a ricostituirsi prendendo origine in corrispondenza di una particolare regione chiamata CENTROSOMA ovvero centro di organizzazione dei microtubuli, MTOC (Micro Tubule Organizing Centre).
.")
62
Il Centrosoma Il centrosoma è di solito localizzato in prossimità del nucleo; Esso è costituito da una coppia di centrioli contenuti entro una piccola quantità di materiale scarsamente opaco agli elettroni che si continua in modo indefinito con il citoplasma fondamentale; La struttura dei centrioli è caratteristica ed analoga a quella dei microtubuli; I centrioli si presentano come due piccoli cilindri cavi e la loro parete è composta da 9 triplette di microtubuli condensati, orientate in maniera convergente verso l’asse centrale del cilindro.

63
I Centrioli

64
Il Centrosoma Tra centrosoma e microtubuli esiste una relazione di interdipendenza della quale ancora non è possibile definirne con esattezza le basi molecolari. La relazione è dimostrata dal fatto che il centrosoma appare come la sede degli eventi molecolari necessari ad innescare l’assemblaggio dei microtubuli che si formano e cominciano a crescere nella regione pericentriolare. Questo processo ha luogo sia nelle cellule intercinetiche sia in preparazione della divisione cellulare e cioè nel momento in cui la formazione del fuso mitotico procede da due poli costituiti dai centrioli duplicati e dal rispettivo materiale pericentriolare.

65
Il Centrosoma I microtubuli non hanno però contatto diretto con i centrioli, ma solo con la regione pericentriolare.

66
Il Centrosoma I centrosomi contengono migliaia di strutture ad anello costituite da Tubulina g. A: ogni anello rappresenta un sito di nucleazione per la crescita di un microtubulo; B: i dimeri di a-b tubulina si associano alla tubulina g secondo un orientamento specifico.

67
Il Centrosoma I siti di nucleazione risultano fondamentali nel processo di formazione di un microtubulo. Infatti, la tubulina ab, libera e purificata, polimerizza in vitro spontaneamente solo se è concentrata, ma nella cellula vivente la sua concentrazione è troppo bassa perché si possa avviare la prima fase di assemblaggio in assenza di un sito di nucleazione.

68
Instabilità Dinamica dei Microtubuli
A seguito dell’instabilità dinamica dei microtubuli,il centrosoma è costretto ad avviare continuamente la neoformazione di microtubuli stessi. Il microtubulo può proteggersi dalla dissoluzione nel momento in cui si lega ad una molecola oppure ad una struttura cellulare opportuna. La stabilizzazione selettiva del microtubulo determina la polarità cellulare. Nella cellula l’estremità negativa del microtubulo è protetta dal sito di nucleazione, mentre l’estremità positiva viene stabilizzata da altre proteine; il legame con queste è appunto responsabile della polarizzazione della cellula stessa.

69
Le cellule animali a differenziamento completato risultano polarizzate
Nella cellula nervosa tutti i microtubuli assonici puntano nella stessa direzione con l’estremità positiva rivolta verso il neuroterminale. Grazie a questa organizzazione la cellula nervosa invia vescicole e proteine di secrezione prodotte nel corpo cellulare, ma necessarie al terminale assonico.

70
Le CHINESINE si spostano verso l’estremità positiva di un microtubulo;
Le Proteine Motrici Effettuano Il Trasporto Intracellulare Le proteine motrici che si muovono lungo i microtubuli citoplasmatici, come quelle assoniche, appartengono a due famiglie: Le CHINESINE si spostano verso l’estremità positiva di un microtubulo; Le DINEINE si spostano verso l’estremità negativa e cioè verso il centrosoma.

71
Le Proteine Motrici Effettuano Il Trasporto Intracellulare
Le chinesine e le dineine possiedono due teste globulari che legano ATP, nonché i microtubuli, ed una coda che lega il carico. Le teste globulari hanno attività enzimatica idrolitica per ATP e grazie all’energia liberata in questa reazione vanno incontro ad una serie di cambiamenti conformazionali che le fanno procedere lungo i microtubuli stessi.

72
Le Proteine Motrici Effettuano Il Trasporto Intracellulare
ATP Idrolisi ATP Il passaggio da una conformazione all’altra secondo una precisa sequenza è alimentato dall’idrolisi di ATP legata alla proteina. Basta che una di queste transizioni sia accoppiata all’idrolisi di ATP che l’intero ciclo risulta praticamente irreversibile e ad ogni ripetizione del ciclo la proteina avanza lungo il microtubulo (in questo caso verso destra).
.")
73
I microtubuli, le Proteine Motrici e la Localizzazione degli Organelli
ATP Idrolisi ATP La corretta localizzazione degli organelli delimitati da membrana quali reticolo endoplasmatico ed apparato del Golgi dipende dalla presenza dei microtubuli. Man mano che la cellula si sviluppa, le membrane del reticolo si distendono, dall’involucro nucleare fino alla membrana plasmatica, decorrendo parallele ai microtubuli stessi grazie all’azione delle chinesine (direzione centrifuga). In maniera opposta, le dineine provvedono (in direzione centripeta) alla corretta localizzazione dell’apparato del Golgi al centro della cellula.
. In maniera opposta, le dineine provvedono (in direzione centripeta) alla corretta localizzazione dell’apparato del Golgi al centro della cellula.")
74
Cilia, Flagelli e Microtubuli
Le cilia, propulsori di fluidi, sono strutture piliformi che si protendono da molte cellule eucariotiche, vedi cellule epiteliali del tratto respiratorio; I flagelli sono molto simili alle cilia in relazione alla loro struttura interna, ma molto più lunghi, vedi flagelli di uno spermatozoo; La tipica struttura di cilia e flagelli è di “9+2” microtubuli.

75
I Corpuscoli Basali I corpuscoli basali rappresentano i centri di formazione di cilia e flagelli. A: Fotografia al microscopio elettronico di una sezione trasversale di tre corpi basali; B: Disegno di un corpo basale.

76
I microtubuli sono associati a numerose proteine, ma la più importante è la dineina ciliare cui si deve il moto flessorio del fascio di microtubuli. La dineina ciliare lega con la coda un microtubulo, mentre mediante la testa lega un microtubulo adiacente, generando così una forza che li fa scorrere l’uno sull’altro.

77
Cilia, Flagelli e Microtubuli
Il movimento di un ciglio (B) o di un flagello (A) deriva dal piegamento del suo nucleo chiamato Assonema; esso è composto interamente da microtubuli e dalle proteine a loro associate. Da notare che per quanto riguarda il movimento delle cilia, un veloce colpo di potenza delle fasi 1 e 2 in cui il fluido è spinto sopra la superficie della cellula, è seguito da un colpo lento nelle fasi 3, 4 e 5.
 o di un flagello (A) deriva dal piegamento del suo nucleo chiamato Assonema; esso è composto interamente da microtubuli e dalle proteine a loro associate. Da notare che per quanto riguarda il movimento delle cilia, un veloce colpo di potenza delle fasi 1 e 2 in cui il fluido è spinto sopra la superficie della cellula, è seguito da un colpo lento nelle fasi 3, 4 e 5.")
78
Cilia, Flagelli e Microtubuli
A: Scivolamento delle doppiette di microtubuli con le relative molecole di dineina all’atto dell’isolamento; B: Flessibilità delle stesse doppiette di microtubuli intatti.

79
Il Citoscheletro: I Filamenti Actinici
Al microscopio elettronico appaiono come fili del diametro di 7 nm; Riconosciamo 2 polimeri lineari avvolti l’uno sull’altro a spirale con passo di 37 nm. Il Citoscheletro: I Filamenti Actinici

80
Il Citoscheletro: I Filamenti Actinici
I filamenti actinici si trovano in tutte le cellule eucariotiche e sono essenziali per i movimenti (vedi fagocitosi e mitosi); La proteina principale dei microfilamenti è l’actina presente in forma non polimerizzata mediante la combinazione con Profilina; Una volta dissociata dalla profilina, le molecole di actina libere, ovvero actina G, possono associarsi tra loro formando lunghi filamenti avvolti a 2 a 2 in eliche destrorse, ovvero actina F. Actina Sequestrata Actina G Actina F
; La proteina principale dei microfilamenti è l’actina presente in forma non polimerizzata mediante la combinazione con Profilina; Una volta dissociata dalla profilina, le molecole di actina libere, ovvero actina G, possono associarsi tra loro formando lunghi filamenti avvolti a 2 a 2 in eliche destrorse, ovvero actina F. Actina Sequestrata. Actina G. Actina F.")
81
Il Citoscheletro: I Filamenti Actinici
Ogni monomero libero di actina sequestrata porta saldamente legata una molecola di ATP; L’ATP viene idrolizzato poco dopo l’incorporazione del monomero actinico nel filamento; L’ADP può dissociarsi dal monomero soltanto nel momento in cui il monomero stesso depolimerizza. Actina Sequestrata Actina G Actina F

83
Il Citoscheletro: I Filamenti Actinici
Come abbiamo visto accadere anche per la tubulina, la cui polimerizzazione è GTP-dipendente, non è chiaro se l’ATP serva solo come regolatore allosterico oppure anche come fonte di energia necessaria alla polimerizzazione; Come i microtubuli anche i microfilamenti hanno una chiara polarità funzionale cioè possono allungarsi aggiungendo monomeri preferenzialmente ad una sola delle due estremità e possono accorciarsi perdendo monomeri dall’estremità opposta; La forma libera di Actina G e la forma polimerizzata di Actina F si trovano in uno stato di equilibrio dinamico che può essere rapidamente spostato verso sinistra o verso destra da appositi fattori di controllo; Oltre alla profilina, altre proteine contribuiscono a controllare la lunghezza dei microfilamenti e sono definite come fattore di allungamento, di accorciamento, di frammentazione e di stabilizzazione; Sono riconosciuti anche i fattori di associazione capaci di favorire l’associazione di microfilamenti in fasci o reti come pure i fattori di ancoraggio capaci di stabilire legami di tipo laterale o terminale con le membrane ed altre strutture cellulari.

84
Sistemi Motori Fondati su Microfilamenti
L’esempio più perfezionato e meglio conosciuto si ritrova nelle cellule muscolari che fanno della contrattilità l’aspetto caratterizzante del loro stato differenziato. Tutte le proteine motrici actino-dipendenti appartengono alla famiglia delle MIOSINE. Nelle cellule le sottofamiglie più rappresentate sono quelle della MIOSINA I e II.

85
Sistemi Motori Fondati su Microfilamenti
La coda corta della molecola di miosina I possiede siti di legame per vari componenti cellulari, incluse vescicole e membrane plasmatiche; La miosina II è tipicamente muscolare ed i membri di tale famiglia presentano sempre due teste ATPasiche ed una lunga coda a bastoncello. Le molecole di miosina II si riuniscono in gruppi legandosi per le code a spirale e formando un filamento miosinico in cui le teste si proiettano ai lati.

86
Sistemi Motori Fondati su Microfilamenti
Una serie di teste si lega ai filamenti actinici puntando in una direzione e li tira da una parte, l’altra serie di teste si lega ad altri filamenti actinici puntando in direzione opposta e li tira dall’altra parte. L’effetto complessivo è quello di fare scorrere uno sopra l’altro fasci di actina orientati in senso opposto. Da questo si capisce come possa avvenire che un fascio, dove i filamenti di actina sono disposti così, generi una forza contrattile.

87
La Contrazione Muscolare
La contrazione volontaria dipende dai muscoli scheletrici; La contrazione involontaria come ad esempio il battito cardiaco e la peristalsi intestinale dipendono dal muscolo cardiaco e dalla muscolatura liscia, rispettivamente; In entrambi i casi l’actina e la miosina sono utilizzati in maniera analoga per contrarsi.

88
La Contrazione Muscolare
Le cellule del muscolo scheletrico sono multinucleate, hanno in genere un diametro di 50mm e sono lunghe parecchi centimetri; Il citoplasma è composto da miofibrille, che rappresentano l’unità contrattile della cellula muscolare, allineate in parallelo.

89
Ciascuna miofibrilla consiste di una catena di minuscole unità contrattili ovvero SARCOMERI che conferiscono alla miofibrilla dei vertebrati il caratteristico aspetto striato; I sarcomeri sono composti da due tipi di filamenti: i filamenti actinici e quelli di miosina di tipo II.

90
Il disco Z a ciascuna estremità del sarcomero sono punti di attacco dei filamenti sottili di actina;
La linea M indica la posizione di proteine specifiche che collegano filamenti spessi di miosina II fra loro; Le bande scure segnano la posizione dei filamenti spessi (miosina); Le bande chiare contengono soltanto i filamenti sottili Actina).
; Le bande chiare contengono soltanto i filamenti sottili Actina).")
91
L’accorciamento del sarcomero è dovuto allo scivolamento dei filamenti di miosina sui filamenti di actina senza cambiamento nella lunghezza dei due tipi di filamenti. Questo modello dello scivolamento dei filamenti è stato proposto per la prima volta nel 1954 ed è stato cruciale per la comprensione dei meccanismi della contrazione.

92
La contrazione muscolare è dovuta ad un simultaneo accorciamento dei sarcomeri ovvero allo slittamento dei filamenti di actina sopra quelli di miosina senza che vi sia variazione di lunghezza. Nel momento in cui un muscolo viene contratto, le teste miosiniche procedono lungo il filamento actinico mediante cicli ripetuti di attacco e distacco.

93
ATTACCO: La testa miosinica è rigidamente ancorata al filamento actinico nella configurazione di rigor.

94
RILASCIO: Una molecola di ATP si lega dietro la testa della miosina provocando da un lato un cambiamento nella conformazione dei domini che compongono il sito di legame e dall’altro una riduzione dell’affinità della testa miosinica per l’actina.

95
TRAZIONE: Il solco al quale è legato l’ATP si chiude inducendo così un cambiamento di conformazione che fa spostare la testa lungo il filamento; l’ATP si idrolizza, ma l’ADP ed il fosfato inorganico restano legati alla proteina.

96
GENERAZIONE della FORZA: Il debole legame della testa di miosina con un nuovo sito sul filamento di actina fa rilasciare il fosfato organico scatenando così il colpo di potenza durante il quale la testa riacquista la sua conformazione originale.

97
ATTACCO: Alla fine del ciclo, la testa della miosina è di nuovo legata saldamente al filamento di actina in una configurazione rigor. Il risultato conduce all’efficace spostamento della testa di miosina sul filamento di actina.

98
Il segnale proveniente da una terminazione nervosa innesca un potenziale d’azione nella membrana plasmatica della cellula muscolare. Il segnale passa poi dalla membrana a ciascun sarcomero attraverso i tubuli T ed il reticolo sarcoplasmatico.

99
Il reticolo è molto ricco in vescicole contenenti calcio che, in risposta all’eccitazione elettrica in arrivo, viene liberato nel citosol attraverso i canali ionici. Essi, infatti, si aprono dalla sua membrana alla variazione di voltaggio della membrana plasmatica stessa.

100
Nel caso di una miofibrilla normale, il Ca++ interagisce con un interruttore molecolare fatto di proteine accessorie, specializzate per interagire con i filamenti actinici come ad esempio la Tropomiosina e la Troponina. L’attacco del Ca++ alla Troponina è responsabile del rilascio del blocco, dovuto alla tropomiosina, delle interazioni sostenute dalle teste delle miosine con le molecole di actina. Se la miosina viene mescolata a filamenti puri di actina in provetta l’attività ATPasica della miosina viene stimolata indipendentemente dalla presenza di Ca++.


















>")
 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")


