Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Genetica Mendeliana Presupposti
Tutte le piante ibride, discendenti da genitori geneticamente diversi e puri hanno un aspetto simile Gli ibridi incrociati fra di loro non generalo linee pure. I discendenti mostrano mescolanza di caratteri: alcuni sono simili ai genitori, altri alla generazione di partenza

2
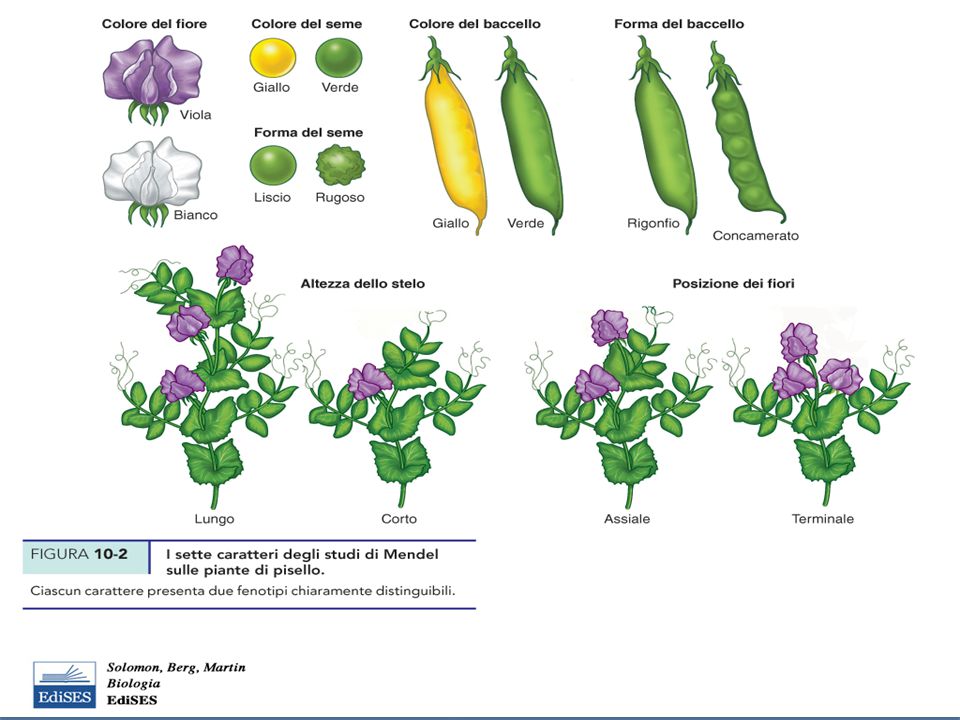
Genetica Mendeliana Mendel (1856) individuò regole precise sulla modalità di trasmissione dei caratteri alla progenie Come organismo scelse le piante di Pisum sativus: crescono facilmente, ve ne sono molte varietà, consentono impollinazione controllata Selezionò le linee pure per determinati caratteri (che vengono conservati nelle generazioni successive) Scelse varietà rappresentative per sette caratteri che mostrassero fenotipi diversi Limitò la variabilità genetica studiata in ciascun esperimento
 Scelse varietà rappresentative per sette caratteri che mostrassero fenotipi diversi. Limitò la variabilità genetica studiata in ciascun esperimento.")
3
Il fiore del pisello, si autoimpollina, ma le antere che producono il polline si possono rimuovere, e sullo stimma può essere posto il polline di un altro individuo

5
Incrociò piante di due differenti linee pure con fenotipi diversi (Generazione parentale, P)
La prima generazione filiale (F1) era costituita da individui tutti uguali che, per un certo carattere, somigliavano sempre ad un genitore La seconda generazione filiale (F2), prodotta per incrocio di individui della F1, mostrava per ¾ il carattere della F1, ma per ¼ il carattere della generazione P che si era perso nella generazione F1
 era costituita da individui tutti uguali che, per un certo carattere, somigliavano sempre ad un genitore. La seconda generazione filiale (F2), prodotta per incrocio di individui della F1, mostrava per ¾ il carattere della F1, ma per ¼ il carattere della generazione P che si era perso nella generazione F1.")
6
I fattori ereditari Mendel concluse che esistono dei “fattori ereditari” (caratteri dominanti) di un genitore che mascherano quelli dell’ altro genitore (recessivi) quando coesistono nella generazione F1 Questo concetto smontava l’ assunto che l’ ereditarietà fosse controllata di “fluidi” che si mescolavano per formare gli ibridi I Fattori di Mendel, capaci di controllare le caratteristiche ereditarie sono i geni
 di un genitore che mascherano quelli dell’ altro genitore (recessivi) quando coesistono nella generazione F1. Questo concetto smontava l’ assunto che l’ ereditarietà fosse controllata di fluidi che si mescolavano per formare gli ibridi. I Fattori di Mendel, capaci di controllare le caratteristiche ereditarie sono i geni.")
7
Principio della segregazione(I)
Definito l’ allele come forma alternativa di un gene, si possono individuare alleli dominanti e recessivi. Quando sono presenti entrambi il fenotipo è quello dell’ allele dominante. Il principio della segregazione stabilisce che, prima della riproduzione sessuata, gli alleli portati da un genitore si separano (segregano). Il gamete contiene un solo allele di ciascun paio. Gli alleli durante la segregazione rimangono intatti (non vengono contaminati), per cui gli alleli recessivi possono ricomparire nella generazione F2
. Il gamete contiene un solo allele di ciascun paio. Gli alleli durante la segregazione rimangono intatti (non vengono contaminati), per cui gli alleli recessivi possono ricomparire nella generazione F2.")
8
Principio della segregazione(II)
Da una generazione P (TT) e (tt), si ottiene una F1 (Tt) in cui domina il fenotipo T; la casualità della fecondazione porta in F2 a ¼ (TT) con fenotipo T, ½ (Tt) con fenotipo T, ¼ (tt) con fenotipo t. Nel 1902 Sutton e Boveri correlarono la segregazione mendeliana degli alleli, con la segregazione dei cromosomi omologhi alla meiosi
 e (tt), si ottiene una F1 (Tt) in cui domina il fenotipo T; la casualità della fecondazione porta in F2 a ¼ (TT) con fenotipo T, ½ (Tt) con fenotipo T, ¼ (tt) con fenotipo t. Nel 1902 Sutton e Boveri correlarono la segregazione mendeliana degli alleli, con la segregazione dei cromosomi omologhi alla meiosi.")
9
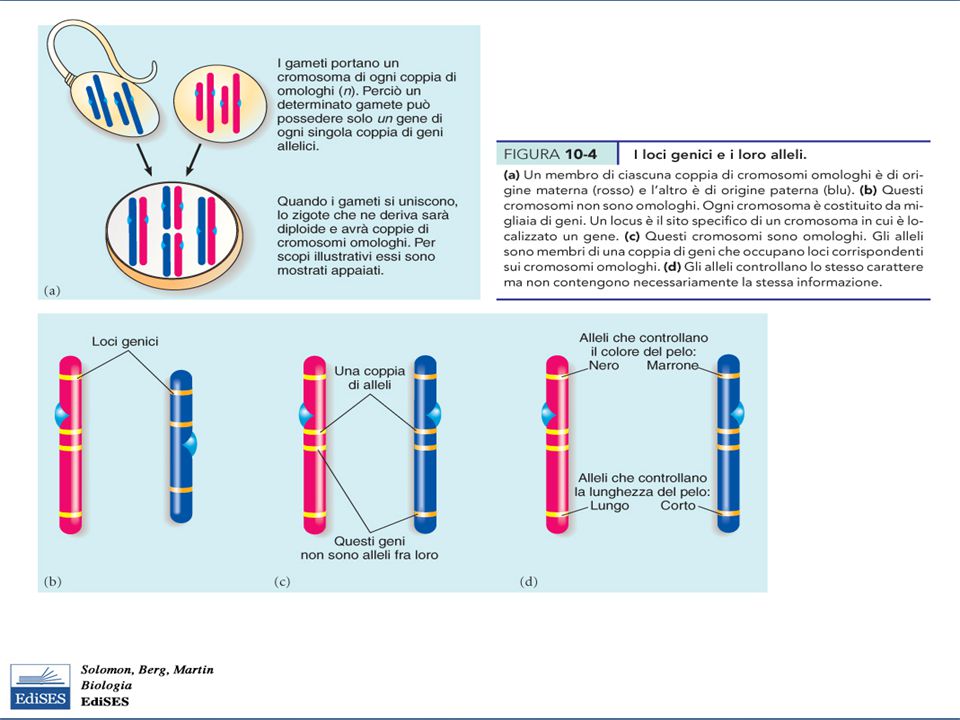
Loci genici Locus: indica la posizione di un particolare gene sul cromosoma, ma anche la porzione di DNA che possiede l’ informazione per il controllo di un carattere di un organismo Un locus può essere identificato se esistono almeno due varianti alleliche che producono fenotipi differenti Alleli: geni che controllano varianti diverse dello stesso carattere ed occupano loci corrispondenti sui cromosomi omologhi

11
Incroci monoibridi (I)
Sono incroci fra individui che portano alleli diversi di un singolo locus: femmina omozigote per il colore nero ( B allele dominante), maschio omozigote per il colore marrone (b allele recessivo). Per il principio della segregazione, alla meiosi si otterranno gameti femminili B e gameti maschili b. All’ F1, gli individui (Bb) avranno tutti pelo nero (dominante)
, maschio omozigote per il colore marrone (b allele recessivo). Per il principio della segregazione, alla meiosi si otterranno gameti femminili B e gameti maschili b. All’ F1, gli individui (Bb) avranno tutti pelo nero (dominante)")
12
Incroci monoibridi (II)
I soggetti dell’ F1 (Bb) per il principio della segregazione, alla meiosi produrranno gameti femminili e maschili B e b in ugual numero. La fecondazione è un fenomeno casuale e si ottiene il rapporto fenotipico della generazione F2 di un incrocio di monoibridi 3:1 Quadrato di Punnet: rappresentazione delle possibili combinazioni di gameti
 per il principio della segregazione, alla meiosi produrranno gameti femminili e maschili B e b in ugual numero. La fecondazione è un fenomeno casuale e si ottiene il rapporto fenotipico della generazione F2 di un incrocio di monoibridi 3:1. Quadrato di Punnet: rappresentazione delle possibili combinazioni di gameti.")
13
Leggi della Probabilità
Due eventi sono indipendenti quando il verificarsi dell’ uno non influenza la probabilità del verificarsi dell’ altro. Legge del prodotto: la probabilità che si verifichino entrambi contemporaneamente è pari al prodotto delle singole probabilità Se esiste più di un modo per avere un risultato (eventi escludentisi), la probabilità totale si ottiene sommando le singole probabilità: da due genitori Bb, si può avere un figlio Bb con uovo B e spermio b (¼) o con uovo b e spermio B (1/4) e la probabilità totale è 1/2
, la probabilità totale si ottiene sommando le singole probabilità: da due genitori Bb, si può avere un figlio Bb con uovo B e spermio b (¼) o con uovo b e spermio B (1/4) e la probabilità totale è 1/2.")
14
Fenotipo e Genotipo Non sempre dal fenotipo possiamo risalire al genotipo (individui BB e Bb sono entrambi a pelo nero) Il fenomeno della dominanza spiega perché si può somigliare di più ad un genitore anche se i due genitori contribuiscono in egual misura al genotipo della progenie La dominanza non è prevedibile e può essere determinata solo sperimentalmente In una popolazione non è detto che il fenotipo dominante sia più comune del fenotipo recessivo

15
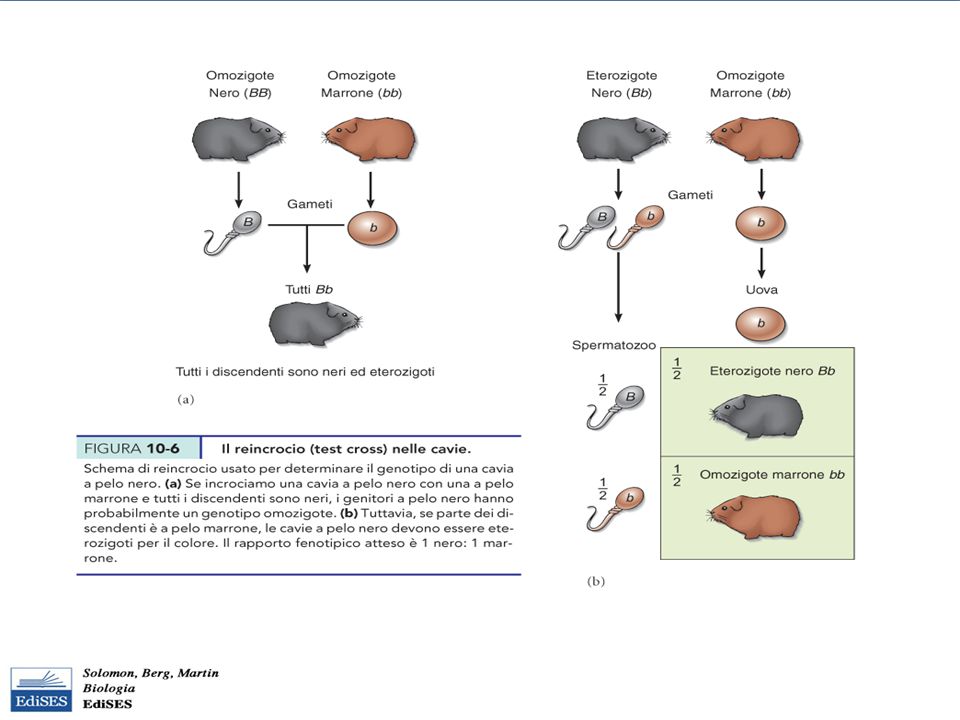
Reincrocio (test cross)
Serve a stabilire il genotipo della cavia a pelo nero (BB o Bb??) cioè per svelare la condizione di eterozigosi Si incrociano le cavie a pelo nero con individui a pelo marrone, (omozigoti per il carattere recessivo, bb) In un reincrocio, i due tipi di gameti prodotti dal genitore con genotipo sconosciuto, non sono “nascosti” nella progenie dagli alleli che provengono dall’ altro genitore (gameti tutti b) Il principio mendeliano della segregazione consente di pervedere il rapporto fenotipico dei reincroci
 cioè per svelare la condizione di eterozigosi. Si incrociano le cavie a pelo nero con individui a pelo marrone, (omozigoti per il carattere recessivo, bb) In un reincrocio, i due tipi di gameti prodotti dal genitore con genotipo sconosciuto, non sono nascosti nella progenie dagli alleli che provengono dall’ altro genitore (gameti tutti b) Il principio mendeliano della segregazione consente di pervedere il rapporto fenotipico dei reincroci.")
17
Incrocio diibrido Incrocio fra individui che differiscono per gli alleli di due loci Nel caso in cui le coppie di alleli sono localizzate su cromosomi non omologhi, ogni coppia di alleli alla meiosi segrega indipendentemente dall’ altra

18
Principio dell’ assortimento indipendente
I membri di una coppia di geni segregano indipendentemente dai membri di un’ altra coppia Ogni gamete contiene un singolo allele per ogni locus, ma gli alleli di loci differenti vengono assortiti nei gameti in modo casuale l’ uno rispetto all’ altro Ogni cavia della F1 produce 4 tipi di gameti (BS,Bs,bS,bs) con uguale probabilità Nel quadrato di Punnet ci sono 42 =16 quadrati Se i loci per la lunghezza del pelo e il colore del pelo sono su cromosomi non omologhi, nella generazione F2 di un incrocio diibrido, il rapporto fenotipico è 9:3:3:1
 con uguale probabilità. Nel quadrato di Punnet ci sono 42 =16 quadrati. Se i loci per la lunghezza del pelo e il colore del pelo sono su cromosomi non omologhi, nella generazione F2 di un incrocio diibrido, il rapporto fenotipico è 9:3:3:1.")
19
L’ assortimento indipendente è correlato agli eventi della meiosi
Ci sono due modi diversi in cui due coppie di Ch omologhi possono essere disposte alla metafase I che li distribuisce nelle due cellule figlie Tale disposizione è casuale L’orientamento dei Ch omologhi in piastra determina il modo in cui si separeranno e verranno distribuiti nei gameti

20
La segregazione degli alleli è il risultato diretto della separazione dei cromosomi omologhi nel corso della meiosi

21
Geni associati (I) Thomas Morgan (1910)usò il moscerino della frutta, Drosophila melanogaster, per dimostrare che i geni sono disposti sui cromosomi in maniera lineare L’ assortimento indipendente non è applicabile quando due loci genici sono situati sulla stessa coppia di cromosomi omologhi, specialmente se non sono distanti. Questi loci genici sono associati e la loro tendenza ad essere ereditati insieme si definisce linkage
usò il moscerino della frutta, Drosophila melanogaster, per dimostrare che i geni sono disposti sui cromosomi in maniera lineare. L’ assortimento indipendente non è applicabile quando due loci genici sono situati sulla stessa coppia di cromosomi omologhi, specialmente se non sono distanti. Questi loci genici sono associati e la loro tendenza ad essere ereditati insieme si definisce linkage.")
22
Geni associati (II) Drosophila ha un locus che controlla la forma delle ali (V, dominante per le ali normali; v, recessivo per le ali vestigiali), ed un locus che controlla il colore del corpo (B, dominante per il grigio; b, recessivo per il nero) Dall’ incrocio BBVV (corpo grigio, ali normali) con bbvv (corpo nero, ali vestigiali), si ottiene una F1 con genotipo BbVv e fenotipo corpo grigio, ali normali Il Linkage si osserva dal risultato del reincrocio a due punti, riguardante cioè due loci genici: generazione F1 (BbVv) con omozigote recessivo (bbvv)
, ed un locus che controlla il colore del corpo (B, dominante per il grigio; b, recessivo per il nero) Dall’ incrocio BBVV (corpo grigio, ali normali) con bbvv (corpo nero, ali vestigiali), si ottiene una F1 con genotipo BbVv e fenotipo corpo grigio, ali normali. Il Linkage si osserva dal risultato del reincrocio a due punti, riguardante cioè due loci genici: generazione F1 (BbVv) con omozigote recessivo (bbvv)")
23
Reincrocio a due punti (I)
Se i loci fossero stati non associati (su Cromosomi differenti), i gameti BV,bv (parentali = a quelli della generazione P) e Bv,bV (ricombinanti ≠ da quelli della generazione P) sarebbero stati di numero uguale e la progenie avrebbe mostrato i 4 fenotipi con uguale frequenza I loci associati vengono ereditati come unità e se l’associazione fosse totale avremmo solo moscerini di tipo parentale: 1/2 grigio, normale (BbVv); ½ nero, vestgiale (bbvv)
, i gameti BV,bv (parentali = a quelli della generazione P) e Bv,bV (ricombinanti ≠ da quelli della generazione P) sarebbero stati di numero uguale e la progenie avrebbe mostrato i 4 fenotipi con uguale frequenza. I loci associati vengono ereditati come unità e se l’associazione fosse totale avremmo solo moscerini di tipo parentale: 1/2 grigio, normale (BbVv); ½ nero, vestgiale (bbvv)")
24
Reincrocio a due punti (II)
La progenie ricombinante (grigio- vestigiale, nero-normale) ha ricevuto un gamete ricombinante dal genitore della F1 I gameti ricombinanti originano dai Crossing Over (scambio di materiale genetico fra cromatidi omologhi nella I profase meiotica) In Drosophila i Crossing Over avvengono solo nelle femmine
 ha ricevuto un gamete ricombinante dal genitore della F1. I gameti ricombinanti originano dai Crossing Over (scambio di materiale genetico fra cromatidi omologhi nella I profase meiotica) In Drosophila i Crossing Over avvengono solo nelle femmine.")
25
% di ricombinazione Nella progenie il numero di ricombinanti rispetto ai genotipi parentali dipende dal numero di crossing over Due loci hanno una % di ricombinazione che si calcola: N° ricombinanti/N° totale di figli% In genere più due loci sono distanti, più frequentemente può avvenire un crossing over

26
Mappe geniche Per convenzione 1 unità di mappa = 1% di ricombinazione fra due loci genici

27
Doppio Crossing Over In realtà, la frequenza di crossing over è leggermente maggiore di quella dei gameti ricombinanti Infatti il verificarsi contemporaneo di due crossing over sugli stessi due cromatidi omologhi, ricostituisce la combinazione genica originale Su due loci molto vicini il doppio crossing over è assai improbabile

28
Determinismo del sesso (I)
Il sesso in alcune specie è controllato dall’ ambiente, ma di solito è determinato da geni che mappano sui cromosomi sessuali In molte specie animali, le femmine hanno 2 cromosomi X, i maschi hanno XY. Tutte le altre coppie di cromosomi si definiscono Autosomi

29
Determinismo del sesso (II)
Tutti gli individui per vivere hanno bisogno almeno di un X (Y0 non è vitale) Y determina il sesso maschile (XXY,Klinefelter, è fenotipicamente maschio, X=, Turner è fenotipicamente femmina Il gene SRY è localizzato su Y, e causa nel feto lo sviluppo dei testicoli. Questi producono il testosterone che induce lo sviluppo del fenotipo maschile Altri geni su Y, geni su X e geni sugli autosomi sono coinvolti nel determinismo del sesso
 Y determina il sesso maschile (XXY,Klinefelter, è fenotipicamente maschio, X=, Turner è fenotipicamente femmina. Il gene SRY è localizzato su Y, e causa nel feto lo sviluppo dei testicoli. Questi producono il testosterone che induce lo sviluppo del fenotipo maschile. Altri geni su Y, geni su X e geni sugli autosomi sono coinvolti nel determinismo del sesso.")
30
Determinismo del sesso (III)
Probabilmente XY originariamente erano una coppia di omologhi, attualmente non sono omologhi, ma hanno una regione di omologia (di appaiamento) che permette loro di segregare durante la meiosi Il rapporto maschi/femmine sia concepiti che nati, è leggermente a favore dei maschi (forse lo spermatozoo che porta Y ha qualche vantaggio competitivo) Drosophila ha maschi XY, ma Y non determina la mascolinità In uccelli e farfalle i maschi sono XX e le femmine XY
 che permette loro di segregare durante la meiosi. Il rapporto maschi/femmine sia concepiti che nati, è leggermente a favore dei maschi (forse lo spermatozoo che porta Y ha qualche vantaggio competitivo) Drosophila ha maschi XY, ma Y non determina la mascolinità. In uccelli e farfalle i maschi sono XX e le femmine XY.")
31
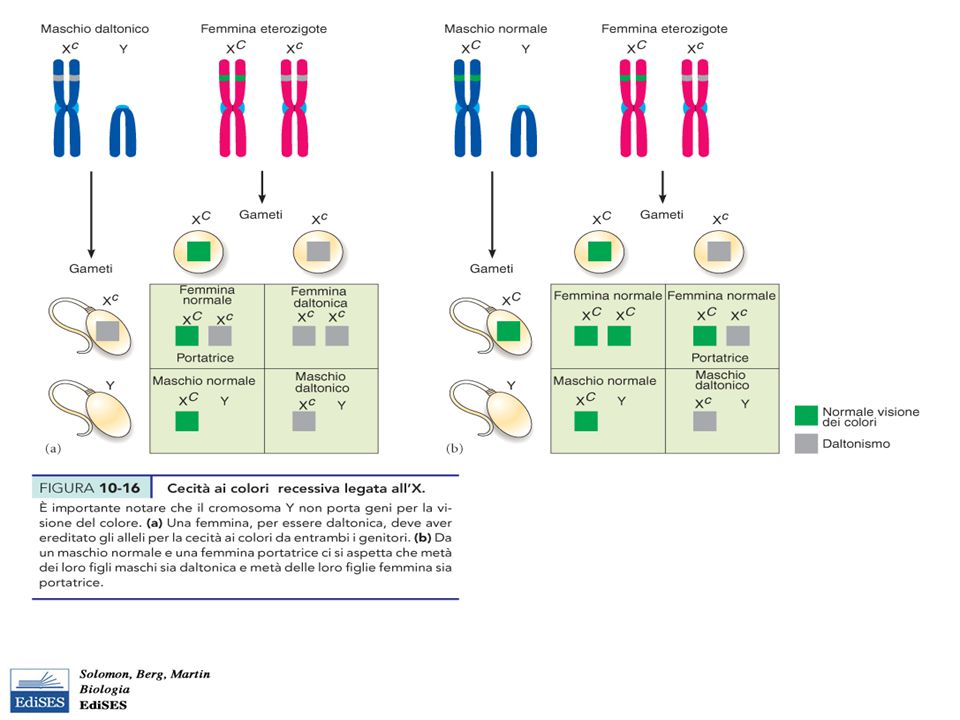
Geni X-linked Ci sono dei caratteri non sessuali (percezione dei colori, coagulazione del sangue) i cui geni mappano sul Cromosoma X (X-linked) Il maschio XY, ha per il Ch X una condizione di emizigosi (una sola copia), per cui l’allele, dominante o recessivo che sia, viene sempre espresso, mentre nella femmina, l’ etrozigote non esprime il carattere recessivo Per questo motivo, i caratteri recessivi associati al Cromosoma X si esprimono più nei maschi che nelle femmine
, per cui l’allele, dominante o recessivo che sia, viene sempre espresso, mentre nella femmina, l’ etrozigote non esprime il carattere recessivo. Per questo motivo, i caratteri recessivi associati al Cromosoma X si esprimono più nei maschi che nelle femmine.")
33
Trasmissione emofilia: madre portatrice e padre sano
XE X X Y XE X X X XE Y X Y Femmina portatrice Femmina sana Maschio emofilico Maschio sano

34
Trasmissione emofilia: madre sana e padre emofilico
X X XE Y X XE X XE X Y X Y Femmina portatrice Maschio sano Maschio sano

35
Trasmissione emofilia: madre portatrice e padre emofilico
XE X XE Y XE XE X XE XE Y X Y Femmina emofilica Femmina portatrice Maschio emofilico Maschio sano

36
Compensazione del dosaggio
Nelle specie in cui la femmine ha XX e il maschio ha un solo X, esistono dei meccanismi di compensazione del dosaggio che rendono equivalente l’ espressione di X nei due sessi In Drosophila il maschio incrementa l’ attività del suo unico X Nei mammiferi la femmina inattiva uno dei due X

37
Dominanza incompleta e codominanza
Dominanza incompleta: l’ eterozigote mostra un fenotipo intermedio Codominanza: l’ eterozigote mostra il fenotipo di entrambi i genitori Il gruppo sanguigno è controllato da tre alleli di un singolo locus: IA,IB,i. IAeIB sono codominanti, ma entrambi sono dominanti su i

38
Alleli multipli Un locus ha alleli multipli quando in una popolazione esistono tre o più locus In seguito a studi di incroci, si stabilisce l’ ordine di dominanza (C>cch>ch>c) Alleli multipli possono presentare codominanza o dominanza incompleta (eterozigoti con fenotipo intermedio)
 Alleli multipli possono presentare codominanza o dominanza incompleta (eterozigoti con fenotipo intermedio)")
39
Pleiotropia La maggior parte dei geni ha più di un effetto fenotipico (pleiotropia) In genere i diversi effetti sono riconducibili ad una stessa causa (es. deficit enzimatico) Es. sintomatologia variegata di malattie genetiche
 Es. sintomatologia variegata di malattie genetiche.")
40
Interazione genica Coppie di alleli diversi possono interagire nell’ espressione di un carattere o una coppia può inibire o cambiare l’ effetto di un’ altra

41
Epistasi la presenza di determinati alleli in un locus impedisce o maschera l’ espressione di altri alleli di un altro locus

42
Eredità poligenica Più coppie di geni indipendenti hanno effetti simili e additivi sullo stesso carattere (60 loci diversi per il colore della pelle) In F1 si hanno caratteristiche intermedie a quelle della generazione parentale omozigote; in F2 di ha elevata variabilità con una curva di distribuzione a campana

43
Interazioni con l’ ambiente
Fattori genetici ed ambientali interagiscono per determinare il fenotipo Quale l’ influenza dei geni e quale quella dell’ ambiente??? Norma di reazione : varietà di possibili fenotipi che si possono sviluppare da un unico genotipo in diverse condizioni ambientali

Presentazioni simili



 Dominanza incompleta>")





 è dominante sul pelo liscio ( r )>")





>")



