Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Capitolo 2: Biosintesi dei lipidi

2
Ruoli dei lipidi Conservazione energia Costituenti membrane cellulari
Pigmenti (retinale, carotene) Cofattori (vitamina K) Detergenti (sali biliari) Trasportatori Ormoni (derivati della vitamina D; sessuali) Messagi intra- o extracellulari Ancore per proteine di membrane
 Cofattori (vitamina K) Detergenti (sali biliari) Trasportatori. Ormoni (derivati della vitamina D; sessuali) Messagi intra- o extracellulari. Ancore per proteine di membrane.")
3
Sistema navetta per il trasporto dei gruppi acetilici dai
mitocondri al citosol Nelson e Cox 6° ed. Figura 21.10: La membrana mitocondriali esterna è liberamente permeabile a tutti questi composti. Il piruvato prodotto dal catabolismo degli amminoacidi nella matrice mitocondriali, o derivante dal glucosio mediante la glicolisi nel citosol, viene convertito in acetil-CoA nella matrice. I gruppi acetilici escono dai mitocondri sotto forma di citrato; nel citosol, viene recuperato l’acetil-CoA per la sintesi degli acidi grassi. L’ossalacetato viene ridotto a malato, che ritorna nella matrice mitocondriale, dove viene convertito in ossalacetato. Il destino principale del malato nel citosol è l’ossidazione catalizzata dall’enzima malico, che produce NADPH citosolico; il piruvato prodotto ritorna nella matrice mitocondriale.

4
sintesi degli acidi grassi
CoA-SH acetil-CoA ADP + Pi ATP citrato liasi citrato ossalacetato citosol trasp. citrato mitocondrio citrato citrato sintasi ossalacetato CoA-SH acetil-CoA acetato aminoacidi piruvato glucoso piruvato acetaldeide etanolo

5
La citrato sintesi agisce nella matrice mitocondriale.
La citrato liasi agisce nel citosol.

6
sintesi degli acidi grassi piruvato carbossilasi
CoA-SH acetil-CoA ADP + Pi ATP NADH + H+ NAD+ NADP+ NADPH + H+ citrato liasi CO2 malDH enzima malico citrato ossalacetato malato piruvato trasp. citrato trasp. mal/αKglu trasp. piruvato citosol mitocondrio citrato malato piruvato citrato sintasi malDH ossalacetato CoA-SH NAD+ NADH + H+ piruvato carbossilasi acetil-CoA CO2 ADP + Pi ATP acetato aminoacidi piruvato glucoso piruvato acetaldeide etanolo

7
La malato deidrogenasi agisce nella matrice mitocondriale.
L’enzima malico agisce nel citosol.

8
Produzione di NADPH Negli epatociti e negli adipociti il NADPH citosolico è in grande parte generato dalla via del pentosio fosfato e dall’enzima malico. Nelson e Cox ed 6: Figura 21.9: Due diverse vie per la produzione di NADPH, una (a) catalizzata dall’enzima malico e l’altra (b) tramite la via del pentosio fosfato.
 catalizzata dall’enzima malico e l’altra (b) tramite la via del pentosio fosfato.")
9
Reazione dell’acetil-CoA carbossilasi 1:
ADP + P i O O CO ATP 2 C CH biotina CH 3 2 C O C O acetil-CoA S-CoA carbossilasi S-CoA (ACC) acetilcoenzima A malonilcoenzima A
 acetilcoenzima A. malonilcoenzima A.")
10
Reazione dell’acetil-CoA carbossilasi 2:
Nelson/Cox 5th FIGURE 21-1 The acetyl-CoA carboxylase reaction. Acetyl-CoA carboxylase has three functional regions: biotin carrier protein (gray); biotin carboxylase, which activates CO2 by attaching it to a nitrogen in the biotin ring in an ATP-dependent reaction (see Figure 16-16); and transcarboxylase, which transfers activated CO2 (shaded green) from biotin to acetyl-CoA, producing malonyl-CoA. The long, flexible biotin arm carries the activated CO2 from the biotin carboxylase region to the transcarboxylase active site. The active enzyme in each step is shaded blue. L’acetil-CoA carbossilasi ha 3 regoni funzionali: la proteina trasportatrice della biotina (in grigio); la biotina carbossilasi che attiva la CO2 e la lega a uno degli atomi di azoto dell’anello della biotina in una reazione ATP-dipendente; la transcarbossilasi, che trasferisce la CO2 attivata (in verde) dalla biotina all’acetil-CoA, formando il malonil CoA. Il lungo braccio flessibile trasporta la CO2 attivata dalla regione della biotina carbossilasi al sito attiva della transcarbossilasi. Il sito attivo di ogni tappa è ombreggiato in celeste.
; biotin carboxylase, which activates CO2 by attaching it to a nitrogen in the biotin ring in an ATP-dependent reaction (see Figure 16-16); and transcarboxylase, which transfers activated CO2 (shaded green) from biotin to acetyl-CoA, producing malonyl-CoA. The long, flexible biotin arm carries the activated CO2 from the biotin carboxylase region to the transcarboxylase active site. The active enzyme in each step is shaded blue. L’acetil-CoA carbossilasi ha 3 regoni funzionali: la proteina trasportatrice della biotina (in grigio); la biotina carbossilasi che attiva la CO2 e la lega a uno degli atomi di azoto dell’anello della biotina in una reazione ATP-dipendente; la transcarbossilasi, che trasferisce la CO2 attivata (in verde) dalla biotina all’acetil-CoA, formando il malonil CoA. Il lungo braccio flessibile trasporta la CO2 attivata dalla regione della biotina carbossilasi al sito attiva della transcarbossilasi. Il sito attivo di ogni tappa è ombreggiato in celeste.")
11
β-chetoacilCoA sintasi (KS) malonil/acetil-CoA ACP transferasi (MAT)
Acido grasso sintasi: fosfopanteteina SH SH SH KS TE SH MAT ACP KR DH ER β-chetoacilCoA sintasi (KS) malonil/acetil-CoA ACP transferasi (MAT) β-idrossiacil-ACP deidratasi (DH) enoil-ACP reduttasi (ER) β-chetoacil-ACP reduttasi (KR) tioesterasi (TE) acyl carrier protein (ACP)
 malonil/acetil-CoA ACP transferasi (MAT) β-idrossiacil-ACP deidratasi (DH) enoil-ACP reduttasi (ER) β-chetoacil-ACP reduttasi (KR) tioesterasi (TE) acyl carrier protein (ACP)")
13
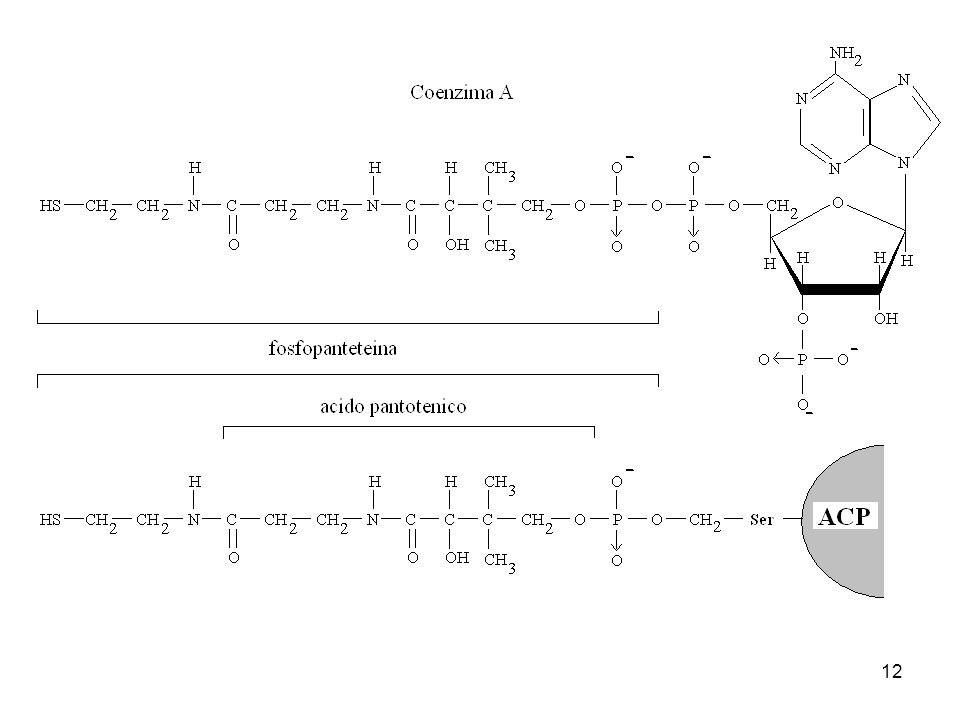
La proteina trasportatrice di acili (ACP):
Il gruppo prostetico è la 4’-fosfopanteteina legata covalentemente al gruppo ossidrilico di un residuo di Ser dell’ACP. Il suo gruppo SH è il sito di ingresso dei gruppi malonilici durante la sintesi degli acidi grassi.

14
D-β-idrossiacil-ACP MAT ACP DH KS KR ER
Nelson e Cox 6° ed. Figura 21.2: Aggiunta di due atomi di carbonio a una catena acilica in fase di crescita: una sequenza di reazioni suddivisa in 4 tappe. Ciascun gruppo malonilico e acetilico (o gruppi acetilici più lunghi) viene attivato da un tioestere, che lo lega alla acido grasso sintasi. 1- la condensazione di un gruppo acetilico attivato (il gruppo acetilico dell’acetil-CoA fornirà il primo gruppo acilico) e di un frammento a 2 atomi di C derivato dal malonil-CoA, con eliminazione di CO2 dal gruppo malinico, allunga la catena acilica di 2 C. il meccanismo del primo passaggio di questa reazione è qui presentato al fine di illustrare il ruolo della decarbossilazione nei processi di condensazione. Il derivato Β-cheto, prodotto dalla reazione di condensazione viene ridotto mediante altri tre passaggi, quasi identici alle reazioni di B ossidazione, ma in sequenza inversa. 2- il gruppo chetonico B viene ridotto ad alcol. 3- l’eliminazione di H2O crea un doppio legame. 4- il doppio legame viene ridotto, per formare il corrispondente gruppo acilico saturo. Sia il cofattore trasportatore di e, sia i gruppi attivati impegnati nella sequenza di reazioni anabolica, differiscono da quelli del processo catabolico ossidativo. Nella B ossidazione il NAD+ e il FAD fungono da accettori di e e il gruppo attivatore è il tiolo (-SH) del coenzima A. L’agente riducente nella via di biosintesi è, invece, il NADPH e i gruppi attivatori sono 2 differenti gruppi –SH legati a due proteine enzimatiche. Nelson/Cox 5th FIGURE 21-2 Addition of two carbons to a growing fatty acyl chain: a four-step sequence. Each malonyl group and acetyl (or longer acyl) group is activated by a thioester that links it to fatty acid synthase, a multienzyme system described later in the text. 1 Condensation of an activated acyl group (an acetyl group from acetyl-CoA is the first acyl group) and two carbons derived from malonyl-CoA, with elimination of CO2 from the malonyl group, extends the acyl chain by two carbons. The mechanism of the first step of this reaction is given to illustrate the role of decarboxylation in facilitating condensation. The β-keto product of this condensation is then reduced in three more steps nearly identical to the reactions of β oxidation, but in the reverse sequence: 2 the β-keto group is reduced to an alcohol, 3 elimination of H2O creates a double bond, and 4 the double bond is reduced to form the corresponding saturated fatty acyl group. KR ER D-β-idrossiacil-ACP
 viene attivato da un tioestere, che lo lega alla acido grasso sintasi. 1- la condensazione di un gruppo acetilico attivato (il gruppo acetilico dell’acetil-CoA fornirà il primo gruppo acilico) e di un frammento a 2 atomi di C derivato dal malonil-CoA, con eliminazione di CO2 dal gruppo malinico, allunga la catena acilica di 2 C. il meccanismo del primo passaggio di questa reazione è qui presentato al fine di illustrare il ruolo della decarbossilazione nei processi di condensazione. Il derivato Β-cheto, prodotto dalla reazione di condensazione viene ridotto mediante altri tre passaggi, quasi identici alle reazioni di B ossidazione, ma in sequenza inversa. 2- il gruppo chetonico B viene ridotto ad alcol. 3- l’eliminazione di H2O crea un doppio legame. 4- il doppio legame viene ridotto, per formare il corrispondente gruppo acilico saturo. Sia il cofattore trasportatore di e, sia i gruppi attivati impegnati nella sequenza di reazioni anabolica, differiscono da quelli del processo catabolico ossidativo. Nella B ossidazione il NAD+ e il FAD fungono da accettori di e e il gruppo attivatore è il tiolo (-SH) del coenzima A. L’agente riducente nella via di biosintesi è, invece, il NADPH e i gruppi attivatori sono 2 differenti gruppi –SH legati a due proteine enzimatiche. Nelson/Cox 5th. FIGURE 21-2 Addition of two carbons to a growing fatty acyl chain: a four-step sequence. Each malonyl group and acetyl (or longer acyl) group is activated by a thioester that links it to fatty acid synthase, a multienzyme system described later in the text. 1 Condensation of an activated acyl group (an acetyl group from acetyl-CoA is the first acyl group) and two carbons derived from malonyl-CoA, with elimination of CO2 from the malonyl group, extends the acyl chain by two carbons. The mechanism of the first step of this reaction is given to illustrate the role of decarboxylation in facilitating condensation. The β-keto product of this condensation is then reduced in three more steps nearly identical to the reactions of β oxidation, but in the reverse sequence: 2 the β-keto group is reduced to an alcohol, 3 elimination of H2O creates a double bond, and 4 the double bond is reduced to form the corresponding saturated fatty acyl group. KR. ER. D-β-idrossiacil-ACP.")
15
H MAT Un gruppo acilico saturo, prodotto da una serie di 4 reazioni, diventa il substrato della successiva condensazione con un gruppo malonico attivato.in ogni passaggio attraverso questa via anabolica, la catena dell’acido grasso si allunga di 2 C. MAT= malonil/acetil-CoA-ACP trasferasi

16
Sequenza degli eventi durante la sintesi di un acido grasso
Nelson e Cox 6°ed. Figura 21.6: Il complesso dell’acido grasso sintasi è mostrato schematicamente, con i domini catalitici colorati. Ciascun dominio del polipeptide rappresenta uno dei sei enzimi del complesso. La proteina trasportatrice di acili è attaccata al dominio KS. Il braccio di fosfopanteteina dell’ACP termina con un –SH. Prima delle reazioni di condensazione che portano alla sintesi dell’acido grasso, i due gruppi tiolici devono essere caricati con i due gruppi acilici corretti. Dapprima il gruppo acetilico dell’acetil.CoA viene trasferito all’ACP.

17
Sintesi del palmitato Nelson e Cox 6°ed. Figura 21.4:
La catena dell’acido grasso cresce di due C per volta, donati dal malonato attivato, con perdita di una molecola di CO2. Il gruppo acetilico iniziale è in giallo; il C1 e il C2 del malonato sono in rosa, e il C rilasciato sotto forma di CO2 in verde. Dopo l’aggiunta di ogni unità bicarboniosa, due reazioni di riduzione convertono la catena nascente in acido grasso saturo con 4, 6, 8 e così via atomi di C. il prodotto finale è il palmitato. Nelson/Cox 3rd The overall process of palmitate synthesis. The fatty acyl chain grows by two-carbon units donated by activated malonate, with loss of CO2 at each step. The initial acetyl group is shaded yellow, C-1 and C-2 of malonate are shaded pink, and the carbon released as CO2 is shaded green. After each two-carbon addition, reductions convert the growing chain to a saturated fatty acid of four, then six, then eight carbons, and so on. The final product is palmitate (16:0).
.")
18
tioesterasi (TE) Per produrre il palmitato saturo a 16 C, sempre legato all’ACP, sono necessari sette cicli di reazioni di condensazione e di riduzione. Per ragione non ancora note, in genere l’allungamento della catena termina a questo punto e il palmitato viene staccato dall’ACP per azione di un tioesterasi (TE), presente nella proteina multifunzionale.
, presente nella proteina multifunzionale.")
19
Richiesta energetica per la biosintesi del palmitato:
8 acetil-CoA 16 ATP (sistema navette per il trasporto dei gruppi acetilici dai mitocondri al citosol, citrato liasi e piruvato carbossilasi) 7 ATP (acetil-CoA carbossilasi) 14 NADPH
 7 ATP (acetil-CoA carbossilasi) 14 NADPH.")
20
Localizzazione subcellulare del metabolismo lipidico

21
Regolazione della sintesi degli acidi grassi
acetil-CoA carbossilasi (monomero) Nelle cellule dei vertebrati il flusso dei precursori del malonil-CoA è regolato sia allostericamente sia mediante modificazione covalente ormone-dipendente. (b) Filamenti di acetil-CoA carbossilasi di epatociti (forma defosforilata ed attiva). Nelson/Cox 5th FIGURE Regulation of fatty acid synthesis. (a) In the cells of vertebrates, both allosteric regulation and hormone-dependent covalent modification influence the flow of precursors into malonyl-CoA. (b) Filaments of acetyl-CoA carboxylase (the active, dephosphorylated form) as seen with the electron microscope.
 Nelle cellule dei vertebrati il flusso dei precursori del malonil-CoA è regolato sia allostericamente sia mediante modificazione covalente ormone-dipendente. (b) Filamenti di acetil-CoA carbossilasi di epatociti (forma defosforilata ed attiva). Nelson/Cox 5th. FIGURE Regulation of fatty acid synthesis. (a) In the cells of vertebrates, both allosteric regulation and hormone-dependent covalent modification influence the flow of precursors into malonyl-CoA. (b) Filaments of acetyl-CoA carboxylase (the active, dephosphorylated form) as seen with the electron microscope.")
22
Regolazione della fosfofruttocinasi-1: modificazione del flusso
degli atomi di C attraverso la glicolisi Nelson/Cox 5th FIGURE 15-14c Phosphofructokinase-1 (PFK-1) and its regulation. (c) Summary of the regulators affecting PFK-1 activity.
 and its regulation. (c) Summary of the regulators affecting PFK-1 activity.")
23
Elongasi mitocondriale:
Elongasi del RE: 2 NADPH + 2 H+ 2 NADP+ palmitoil-CoA stearoil-CoA + CO2 malonil-CoA CoA Elongasi mitocondriale: 2 NAD(P)H + 2 H+ 2 NAD(P)+ Gli acidi a catena lunga sono sintetizzati dal palmitato.Reazioni catalizzate dal sistema di allungamento degli acidi grassi presente nel reticolo endoplasmatico e nei mitocondri. Anche se in questa reazione sono coinvolti enzimi diversi e il trasportatore di acili è il Coenzima A e non l’ACP, il meccanismo di allungamento nell RE è identico a quello usato nella sintesi del palmitato. palmitoil-CoA stearoil-CoA acetil-CoA CoA
H + 2 H+ 2 NAD(P)+ Gli acidi a catena lunga sono sintetizzati dal palmitato.Reazioni catalizzate dal sistema di allungamento degli acidi grassi presente nel reticolo endoplasmatico e nei mitocondri. Anche se in questa reazione sono coinvolti enzimi diversi e il trasportatore di acili è il Coenzima A e non l’ACP, il meccanismo di allungamento nell RE è identico a quello usato nella sintesi del palmitato. palmitoil-CoA. stearoil-CoA. acetil-CoA. CoA.")
24
Desaturasi (ret. endopl.):
D9 desaturasi: indotta da insulina, repressa da PUFA, glucagone e adrenalina Nelson e Cox 6° ed. Figura 21.13: La via di trasferimento degli elettroni nel processo di desaturazione degli acidi grassi. Le frecce blu mostrano come vengono ossidati dall’ossigeno molecolare due substrati diversi: un acil-CoA e il NADPH. Queste reazioni avvengono sulla faccia che guarda verso il lume della membrana del RE. Il doppio legame viene introdotto nella catena dell’acido grasso da una reazione ossidativa catalizzata dalla acil-CoA desaturasi. Questo enzima è un esempio di ossidasi a funzione mista. I due substrati dell’enzima, l’acido grasso e il NADH o NADPH, vanno incontro simultaneamente a ossidazioni che coinvolgono due elettroni: gli e fluiscono all’interno di una via costituita da un citocromo (il citocromo b5) e da una flavoproteina (la citocroma b5 reduttasi), presenti nel RE liscio.
 e da una flavoproteina (la citocroma b5 reduttasi), presenti nel RE liscio.")
25
Ulteriori insaturazioni nell’uomo dopo il D9: D4 desaturasi
N:B: in tutti i passaggi qui indicati l’acido grasso è legato al coenzima A, e non è libero come appare qui, per semplicità di descrizione. Ulteriori insaturazioni nell’uomo dopo il D9: D4 desaturasi D5 desaturasi D6 desaturasi Nelson/Cox 5th FIGURE Routes of synthesis of other fatty acids. Palmitate is the precursor of stearate and longer-chain saturated fatty acids, as well as the monounsaturated acids palmitoleate and oleate. Mammals cannot convert oleate to linoleate or α-linolenate (shaded pink), which are therefore required in the diet as essential fatty acids. Conversion of linoleate to other polyunsaturated fatty acids and eicosanoids is outlined. Unsaturated fatty acids are symbolized by indicating the number of carbons and the number and position of the double bonds, as in Table 10-1. (diomo-γ-linolenic acid)
, which are therefore required in the diet as essential fatty acids. Conversion of linoleate to other polyunsaturated fatty acids and eicosanoids is outlined. Unsaturated fatty acids are symbolized by indicating the number of carbons and the number and position of the double bonds, as in Table (diomo-γ-linolenic acid)")
26
Biosintesi di acidi grassi polinsaturi della serie ω-9:
L’acido oleico, capostipite della serie ω-9, è il costituente più abbondanti della maggioranza degli oli vegetali. Possono essere sintetizzati dall’organismo umane (non essenziali) dall’ acido oleico per successive aggiunte di unità bicarboniose, sotto forma di derivati del CoASH, all’estremità carbossilica. L’acido erucico ha un’azione tossica sul miocardio, e gli oli che lo contengono non possono essere venduti a scopo alimentare. L’acido nervonico è un acido grasso significativamente presente nel tessuto nervoso e nella materia bianca cerebrale.
 dall’ acido oleico per successive aggiunte di unità bicarboniose, sotto forma di derivati del CoASH, all’estremità carbossilica. L’acido erucico ha un’azione tossica sul miocardio, e gli oli che lo contengono non possono essere venduti a scopo alimentare. L’acido nervonico è un acido grasso significativamente presente nel tessuto nervoso e nella materia bianca cerebrale.")
27
Biosintesi di acidi grassi polinsaturi della serie ω-6:
Gli acidi grassi delle serie ω-6 e ω-3 si formano a partire dall’acido linoleico e dall’acido linolenico, rispettivamente. Da questi due acidi grassi (introdotti della dieta) tramite elongasi e desaturasi, si formani gli altri PUFA, componenti indispensabili di tutte le membrane cellulari.
 tramite elongasi e desaturasi, si formani gli altri PUFA, componenti indispensabili di tutte le membrane cellulari.")
28
Biosintesi dell’arachidonato:
L’acido arachidonico (ω-6 ): Biosintesi a livello del fegato. Molto importante perché spesso non sufficientemente fornito dagli alimenti. È costituente importante dei fosfolipidi di membrane e precursore della biosintesi degli eicosanoidi.
: Biosintesi a livello del fegato. Molto importante perché spesso non sufficientemente fornito dagli alimenti. È costituente importante dei fosfolipidi di membrane e precursore della biosintesi degli eicosanoidi.")
29
Formazione degli eicosanoidi:
Agiscono a concentrazioni molto basse e sono coinvolti nella produzione di dolore e febbre e nella regolazione della pressione sanguina, della coagulazione del sangue e della riproduzione.

30
Biosintesi di acidi grassi polinsaturi della serie ω-3:
EPA Nell’uomo l’acido α-linolenico rappresenta il precursore di altri fondamentali acidi grassi della serie ω-3, che vengono anch’essi considerati essenziale dato l’elevato valore biologico. DPA DHA

31
Il giusto rapporto tra omega 6 ed omega 3
Secondo le fonti LARN (Livelli di Assunzione Raccomandata di Nutrienti per la popolazione italiana), nel nostro Paese il rapporto tra omega 6 ed omega 3 è di circa 13:1, che corrisponde ad una percentuale energetica complessiva di circa il 6% del consumo calorico quotidiano. Mentre la razione raccomandata di omega 6 per l'adulto dovrebbe essere l'1-2% delle calorie totali giornaliere, e quella di omega 3 circa lo 0,2-0,5% - la popolazione italiana introduce circa il 5,54% di omega 6 e lo 0,46% di omega 3. Interventi dietetici sono: Riduzione drastica dell'apporto di omega 6 (in particolare dell'acido linoleico) contenuto nei semi di girasole, nel germe di grano, nel sesamo, nelle noci, nei semi di soia, nel mais, nelle olive, quindi nei relativi oli. Aumento significativo dell'apporto di omega 3 (acidi grassi alfa linolenico, eicosapentaenoico e docosaesanoico), le cui fonti alimentari sono: oli e carni del pesce azzurro (ricchi soprattutto in eicosapentaenoico e docosaesanoico), semi di Chia, del kiwi, di Perilla, di lino, di mirtillo rosso; noci e olio di noci, olio di canapa, olio di lino, olio di colza, olio di canola e olio di soia (ricchi soprattutto di alfa-linolenico).
, nel nostro Paese il rapporto tra omega 6 ed omega 3 è di circa 13:1, che corrisponde ad una percentuale energetica complessiva di circa il 6% del consumo calorico quotidiano. Mentre la razione raccomandata di omega 6 per l adulto dovrebbe essere l 1-2% delle calorie totali giornaliere, e quella di omega 3 circa lo 0,2-0,5% - la popolazione italiana introduce circa il 5,54% di omega 6 e lo 0,46% di omega 3. Interventi dietetici sono: Riduzione drastica dell apporto di omega 6 (in particolare dell acido linoleico) contenuto nei semi di girasole, nel germe di grano, nel sesamo, nelle noci, nei semi di soia, nel mais, nelle olive, quindi nei relativi oli. Aumento significativo dell apporto di omega 3 (acidi grassi alfa linolenico, eicosapentaenoico e docosaesanoico), le cui fonti alimentari sono: oli e carni del pesce azzurro (ricchi soprattutto in eicosapentaenoico e docosaesanoico), semi di Chia, del kiwi, di Perilla, di lino, di mirtillo rosso; noci e olio di noci, olio di canapa, olio di lino, olio di colza, olio di canola e olio di soia (ricchi soprattutto di alfa-linolenico).")
33
Rapporto Ω6/Ω3 in alcuni oli di uso comune
Olio di semi di lino 1:4 Olio di colza 2:1 Olio di canapa 3:1 Olio di soia 8:1 Olio di oliva 9:1 Olio di germe di grano 10:1 Olio di arachidi 62:1 Olio di girasole 71:1

34
Perché è determinante correggere il bilancio degli acidi grassi essenziali nel mantenimento dello stato di salute? Il giusto rapporto tra omega 6 ed omega 3 favorisce: l'omeostasi della lipidemia la regolazione della pressione arteriosa GARANTISCE l'equilibrio degli EICOSANOIDI endogeni. Gli omega 3 sono precursori degli eicosanoidi tipo PG1 e PG3, pertanto svolgono una funzione antiaggregante, vasoprotettiva ed antitrombotica; al contrario, gli omega 6 sono anche precursori degli eicosanoidi PG2, che si avvalgono di capacità pro infiammatorie e pro trombotiche. TUTTI gli eicosanoidi (PG1, PG2 e PG3) sono molecole essenziali al corretto funzionamento dell'organismo; tuttavia, la tendenza all'infiammazione CRONICA (potenzialmente aggravata dall'eccesso di omega 6) rappresenta un importante fattore di rischio per le malattie autoimmuni (artrite reumatoide, retto-colite ulcerosa, morbo di Crohn, ecc.) e può aggravare alcune patologie metaboliche pre-esistenti incidendo sul rischio cardiovascolare.
 sono molecole essenziali al corretto funzionamento dell organismo; tuttavia, la tendenza all infiammazione CRONICA (potenzialmente aggravata dall eccesso di omega 6) rappresenta un importante fattore di rischio per le malattie autoimmuni (artrite reumatoide, retto-colite ulcerosa, morbo di Crohn, ecc.) e può aggravare alcune patologie metaboliche pre-esistenti incidendo sul rischio cardiovascolare.")
35
Biosintesi dei triacilgliceroli:
Nelson e Cox 6° ed. Figura 21.17: La via biosintetica del fosfatidato parte 1 Un gruppo acilico viene prima attivato mediante la formazione di acil-CoA e poi trasferito attraverso un legame estere all’L-glicerolo 3-fosfato formato in una delle vie indicate. Nelson/Cox 5th ed. FIGURE (part 1) Biosynthesis of phosphatidic acid. A fatty acyl group is activated by formation of the fatty acyl-CoA, then transferred to ester linkage with L-glycerol 3-phosphate, formed in either of the two ways shown. Phosphatidic acid is shown here with the correct stereochemistry at C-2 of the glycerol molecule. To conserve space in subsequent figures (and in Figure 21-14), both fatty acyl groups of glycerophospholipids, and all three acyl groups of triacylglycerols, are shown projecting to the right.
 Biosynthesis of phosphatidic acid. A fatty acyl group is activated by formation of the fatty acyl-CoA, then transferred to ester linkage with L-glycerol 3-phosphate, formed in either of the two ways shown. Phosphatidic acid is shown here with the correct stereochemistry at C-2 of the glycerol molecule. To conserve space in subsequent figures (and in Figure 21-14), both fatty acyl groups of glycerophospholipids, and all three acyl groups of triacylglycerols, are shown projecting to the right.")
36
lipoproteins (chylomicrons, VLDL)
endogenous synthesis NEFA-albumin lipoproteins (chylomicrons, VLDL) insulin glucagon Nelson/Cox 5th ed. FIGURE (part 2) Biosynthesis of phosphatidic acid. A fatty acyl group is activated by formation of the fatty acyl-CoA, then transferred to ester linkage with L-glycerol 3-phosphate, formed in either of the two ways shown. Phosphatidic acid is shown here with the correct stereochemistry at C-2 of the glycerol molecule. To conserve space in subsequent figures (and in Figure 21-14), both fatty acyl groups of glycerophospholipids, and all three acyl groups of triacylglycerols, are shown projecting to the right.
 insulin. glucagon. Nelson/Cox 5th ed. FIGURE (part 2) Biosynthesis of phosphatidic acid. A fatty acyl group is activated by formation of the fatty acyl-CoA, then transferred to ester linkage with L-glycerol 3-phosphate, formed in either of the two ways shown. Phosphatidic acid is shown here with the correct stereochemistry at C-2 of the glycerol molecule. To conserve space in subsequent figures (and in Figure 21-14), both fatty acyl groups of glycerophospholipids, and all three acyl groups of triacylglycerols, are shown projecting to the right.")
37
Nelson e Cox 6°ed. Figura 21.18: L’acido fosfatidico nella biosintesi lipidica.
L’acido fosfatidico è il precursore sia dei triacilgliceroli sia dei glicerofosfolipidi Nelson/Cox 5th ed. mod. FIGURE Phosphatidic acid in lipid biosynthesis. Phosphatidic acid is the precursor of both triacylglycerols and glycerophospholipids. The mechanisms for head-group attachment in phospholipid synthesis are described later in this section.

38
Nelson/Cox 5th FIGURE Regulation of triacylglycerol synthesis by insulin. Insulin stimulates conversion of dietary carbohydrates and proteins to fat. Individuals with diabetes mellitus lack insulin; in uncontrolled disease, this results in diminished fatty acid synthesis, and the acetyl-CoA arising from catabolism of carbohydrates and proteins is shunted instead to ketone body production. People in severe ketosis smell of acetone, so the condition is sometimes mistaken for drunkenness (p. 929).
.")
39
VLDL albumin Nelson e Cox 6° ed. Figura 21.20: Il ciclo del triacilglicerolo Nei mammiferi, durante il digiuno, i triacilgliceroli sono prima degradati e poi nuovamente sintetizzate in un ciclo del triacilglicerolo. Parte dei acidi grassi rilasciati dalla lipolisi dei triacilgliceroli nel tessuto adiposo passa nel sangue, mentre la parte restante viene impiegata per essere di nuovo sintetizzata come triacilgliceroli. Parte degli acidi grassi rilasciati nel sangueviene utilizzata come fonte di energia (per esempio nel muscolo) e parte viene assorbita dal fegato per la sintesi di triacilgliceroli. I triacilgliceroli formati nel fegato passano nel sangue e sono trasportati al tessuto adiposo dove una lipoproteina lipasi extracellulare rilascia gli acidi grassi. Gli adipociti assorbono gli acidi grassi e li riesterificano a triacilgliceroli. Nelson/Cox 5° FIGURE The triacylglycerol cycle. In mammals, triacylglycerol molecules are broken down and resynthesized in a triacylglycerol cycle during starvation. Some of the fatty acids released by lipolysis of triacylglycerol in adipose tissue pass into the bloodstream, and the remainder are used for resynthesis of triacylglycerol. Some of the fatty acids released into the blood are used for energy (in muscle, for example), and some are taken up by the liver and used in triacylglycerol synthesis. The triacylglycerol formed in the liver is transported in the blood back to adipose tissue, where the fatty acid is released by extracellular lipoprotein lipase, taken up by adipocytes, and reesterified into triacylglycerol. DAP DAP triacylglycerol cycle: continuous recycling in both hepatocyte and adipocyte, both after a meal and during fasting
 e parte viene assorbita dal fegato per la sintesi di triacilgliceroli. I triacilgliceroli formati nel fegato passano nel sangue e sono trasportati al tessuto adiposo dove una lipoproteina lipasi extracellulare rilascia gli acidi grassi. Gli adipociti assorbono gli acidi grassi e li riesterificano a triacilgliceroli. Nelson/Cox 5° FIGURE The triacylglycerol cycle. In mammals, triacylglycerol molecules are broken down and resynthesized in a triacylglycerol cycle during starvation. Some of the fatty acids released by lipolysis of triacylglycerol in adipose tissue pass into the bloodstream, and the remainder are used for resynthesis of triacylglycerol. Some of the fatty acids released into the blood are used for energy (in muscle, for example), and some are taken up by the liver and used in triacylglycerol synthesis. The triacylglycerol formed in the liver is transported in the blood back to adipose tissue, where the fatty acid is released by extracellular lipoprotein lipase, taken up by adipocytes, and reesterified into triacylglycerol. DAP. DAP. triacylglycerol cycle: continuous recycling in both hepatocyte and adipocyte, both after a meal and during fasting.")
40
Glyceroneogenesis: (adipocyte and hepatocyte)
Nelson e Cox 6° ed. Figura 21.21: Gliceroneogenesi La via è essenzialmente une versione abbreviata della gluconeogenesi, dal piruvato al diidrossiacetato fosfato (DHAP), a cui fa seguito la conversione del DHAP in glicerolo 3-fosfato (per azione della glicerolo 3-fosfato deidrogenasi NAD-dipendente del citosol), che viene usato per la sintesi dei triacilgliceroli. Nelson/Cox 5th FIGURE Glyceroneogenesis. The pathway is essentially an abbreviated version of gluconeogenesis, from pyruvate to dihydroxyacetone phosphate (DHAP), followed by conversion of DHAP to glycerol 3-phosphate, which is used for the synthesis of triacylglycerol.
, a cui fa seguito la conversione del DHAP in glicerolo 3-fosfato (per azione della glicerolo 3-fosfato deidrogenasi NAD-dipendente del citosol), che viene usato per la sintesi dei triacilgliceroli. Nelson/Cox 5th. FIGURE Glyceroneogenesis. The pathway is essentially an abbreviated version of gluconeogenesis, from pyruvate to dihydroxyacetone phosphate (DHAP), followed by conversion of DHAP to glycerol 3-phosphate, which is used for the synthesis of triacylglycerol.")
41
Randle Cycle Increased fatty acid oxidation inhibits glucose oxidation via the Randle Cycle. In 1963, Randle first described the glucose-fatty acid cycle whereby the products of fatty acid oxidation limit glucose utilization. Briefly, acetyl-CoA generated from fatty acid oxidation inhibits pyruvate dehydrogenase (PDH) both allosterically and by activating PDH kinases that phosphorylate and inhibit the enzyme. In addition, citrate generated from the Krebs cycle and exported to the cytosol inhibits phosphofructiokinase 1 (PFK-1) reducing glycolytic flux and the production of pyruvate. Thus conditions that enhance fatty acid oxidation, such as high dietary fat, obesity, and starvation, limit the generation and oxidation of pyruvate. Conversely, acetyl-CoA generated from citrate provides substrate for acetyl-CoA cocarboxylase (ACC), thereby generating malonyl-CoA that inhibits mitochondrial fatty acid uptake and oxidation. CPT1, carnitine palmitoyltransferase 1; MCD, malonyl-CoA decarboxylase.
 both allosterically and by activating PDH kinases that phosphorylate and inhibit the enzyme. In addition, citrate generated from the Krebs cycle and exported to the cytosol inhibits phosphofructiokinase 1 (PFK-1) reducing glycolytic flux and the production of pyruvate. Thus conditions that enhance fatty acid oxidation, such as high dietary fat, obesity, and starvation, limit the generation and oxidation of pyruvate. Conversely, acetyl-CoA generated from citrate provides substrate for acetyl-CoA cocarboxylase (ACC), thereby generating malonyl-CoA that inhibits mitochondrial fatty acid uptake and oxidation. CPT1, carnitine palmitoyltransferase 1; MCD, malonyl-CoA decarboxylase.")
42
Legame della testa polare dei glicerofosfolipidi
Nelson e Cox 6° ed. Figura 21.23: Legame della testa polare La testa polare dei fosfolipidi viene legata al diacilgliceroli con un ponte fosfodiestere che si forma quando l’acido fosforico si condensa con due alcoli, eliminando due molecole d’acqua. La testa polare dei glicerofosfolipidi viene legata tramite un legame fosfodiestere, in cui ciascuno dei due ossidrili alcolici (uno sulla testa polare e l’altro sul C-3 del glicerolo) forma un estere con l’acido fosforico.
 forma un estere con l’acido fosforico.")
43
phosphatidic acid pathway salvage pathway
Le cellule utilizzano due strategie per legare le teste polari ai fosfolipidi. Uno degli ossidrili viene prima attivato legandosi a un nucleotide, la citidina difosfato (CDP). La citidina monofosfato (CMP) viene poi rilasciata in seguito a un attaco nucleofilico da parte dell’altro ossidrile. Strategia 1: il CDP si lega al diacilglicerolo, generando una forma attivata dell’acido fosfatidico, il CDP-diacilglicerolo. Strategia 2: il CDP si lega al gruppo ossidrilico della testa polare. Nelson/Cox 5th FIGURE Two general strategies for forming the phosphodiester bond of phospholipids. In both cases, CDP supplies the phosphate group of the phosphodiester bond.
. La citidina monofosfato (CMP) viene poi rilasciata in seguito a un attaco nucleofilico da parte dell’altro ossidrile. Strategia 1: il CDP si lega al diacilglicerolo, generando una forma attivata dell’acido fosfatidico, il CDP-diacilglicerolo. Strategia 2: il CDP si lega al gruppo ossidrilico della testa polare. Nelson/Cox 5th. FIGURE Two general strategies for forming the phosphodiester bond of phospholipids. In both cases, CDP supplies the phosphate group of the phosphodiester bond.")
44
fosfatidilcolina (1): (apporto adeguato di colina: circa 500 mg)
Formazione della fosfatidilcolina: La sintesi della fosfatidilcolina nei mammiferi avviene secondo la strategia 2: fosforilazione e attivazione della testa polare, seguita dalla condensazione col diacilgliceroli. La colina viene riusata («salvata») tramite una fosforilazione,…
 tramite una fosforilazione,…")
45
fosfatidilcolina (2): …, poi convertita in CDP colina grazie alla condensazione col CTP.
: …, poi convertita in CDP colina grazie alla condensazione col CTP.")
46
fosfatidilcolina (3): Una molecola di diacilglicerolo spiazza il CMP dalla CDP-colina, producendo la fosfatidilcolina.
: Una molecola di diacilglicerolo spiazza il CMP dalla CDP-colina, producendo la fosfatidilcolina.")
47
fosfatidiletanolamina (1):
HO CH CH NH3 2 2 etanolamina ATP colina cinasi ADP L’etanolammina ottenuta dalla dieta o quella presente nell’organismo viene riusata («salvata») tramite una fosforilazione,… O O P O CH CH NH3 2 2 O fosfoetanolamina
 tramite una fosforilazione,… O. O. P. O. CH. CH. NH O. fosfoetanolamina.")
48
fosfatidiletanolamina (2):
P O CH CH NH3 2 2 O fosfoetanolamina CTP CTP etanolamina citidiltransferasi PP i citosina …, poi convertita in CDP etanolamina grazie alla condensazione col CTP. O O riboso O P O P O CH CH NH3 2 2 O O citidindifosfoetanolamina (CDPetanolamina)
")
49
fosfatidiletanolamina (3):
citosina O O riboso O P O P O CH CH NH3 2 2 O O fosfatidiletanolamina (3): citidindifosfoetanolamina (CDPetanolamina) diacilglicerolo fosfoetanolamina transferasi CMP Una molecola di diacilglicerolo spiazza il CMP dalla CDP-etanolammina, producendo la fosfatidiletanolammina. O O H C 2 O C C O C H O H C O P O CH CH NH3 2 2 2 O fosfatidiletanolamina
: citidindifosfoetanolamina (CDPetanolamina) diacilglicerolo. fosfoetanolamina. transferasi. CMP. Una molecola di diacilglicerolo spiazza il CMP dalla CDP-etanolammina, producendo la fosfatidiletanolammina. O. O. H C. 2. O. C. C. O. C. H. O. H C. O. P. O. CH. CH. NH O. fosfatidiletanolamina.")
50
PSS = phosphatidylserine synthase
La fosfatidilserina è sintetizzata mediante reazioni di scambio Ca2+-dipendenti promosse dalla fosfatidilserina sintasi 1 (PSS1) o dalla fosfatidilserina sintasi 2 (PSS2). La PSS1 può usare come substrati la fosfatidiletanolammina o la fosfatidilcolina. Nelson/Cox 5th FIGURE 21-28a Pathways for phosphatidylserine and phosphatidylcholine synthesis in mammals. (a) Phosphatidylserine is synthesized by Ca2+-dependent head-group exchange reactions promoted by phosphatidylserine synthase 1 (PSS1) or phosphatidylserine synthase 2 (PSS2). PSS1 can use either phosphatidylethanolamine or phosphatidylcholine as a substrate. PSS = phosphatidylserine synthase
 o dalla fosfatidilserina sintasi 2 (PSS2). La PSS1 può usare come substrati la fosfatidiletanolammina o la fosfatidilcolina. Nelson/Cox 5th. FIGURE 21-28a Pathways for phosphatidylserine and phosphatidylcholine synthesis in mammals. (a) Phosphatidylserine is synthesized by Ca2+-dependent head-group exchange reactions promoted by phosphatidylserine synthase 1 (PSS1) or phosphatidylserine synthase 2 (PSS2). PSS1 can use either phosphatidylethanolamine or phosphatidylcholine as a substrate. PSS = phosphatidylserine synthase.")
51
Nei eucarioti la fosfatidiletanolammina può essere sintetizzata dalla fosfatidilserina in una reazione catalizzata dalla fosfatidilserina decarbossilasi. Nelson/Cox 5th FIGURE (part 1) The major path from phosphatidylserine to phosphatidylethanolamine and phosphatidylcholine in all eukaryotes. AdoMet is S-adenosylmethionine; adoHcy, S-adenosylhomocysteine.
 The major path from phosphatidylserine to phosphatidylethanolamine and phosphatidylcholine in all eukaryotes. AdoMet is S-adenosylmethionine; adoHcy, S-adenosylhomocysteine.")
52
liver only (S-adenosilmetionina) (lecitina)
Nei eucarioti, la fosfatidiletanolammina può essere convertita in fosfatidilcolina (lecitina) per aggiunta di tre gruppi metilici al suo gruppo amminico; il donatore dei gruppi amminici è la S-adenosilmetionina per tutte e tre le reazioni di metilazione. Nelson/Cox 5th FIGURE (part 2) The major path from phosphatidylserine to phosphatidylethanolamine and phosphatidylcholine in all eukaryotes. AdoMet is S-adenosylmethionine; adoHcy, S-adenosylhomocysteine. (lecitina)
 per aggiunta di tre gruppi metilici al suo gruppo amminico; il donatore dei gruppi amminici è la S-adenosilmetionina per tutte e tre le reazioni di metilazione. Nelson/Cox 5th. FIGURE (part 2) The major path from phosphatidylserine to phosphatidylethanolamine and phosphatidylcholine in all eukaryotes. AdoMet is S-adenosylmethionine; adoHcy, S-adenosylhomocysteine. (lecitina)")
53
Sintesi fosfatidilinositolo:
inositolo-1-P glucoso-6-P Il fosfatidilinositolo è sintetizzato usando la strategia 1: per condensazione del CDP-diacilglicerolo con l’inositolo. Specifiche fosfatidilinosotolo chinasi poi convertono il fosfatidilinositolo nei suoi derivati fosforilati. Il fosfatidilinositolo e i suoi derivati fosforilati svolgono un ruolo centrale nel processo di trasduzione dei segnali negli eucarioti. Nelson/Cox 5th FIGURE Synthesis of cardiolipin and phosphatidylinositol in eukaryotes. These glycerophospholipids are synthesized using strategy 1 in Figure Phosphatidylglycerol is synthesized as in bacteria (see Figure 21-25). PI represents phosphatidylinositol.
. PI represents phosphatidylinositol.")
54
Sintesi fosfatidilglicerolo:
Il diacilglicerolo viene attivato per condensazione dell’acido fosfatidico col CTP (strategia 1), con eliminazione del pirofosfato e formazione del CDP-diacilglicerolo. Il distacco del CMP a seguito dell’attaco nucleofilico del gruppo ossidrilico in C1 del glicerolo 3-fosfato produce il fosfatidilglicerolo 3-fosfato. Quest’ultimo viene ulteriormente modificato, per scissione del monoestere fosforico (con rilascio di Pi), per produrre il fosfatidilglicerolo. Nelson/Cox 5th mod. FIGURE Origin of the polar head groups of phospholipids in E. coli. Initially, a head group (either serine or glycerol 3-phosphate) is attached via a CDP-diacylglycerol intermediate (strategy 1 in Figure 21-24). For phospholipids other than phosphatidylserine, the head group is further modified, as shown here. In the enzyme names, PG represents phosphatidylglycerol; PS, phosphatidylserine.
, con eliminazione del pirofosfato e formazione del CDP-diacilglicerolo. Il distacco del CMP a seguito dell’attaco nucleofilico del gruppo ossidrilico in C1 del glicerolo 3-fosfato produce il fosfatidilglicerolo 3-fosfato. Quest’ultimo viene ulteriormente modificato, per scissione del monoestere fosforico (con rilascio di Pi), per produrre il fosfatidilglicerolo. Nelson/Cox 5th mod. FIGURE Origin of the polar head groups of phospholipids in E. coli. Initially, a head group (either serine or glycerol 3-phosphate) is attached via a CDP-diacylglycerol intermediate (strategy 1 in Figure 21-24). For phospholipids other than phosphatidylserine, the head group is further modified, as shown here. In the enzyme names, PG represents phosphatidylglycerol; PS, phosphatidylserine.")
55
Sintesi cardiolipina:
Negli eucarioti, il diacilglicerolo viene attivato per condensazione dell’acido fosfatidico col CTP (strategia 1), poi il fosfatidilglicerolo condensa con il CDP-diacilglicerolo formendo la cardiolipina. Nelson/Cox 5th FIGURE Synthesis of cardiolipin and phosphatidylinositol in eukaryotes. These glycerophospholipids are synthesized using strategy 1 in Figure Phosphatidylglycerol is synthesized as in bacteria (see Figure 21-25). PI represents phosphatidylinositol.
, poi il fosfatidilglicerolo condensa con il CDP-diacilglicerolo formendo la cardiolipina. Nelson/Cox 5th. FIGURE Synthesis of cardiolipin and phosphatidylinositol in eukaryotes. These glycerophospholipids are synthesized using strategy 1 in Figure Phosphatidylglycerol is synthesized as in bacteria (see Figure 21-25). PI represents phosphatidylinositol.")
56
Fosfatidilglicerolo e cardiolipina si trovano nella membrana dei batteri e nella membrana mitocondriale interna degli eucarioti. Il fosfatidilglicerolo si trova anche nel surfattante polmonare, insieme alla fosfatidilcolina e alla dipalmitoillecitina. Champe La cardiolipina è un fosfogliceride umano riconosciuto come antigene dal T. pallidum (test di Wassermann per la diagnosi di sifilide).
.")
57
Champe

58
Champe

59
rimaneggiamento: Rimaneggiamento della fosfatidilcolina dall’azione delle diverse fosfolipasi.

60
rimaneggiamento: Champe

61
synthesis in peroxisomes
La sintesi dei plasmalogeni richiede la formazione di legami etere con alcoli a catena lunga. I lipidi con legami etere, come i plasmalogeni e il fattore di attivazione delle piastrine, sono sintetizzati mediante sostituzione di un gruppo acilico, impegnato con legame estere, con un alcol a catena lunga unito con legame etere. Nelson/Cox 5th FIGURE 10-10a Ether lipids. Plasmalogens have an ether-linked alkenyl chain where most glycerophospholipids have an ester-linked fatty acid (compare Figure 10-9). Platelet-activating factor has a long ether-linked alkyl chain at C-1 of glycerol, but C-2 is ester-linked to acetic acid, which makes the compound much more water-soluble than most glycerophospholipids and plasmalogens. The head-group alcohol is ethanolamine in plasmalogens and choline in platelet-activating factor. synthesis in peroxisomes
. Platelet-activating factor has a long ether-linked alkyl chain at C-1 of glycerol, but C-2 is ester-linked to acetic acid, which makes the compound much more water-soluble than most glycerophospholipids and plasmalogens. The head-group alcohol is ethanolamine in plasmalogens and choline in platelet-activating factor. synthesis in peroxisomes.")
62
Nelson/Cox 5th FIGURE 10-10b Ether lipids. Plasmalogens have an ether-linked alkenyl chain where most glycerophospholipids have an ester-linked fatty acid (compare Figure 10-9). Platelet-activating factor has a long ether-linked alkyl chain at C-1 of glycerol, but C-2 is ester-linked to acetic acid, which makes the compound much more water-soluble than most glycerophospholipids and plasmalogens. The head-group alcohol is ethanolamine in plasmalogens and choline in platelet-activating factor.
. Platelet-activating factor has a long ether-linked alkyl chain at C-1 of glycerol, but C-2 is ester-linked to acetic acid, which makes the compound much more water-soluble than most glycerophospholipids and plasmalogens. The head-group alcohol is ethanolamine in plasmalogens and choline in platelet-activating factor.")
63
PAF: stimolazione dell'aggregazione piastrinica vasodilatazione e vasopermeabilizzazione riduzione della pressione arteriosa e della gittata cardiaca attivazione leucocitaria e stimolo alla chemiotassi leucocitaria stimolazione della contrazione della muscolatura liscia uterina, bronchiale e intestinale ruolo importante nelle reazioni di ipersensibilità, nelle reazioni infiammatorie acute e nello shock anafilattico

64
Biosintesi degli sfingolipidi
Nelson e Cox 6° ed. Figura 21.31: La condensazione del palmitil-CoA con la serina (che forma la B-chetosfinganina), seguita da riduzione con NADPH, produce sfinganina, che viene poi acilata per formare N-acilsfinganina (un ceramide). Negli animali, un’ossidasi a funzione mista forma un doppio legame (ombraggiato in rosa) prima dell’aggiunta di una testa polare: la fosfatidilcolina per formare la sfingomielina o il glucosio per formare un cerebroside.
, seguita da riduzione con NADPH, produce sfinganina, che viene poi acilata per formare N-acilsfinganina (un ceramide). Negli animali, un’ossidasi a funzione mista forma un doppio legame (ombraggiato in rosa) prima dell’aggiunta di una testa polare: la fosfatidilcolina per formare la sfingomielina o il glucosio per formare un cerebroside.")
65
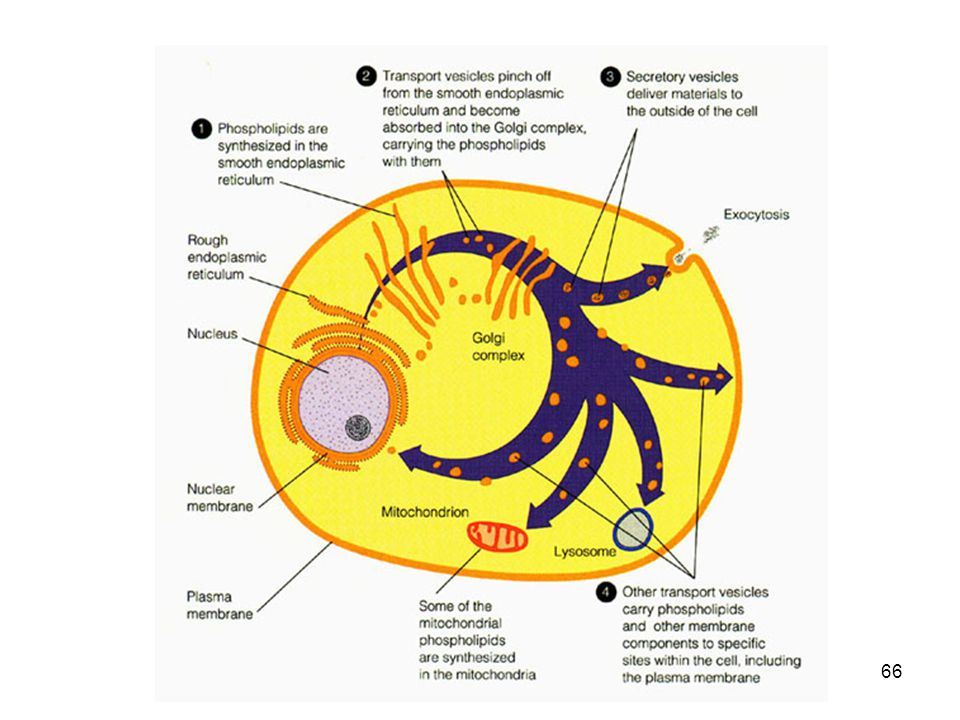
Champe Dopo la sintesi nel RE liscio, i lipidi polari, compresi i glicerofosfolipidi, gli sfingolipidi e i glicolipidi, sono inseriti in diversi proporzioni della membrane cellulari attraverso meccanismi non ancora noti.

67
Lipid storage diseases or sphingolipidoses:

Presentazioni simili