Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Le membrane biologiche

2
Le membrane biologiche

3
Tipica membrana biologica

4
Trasporto di soluti attraverso le membrane……..

5
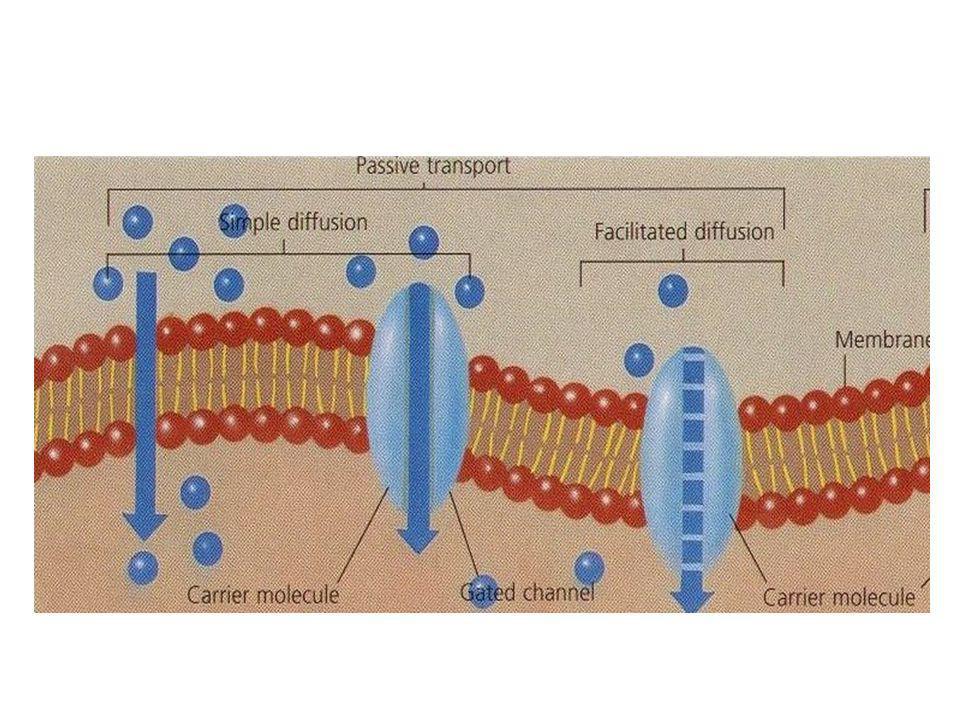
Meccanismi di trasporto
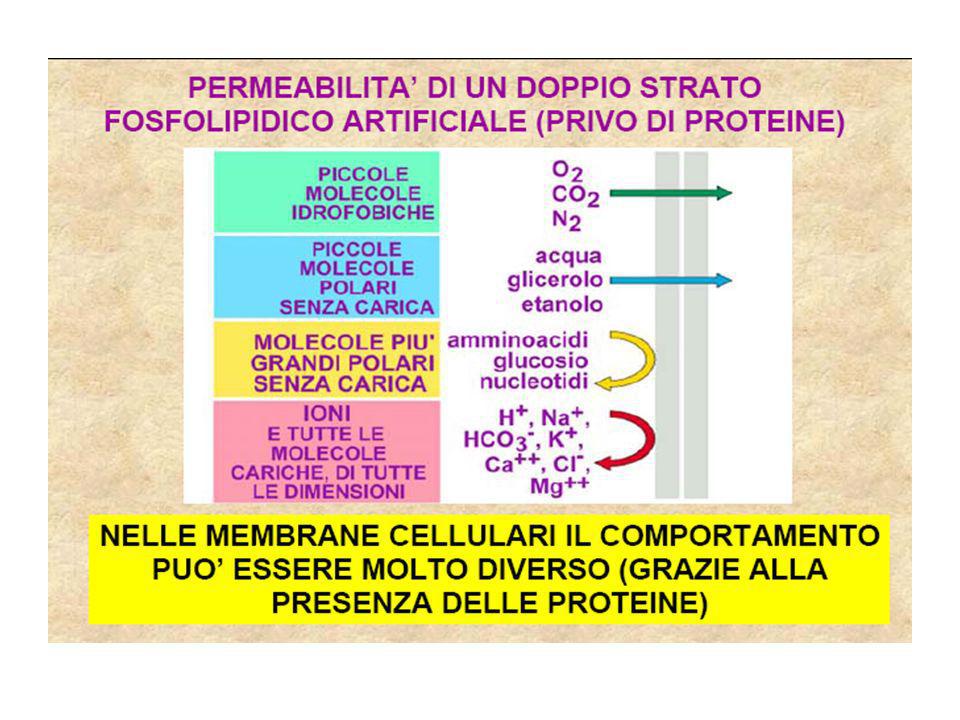
La membrana cellulare è in grado di permettere il passaggio di sostanze in modo selettivo grazie a meccanismi di trasporto. Ricordiamo che il doppio strato lipidico costituisce una barriera al passaggio di tutte le molecole polari cariche, mentre permette il passaggio di piccole molecole polari prive di carica. Le sostanze idrofobiche passano invece bene il doppio strato senza causare problemi (O2, CO2, N2).
.")
7
Movimento di molecole attraverso le membrane: la diffusione semplice
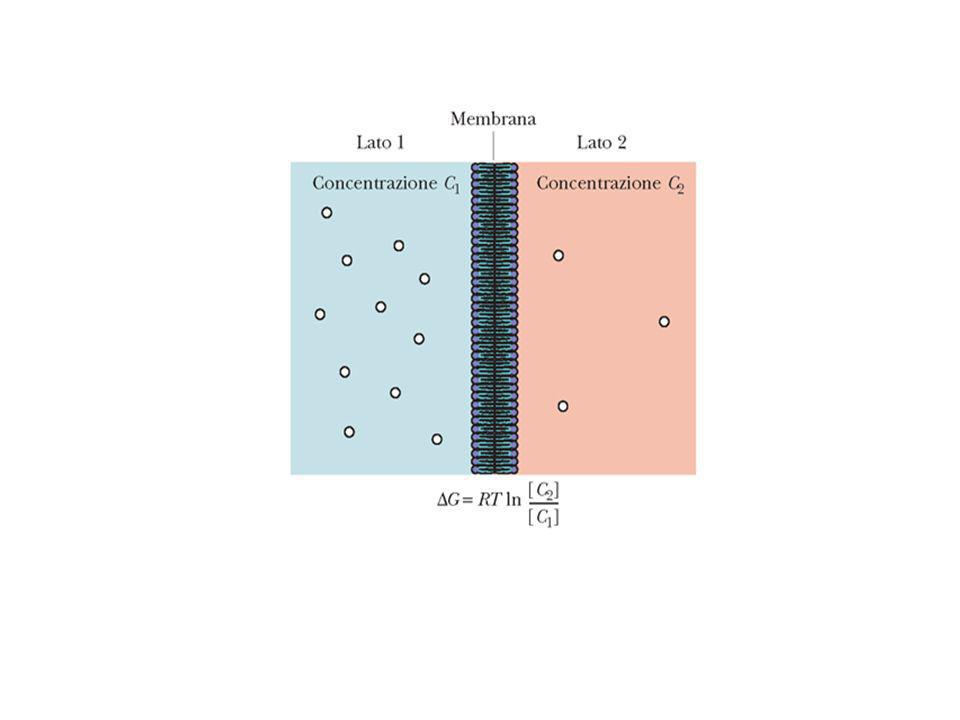
Permeabilità dei doppi strati lipidici Nella diffusione semplice se una molecola di soluto viene in contatto con la fase lipidica della membrana e la sua energia termica è sufficientemente elevata, essa può entrarvi e oltrepassarla. Per poter abbandonare la fase acquosa ed entrare nella fase lipidica, un soluto deve prima di tutto rompere i legami idrogeno con l’acqua (per la rottura di un legame idrogeno si richiedono 5 kcal/mol). Inoltre la molecola di soluto deve essere solubile nella fase lipidica ed il suo grado di solubilità sarà determinante per l’attraversamento della membrana. Quindi tutte le sostanze che formano pochi legami idrogeno con l’acqua possono entrare più facilmente nel doppio strato lipidico mentre gli ioni inorganici non avranno quasi mai la possibilità di farlo. In genere molecole piccole e non polari come Ossigeno (32 dalton) e anidride carbonica (44 dalton) e molecole fortemente idrofobiche (o lipofiliche) diffondono rapidamente attraverso i doppi strati lipidici. Piccole molecole polari come acqua (18 dalton), etanolo (46 dalton) e urea (60 dalton) attraversano rapidamente; il glicerolo (92 dalton) diffonde meno rapidamente mentre il glucosio, che è una grossa molecola polare, praticamente non diffonde. I doppi strati lipidici sono praticamente impermeabili a molecole cariche e a ioni per quanto piccole esse siano; la carica e l’alto grado di idratazione di queste molecole ne impedisce l’ingresso nella fase idrocarburica del doppio strato. In base a queste considerazioni sui doppi strati lipidici artificiale, si può ipotizzare che il grado di diffusione di una molecola attraverso una membrana, ovvero la sua permeabilità relativa, dipenderà dal suo coefficiente di ripartizione olio/acqua (K). Questa ipotesi venne sottoposta a verifica da Collander nel Egli utilizzando la cellula dell’alga gigante Chara ottenne un grafico in cui la permeabilità relativa delle molecole era riportata in funzione del coefficiente di ripartizione olio-acqua. Salvo poche eccezione, il grafico dimostra una correlazione quasi lineare tra permeabilità e coefficiente di ripartizione olio/acqua, indipendentemente dal peso molecolare. La permeabilità dell’acqua attraverso le membrane cellulari è molto maggiore rispetto a quanto si possa dedurre dal suo coefficiente di ripartizione. Ciò è dovuto al fatto che l’acqua può passare attraverso canali selettivi che penetrano il doppio strato lipidico. Canali denominati acquaporine sono stati riscontrati in molti epiteli e sono responsabili della permeabilità dell’acqua a quel livello. Anche nei doppi strati lipidici artificiali, privi di canali, la permeabilità all’acqua è superiore a quanto ci si dovrebbe aspettare in base alla sua solubilità nella fase idrocarburica. La spiegazione risiede nel passaggio di acqua o di altre piccole molecole polari non cariche attraverso dei canali temporanei che si formano tra le molecole lipidiche. In ogni casi la velocità di diffusione di una sostanza attraverso il nucleo idrofobico di una membrana fosfolipidica è 100 – 1000 volte inferiore alla velocità di diffusione della stessa molecola in acqua dato che la membrana cellulare è 100 – 1000 volte più viscosa dell’acqua.
. Inoltre la molecola di soluto deve essere solubile nella fase lipidica ed il suo grado di solubilità sarà determinante per l’attraversamento della membrana. Quindi tutte le sostanze che formano pochi legami idrogeno con l’acqua possono entrare più facilmente nel doppio strato lipidico mentre gli ioni inorganici non avranno quasi mai la possibilità di farlo. In genere molecole piccole e non polari come Ossigeno (32 dalton) e anidride carbonica (44 dalton) e molecole fortemente idrofobiche (o lipofiliche) diffondono rapidamente attraverso i doppi strati lipidici. Piccole molecole polari come acqua (18 dalton), etanolo (46 dalton) e urea (60 dalton) attraversano rapidamente; il glicerolo (92 dalton) diffonde meno rapidamente mentre il glucosio, che è una grossa molecola polare, praticamente non diffonde. I doppi strati lipidici sono praticamente impermeabili a molecole cariche e a ioni per quanto piccole esse siano; la carica e l’alto grado di idratazione di queste molecole ne impedisce l’ingresso nella fase idrocarburica del doppio strato. In base a queste considerazioni sui doppi strati lipidici artificiale, si può ipotizzare che il grado di diffusione di una molecola attraverso una membrana, ovvero la sua permeabilità relativa, dipenderà dal suo coefficiente di ripartizione olio/acqua (K). Questa ipotesi venne sottoposta a verifica da Collander nel Egli utilizzando la cellula dell’alga gigante Chara ottenne un grafico in cui la permeabilità relativa delle molecole era riportata in funzione del coefficiente di ripartizione olio-acqua. Salvo poche eccezione, il grafico dimostra una correlazione quasi lineare tra permeabilità e coefficiente di ripartizione olio/acqua, indipendentemente dal peso molecolare. La permeabilità dell’acqua attraverso le membrane cellulari è molto maggiore rispetto a quanto si possa dedurre dal suo coefficiente di ripartizione. Ciò è dovuto al fatto che l’acqua può passare attraverso canali selettivi che penetrano il doppio strato lipidico. Canali denominati acquaporine sono stati riscontrati in molti epiteli e sono responsabili della permeabilità dell’acqua a quel livello. Anche nei doppi strati lipidici artificiali, privi di canali, la permeabilità all’acqua è superiore a quanto ci si dovrebbe aspettare in base alla sua solubilità nella fase idrocarburica. La spiegazione risiede nel passaggio di acqua o di altre piccole molecole polari non cariche attraverso dei canali temporanei che si formano tra le molecole lipidiche. In ogni casi la velocità di diffusione di una sostanza attraverso il nucleo idrofobico di una membrana fosfolipidica è 100 – 1000 volte inferiore alla velocità di diffusione della stessa molecola in acqua dato che la membrana cellulare è 100 – 1000 volte più viscosa dell’acqua.")
11
Canali di membrana e pori
Canali di tipo alfa: . hanno da 2 a 22 segmenti alfa elica di membrana: Nell’uomo ne sono presenti 15 tipi e facilitano il passaggio di ioni e piccole molecole in favore di gradiente. Una famiglia è quella delle acquaporine che facilita l’ingresso dell’acqua.Ne sono state individuate 11 famiglie Il trasporto attraverso i canali è regolato in vari modi: cambiamenti di potenziale ( canali voltaggio dipendenti) o da molecole modulatrici (ligandi) o da AMPc Canali di tipo beta: è una struttura proteica di tipo beta che forma una struttura a barile intorno all’acqua. Ha un ampio range di selezione del substrato lasciandosi attraversare sia da ione che da proteine e dipende dalla dimensione del poro che forma. La differenza tra canali e pori sta nelle dimensioni dello spazio acquoso che si forma quando è attraversata dai composti
 o da molecole modulatrici (ligandi) o da AMPc. Canali di tipo beta: è una struttura proteica di tipo beta che forma una struttura a barile intorno all’acqua. Ha un ampio range di selezione del substrato lasciandosi attraversare sia da ione che da proteine e dipende dalla dimensione del poro che forma. La differenza tra canali e pori sta nelle dimensioni dello spazio acquoso che si forma quando è attraversata dai composti.")
12
Ionofori Ionofori trasportatori:aumentano la permeabiltà della membrana agli ioni ,diffondendo attraverso la membrana e liberando lo ione all’nterno. Es. la valinomicina trasporta ioni K+ attraverso la membrana con una velocità di 10/4 ioni al secondo:è’ un polipetide ciclico a struttura ottaedrica che coordina al suo interno uno ione K+. Lo ione K* si inserisce perfettamente nel sito di coordinazione ,ma non lo ione Na*,più piccolo. Ionofori formanti canali o pori transmembrana: La gramicidina A polipeptide costituito da 15 aa idrofobici che dimerizzando testa a testa forma un poro che facilita il passaggio di ioni Na+ e K+ di un fattore di 10/7.

13
Diffusione facilitata 1
Aggancio della molecola alla proteina vettrice

14
Diffusione facilitata 2
Liberazione della molecola nel citoplasma Cinetica di saturazione

15
Cinetica di saturazione

19
Trasporto del glucosio

20
Trasporto del glucosio

21
GLUcose Transporters (GLUT)
GLUT1,2,3,5: costituzionale GLUT4: inducibile da insulina

22
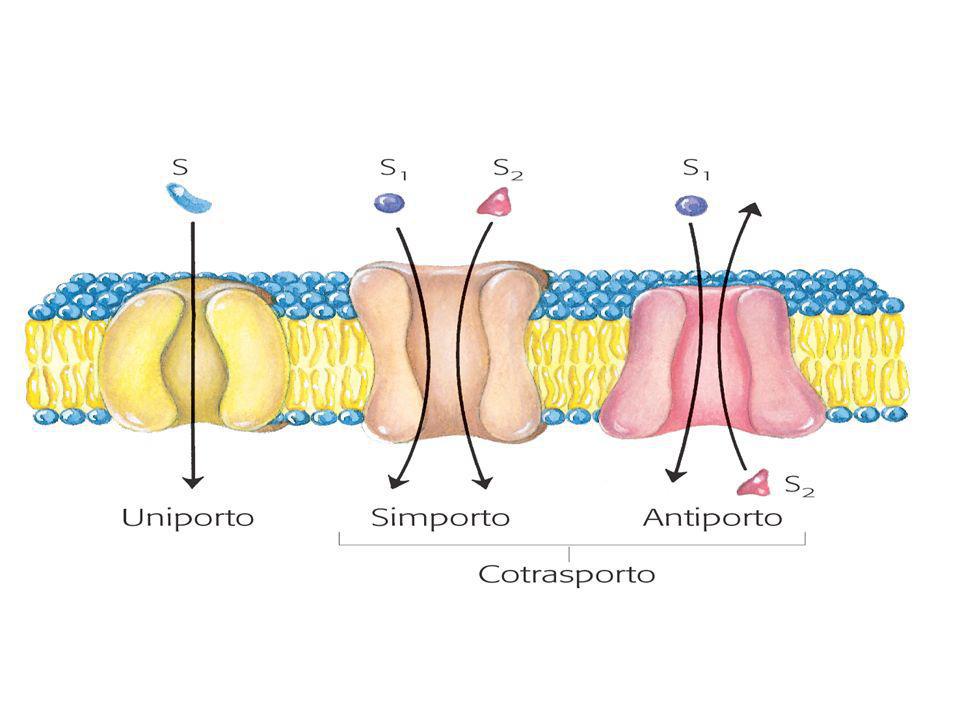
Sistemi di trasporto passivo
Antiport per lo scambio ADP-ATP Simport per l’ingressp H+ e fosfato Trasportatore acidi bicarbossilici ( citrato) Antiport aspartato –glutammato Tutti questi trasportatori sono legati alla produzione di energia e la sua conservazione. Antiport clururo/bicarbonato del rene e dei globuli rossi.
 Antiport aspartato –glutammato. Tutti questi trasportatori sono legati alla produzione di energia e la sua conservazione. Antiport clururo/bicarbonato del rene e dei globuli rossi.")
23
Processi di trasporto attivo
Richiedono molta energia, ma… Consentono di spostare sostanze contro un gradiente (di concentrazione, elettrico, ecc.)
")
24
Trasportatori attivi primari
Si classificano in trasportati di tipo : P (fosfato): durante il trasporto sono fosforilati e defosforilati.Se ne conocono circa 300. V ( come vacuolo): pompe protoniche presenti nei lisosomi,endosomi,app. Golgi e vescicole secretorie:acidificano l’interno della struttura. F (presenti nei mitocondri e nei cloroplasti ) può idrolizzare o sintetizzare ATP in funzione del flusso di H+.
: durante il trasporto sono fosforilati e defosforilati.Se ne conocono circa 300. V ( come vacuolo): pompe protoniche presenti nei lisosomi,endosomi,app. Golgi e vescicole secretorie:acidificano l’interno della struttura. F (presenti nei mitocondri e nei cloroplasti ) può idrolizzare o sintetizzare ATP in funzione del flusso di H+.")
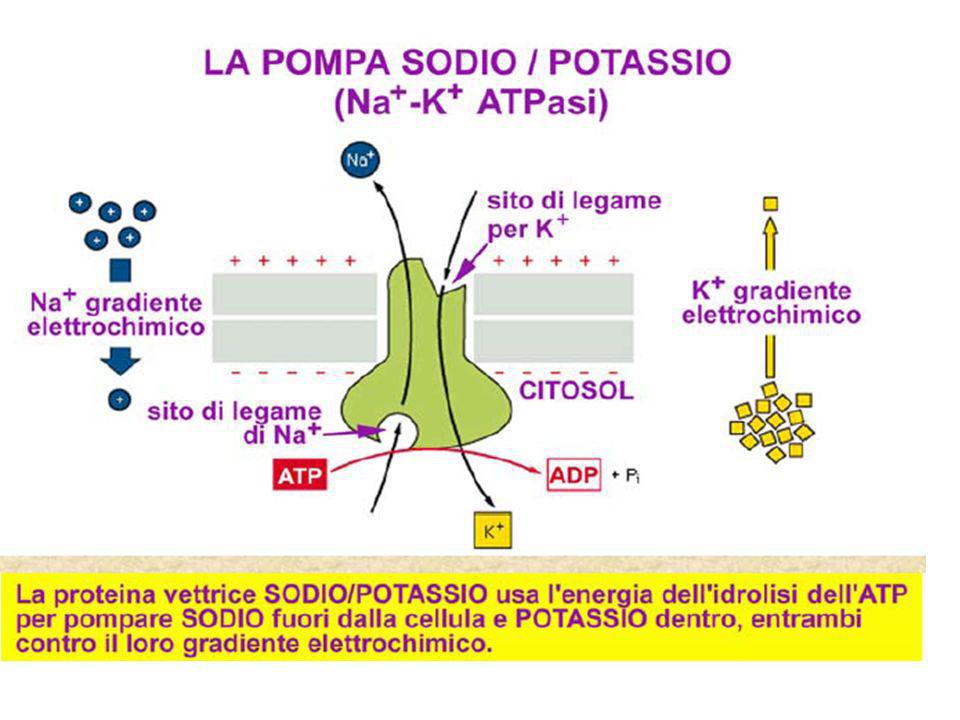
26
La pompa, con ATP legata, lega 3 ioni Na+ intracellulari
3 Na+ fuori La pompa cambia conformazione esponendo gli ioni Na+ fuori dalla cellula dove vengono rilasciati P ATP viene idrolizzata, portando alla fosforilazione della subunità a della pompa P 3 Na+ intracell. ATP b a La pompa, con ATP legata, lega 3 ioni Na+ intracellulari 2 K+ extrac. La pompa lega 2 ioni K+ extracellulari, causando la defosforilazione della subunità a P 2 K+ dentro ATP ATP si lega alla pompa che si riorienta rilasciando i 2 ioni K+ all’interno della cellula Na+/K+ATPasi P

27
Trasporto attivo secondario

28
Necessità di Energia Caratteristiche Strutturali
Non richiede ulteriore energia oltre a quella derivata dal movimento molecolare Necessità di Energia Richiede ATP ESOCITOSI FAGOCITOSI ENDOCITOSI Trasporto Attivo PRIMARIO SECONDARIO Crea gradienti per DIFFUSIONE Diffusione Semplice Facilitata Vescicole rivestite da membrana Molecole attraversano il doppio strato lipidico Caratteristiche Strutturali Trasporto mediato da proteine di membrana

Presentazioni simili













>")








