Scaricare la presentazione
1
Lez 9 Control of Gene Expression –1 procarioti

2
Regolazione genetica nei procarioti
Operone del triptofano

3
I principi della regolazione trascrizionale
Nei batteri i geni sono controllati principalmente da molecole dell’ambiente, che agiscono su proteine regolatrici: attivatori o repressori. Il repressore si lega all’operatore sul DNA L’attivatore si lega ad un sito vicino al promotore ed aiuta il reclutamento della RNA polimerasi. Alcuni attivatori agiscono con meccanismo allosterico, inducendo il passaggio da complesso chiuso ad aperto

4
Reclutamento della RNA polimerasi.
In assenza di attivazione e di repressione la RNA polimerasi si lega occasionalmente al promotore ed inizia un basso livello di espressione, livello basale. Il legame del repressore alla sequenza dell'operatore blocca il legame delle RNA polimerasi e quindi inibisce la trascrizione. Il reclutamento delle RNA polimerasi da parte dell'attivatore porta ad elevati livelli di trascrizione.

5
Attivazione allosterica della RNA polimerasi.
La RNA polimerasi è legata al promotore in un complesso chiuso stabile. L'attivatore interagisce con la polimerasi promuovendo la trascrizione a complesso aperto ed elevati livelli di trascrizione.

6
L'interazione tra proteine legate al DNA.
Il legame cooperativo tra proteine legate a siti adiacenti. Legame cooperativo tra proteine posizionate su siti distanti

7
Una proteina che curva il DNA (architettonica) può facilitare l'interazione tra due proteine che legano il DNA. Una proteina che curva il DNA si lega su di un sito collocato tra il sito di legame dell'attivatore ed il promotore. Questo legame avvicina nello spazio due siti e quindi favorisce l'interazione tra l'attivatore legato al DNA e la polimerasi.

8
Procarioti: meccanismi di controllo

9
Operone Lac L’operone di Lac contiene tre ORF: lacZ: B-galattosidasi, lacY: permeasi del lattosio, lacA: trigalattoside transacetilasi. È regolato dal repressore Lac: lacI Anche CAP (Catabolite Activator Protein) regola l’operone. È un recettore di cAMP. Il repressore Lac lega il DNA solo in assenza di lattosio, CAP è un attivatore, e lega il DNA solo in assenza di glucosio, quando cAMP è alto.
 regola l’operone. È un recettore di cAMP. Il repressore Lac lega il DNA solo in assenza di lattosio, CAP è un attivatore, e lega il DNA solo in assenza di glucosio, quando cAMP è alto.")
10
L'operone Lac. I tre geni lacZ, LlcY e lacA sono trascritti come un unico mRNA dal promotore. Il sito CAP e l'operatore sono lunghi circa 20 bp ciascuno. L'operatore si trova sul promotore nella regione legata dalla RNA polimerasi e di sito CAP si trova proprio a monte del promotore.

11
L'espressione dei geni lac.
La presenza o l'assenza degli zuccheri lattosio e di glucosio controlla il livello di espressione dei geni lac. Livelli elevati di espressione necessitano la presenza del lattosio (quindi la assenza di un repressore lac funzionale) e l'assenza della preferenziale forma di energia, il glucosio (e quindi la presenza dell'attivatore CAP). Quando è legato all'operatore il repressore Lac esclude la polimerasi, indipendentemente dalla presenza di CAP attivo. CAP porta la polimerasi sul promotore sul promotore lac dove va incontro ad una isomerizzazione spontanea a compresso aperto.
 e l assenza della preferenziale forma di energia, il glucosio (e quindi la presenza dell attivatore CAP). Quando è legato all operatore il repressore Lac esclude la polimerasi, indipendentemente dalla presenza di CAP attivo. CAP porta la polimerasi sul promotore sul promotore lac dove va incontro ad una isomerizzazione spontanea a compresso aperto.")
12
Le due metà simmetriche dell'operatore lac
La regione di controllo dell'operone lac. Sono mostrate le sequenze nucleotidiche e l'organizzazione della regione di controllo dell'operone lac. Le barre colorate sotto e sopra indicano le regioni coperte dalla RNA polimerasi e dalle proteine regolatrici. Si noti che il repressore lac copre più DNA della sequenza definita come il minimo sito di legame dell'operatore, e la RNA polimerasi copre più DNA di quello definito dalle sequenze che formano il promotore.

13
Lambda repressor Può agire da attivatore o repressore

14
L'attivazione del promotore lac da parte di CAP.
Il legame dell’RNA polimerasi al promotore lac con l'aiuto di CAP. Il CAP è riconosciuto dalla CTD della subunità alfa. Quando interagisce con il CAP l'alfa-CTD contatta anche il DNA adiacente al sito CAP.

15
struttura del complesso CAP-alfa CTD-DNA
struttura del complesso CAP-alfa CTD-DNA. CAP è mostrato legato al suo sito sotto forma di dimero. Inoltre viene mostrato l‘alfa-CTD della RNA polimerasi legata ad un pezzo di DNA adiacente e mentre interagisce con il CAP. Il sito di interazione su ciascuna proteina coinvolge residui identificati geneticamente. CAP è in turchese alfa-CTD in viola. Ad ogni molecola di CAP è legata una molecola di ATP

16
Il legame al DNA di una proteina con il dominio helix-turn-helix.
Una proteina lega DNA come dimero e le due subunità sono indicate dai cerchi ombreggiati. L'elica di riconoscimento è indicata con R. Legame idrogeno sono instaurati tra il repressore di lambda e le coppie di base nel solco maggiore del suo operatore.

17
Il repressore lac si lega ai due operatori come tetramero
L’ansa si forma tra le repressore legato all'operatore principale e quello a monte, ausiliario a 90 bp. Un'ansa del tutto simile si può formare in alternativa con un operatore a valle, a 400 bp

18
L’attività di Lac repressor e CAP sono regolate allostericamente
Il lattosio si trasforma in allolattosio e si lega al lac repressor. In assenza di allolattosio il repressore è attivo, e si lega al DNA Il legame con allolattosio ne fa cambiare la conformazione, che non è più competente a legare il DNA. Il sito di legame è diverso da quello del DNA. La presenza di glucosio mantiene bassa la concentrazione di cAMP. In basso glucosio cAMP aumenta, si lega a CAP e lo rende competente a legare il DNA, e ad agire da attivatore. CAP regola vari altri geni, tra cui cui quelli dell’operone Galattosio.

19
Duplice controllo dell’operon di lac

20
Fattori sigma alternativi
Il fago batterico SPO1, per regolare l'espressione del proprio genoma utilizza tre fattori sigma in successione. Questo fa sì che i geni virali siano espressi esattamente nello stesso ordine in cui sono richiesti. In E. coli sigma 32 è inducibile al calore. Esso spiazza sigma70.

21
Attivazione mediata da NtrC: esempio di allosteria
NtrC controlla geni del metabolismo dell’azoto come glnA. La polimerasi è legata al promotore, con sigma54, nel complesso chiuso. NtrC interagisce con una subunità s54 della polimerasi, ne induce un cambiamento conformazionale con la transizione a complesso aperto, con attività ATPasica. A basso azoto NtrC viene fosforilato e diventa competente a legare il DNA nel sito a 150 bp dal promotore.

22
MerR ruota il DNA promotore
Gli elementi -10 e -35 del promotore merT sono esposti su due facce diverse della doppia elica perché distano 19 invece delle bp. Quando Hg è assente MerR si lega e stabilizza la forma inattiva del promotore. Quando Hg è presente MerR ruota il DNA in modo da allineare nel modo appropriato gli elementi del promotore.

23
Struttura di un promotore simile a merT.
Promotore con uno spacer di 19 bp. Il promotore con uno spacer di 19 bp compensato con l'attivatore funzionale. Promotore con uno spacer di 17 bp. Il promotore mostrato è quello del genere bmr di Bacillus subtilis controllato dal regolatore di bmr. questo funziona come attivatore quando si lega al composto TPP

24
Alcuni repressori trattengono la RNA polimerasi
Esempio: il repressore Gal di E. coli che controlla enzimi del metabolismo del galattosio. In assenza di Galattosio, il repressore interagisce con la Pol e ne impedisce la transizione a complesso aperto.

25
Antiattivazione: AraC
L’operone araBAD di E. coli è attivato dall’arabinosio. L’arabinosio si lega ad AraC inducendo una modificazione conformazionale nell'attivatore così che questi si lega come dimero ai siti araI1 ed araI2. In questo modo un monomero di AraC si posiziona vicino al promotore in modo da poter attivare la trascrizione. Quando l’arabinosio non è presente il dimero AraC adotta una diversa conformazione e si lega araO2 ed a araI1. In questa posizione non c'è nessun monomero sul sito araI2 quindi le proteina non può attivare il promotore araBAD. Questo promotore è regolato anche da CAP

26
Operone del triptofano
l'operone triptofano di E.coli codifica per il leader e per geni strutturali che codificano gli enzimi trp

27
Trp si lega al Trp-repressor
Il legame con Trp determina un cambiamento conformazionale

28
La attenuazione Anche con basso Trp molti tradotti terminano prematuramente, prima del gene trpE. Solo la sequenza leader viene tradotta. Essa è seguita da un terminatore Ci sono 4 sequenze complementari nel mRNA 1, 2, 3 e 4. Esse si possono appaiare diversamente L’RNA ha una ORF di 14 amino acidi, con una forte RBS e che codifica per 2 trp.

29
In condizioni di elevata concentrazione di triptofano la sequenza 3 si appaia con la sequenza 4 costituendo la hairpin di terminazione della traduzione. In condizioni di bassa concentrazione di triptofano il ribosoma si blocca sugli adiacenti codoni per il triptofano permettendo alla sequenza 2 di appaiarsi con la 3 e quindi impedendo la formazione della forcine di terminazione 3 e 4. Quindi la traduzione può continuare In assenza di sintesi proteica poiché il ribosoma non inizia la traduzione della AUG del peptide leader si forma una forcina appaiando le sequenze 1 e 2 e quindi non si forma quella data dalle sequenze 2 e 3 ne consegue che si forma una forcina con le sequenza 3 4 e gli enzimi per trp non sono espressi

30
Riconoscimento proteine-DNA

31
DNA-binding sites

32
DNA piegato DNA mitocondriale di tripanosoma
AAAANNN repeats formano molecole curve

33
DNA può essere deformato
CAP: catabolite activator protein 434 repressor

34
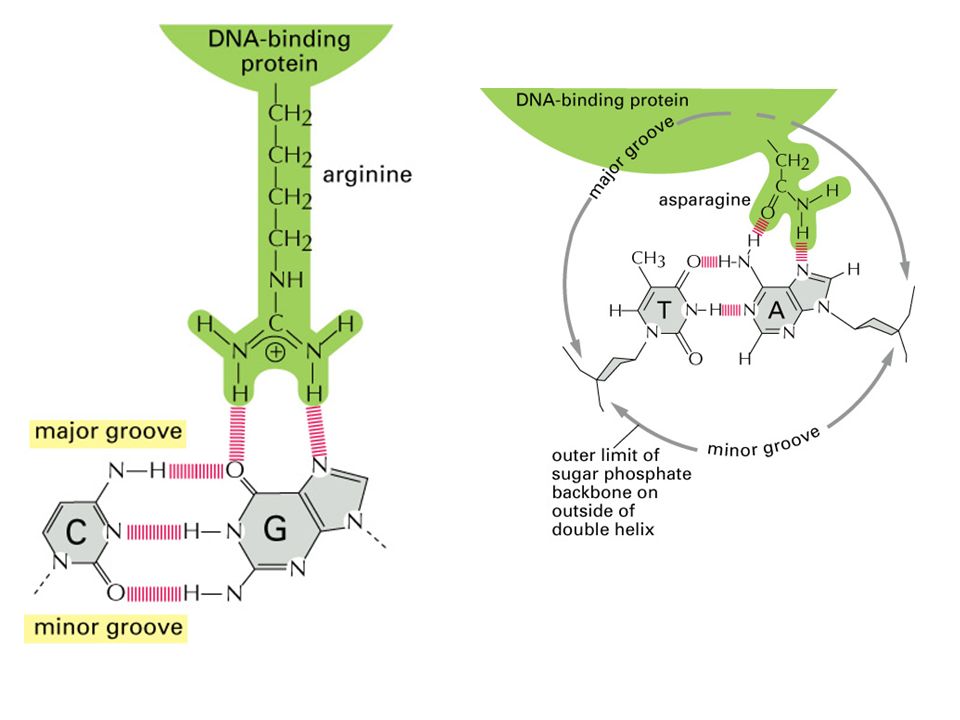
Arg e Asp Aspargina lega A Arginina lega G

35
Helix-turn-helix HTH è una struttura molto diffusa per DNA-binding. Anche nei procarioti

36
Esempi di helix-turn-helix
Sequenza riconosciuta da lambda Cro

37
DNA-binding domains Il zinc finger è un DNA-binding domain.
Gli steroid receptors: la proteina è inattiva finchè non lega un piccolo ligando. helix-turn-helix negli homeodomain, La helix-loop-helix (HLH) anfipatica. La lunghezza del loop varia da amino acidi. Il motivo permette la dimerizzazione la presenza di una regione basica per il contatto con il DNA. Leucine zippers. Consiste in una sequenza di amino acidi con una leucina ogni sette aa per formare un dimero. Vicino ad ogni zipper c’è una regione basica per il contatto con il DNA
 anfipatica. La lunghezza del loop varia da amino acidi. Il motivo permette la dimerizzazione la presenza di una regione basica per il contatto con il DNA. Leucine zippers. Consiste in una sequenza di amino acidi con una leucina ogni sette aa per formare un dimero. Vicino ad ogni zipper c’è una regione basica per il contatto con il DNA.")
38
Struttura homeodomain
Hanno la struttura base HLH.

39
Homeodomains bind related targets in DNA
· The homeodomain is a DNA-binding domain of 60 amino acids that has three a-helices. · The C-terminal a-helix-3 is 17 amino acids and binds in the major groove of DNA. · The N-terminal arm of the homeodomain projects into the minor groove of DNA. · Proteins containing homeodomains may be either activators or repressors of transcription.

40
Eterodimero di due homeodomain

41
Due domini uniti covalentemente

42
Il mitivo zinc finger è un DNA-binding domain
Uno zinc finger è un’ansa di ~23 amino acidi che protrude da un zinc-binding site formato da His and Cys. Una proteina zinc finger di solito ha molti zinc fingers. La parte C-terminale di ogni finger forma una a-elica che lega il solco maggiore del DNA. Alcune Some zinc finger proteins legano anche RNA.

43
Zinc finger Tipo Cys-Cys-His-His

44
Cys-X2-4-Cys-X3-Phe-X5-Leu-X2-His-X3-His
Zinc finger Una "finger protein" normalmente ha una serie di zinc fingers. la consensus sequence di un singolo finger è: Cys-X2-4-Cys-X3-Phe-X5-Leu-X2-His-X3-His

45
Legame al DNA di zinc finger
Zinc fingers possono legare sequenze estese di DNA

46
Dimero zinc finger Alcune zinc fingers possono dimerizzare

48
Interazioni tra diversi zinc fingers e DNA

49
Beta sheet Proteina batterica: Met repressor

50
Leucine zipper Formazione di eterodimeri

51
Le Leucine zippers formano dimeri
La leucine zipper è una elica anfipatica che dimerizza. La zipper è adiacente ad una regione basica che lega il DNA. La dimerizzazione forma un motivo in cui le due regioni basiche legano simmetricamente degli inverted repeats nel DNA.

52
Helix-loop-helix proteins interact by combinatorial association
proteine Helix-loop-helix hanno un motivo di amino acidi che comprende due eliche anfipatiche di residui separate da un loop. Le eliche sono responsabili della formazione dei dimeri . Le proteine HLH hanno una sequenza basica adiacente il motivo HLH che è responsabile del legame al DNA. La classe A delle proteine HLH sono espresse in tutti i tessuti, la classe B è tessuto specifica Le proteine di classe B spesso formano eterodimeri con la classe A. Le proteine HLH che non hanno la regione basica impediscono al partner nell’eterodimero di legarsi al DNA. Le proteine HLH formano associazioni combinatoriali che possono variare durante lo sviluppo per la aggiunta e perdita di proteine specifiche

53
Helix-loop-helix