Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Corso di Laurea in Scienze del Fitness e dei Prodotti della Salute
BIOCHIMICA CLINICA Corso di Laurea in Scienze del Fitness e dei Prodotti della Salute Lezione 2 Il ciclo nutrizione / digiuno Testi consigliati Le Basi della Biochimica- P. C.Champe, R.A.Harvey, D.R. Ferrier- Ed Zanichelli Biochimica per le Discipline Biomediche- J.W. Baynes, M.H.Dominiczak- Casa Editrice Ambrosiana Le Immagini sono state tratte dai testi sopra indicati.

2
Aspetti generali La fase di assorbimento è il periodo che va dalle 2 alle 4 ore successive all’assunzione di un pasto. Durante questo intervallo si verifica un transitorio aumento del livello ematico del glucosio, degli amminoacidi e dei triacilgliceroli, che si trovano principalmente nei chilomicroni sintetizzati dalle cellule della mucosa intestinale. Il tessuto insulare pancreatico risponde a un livello elevato di glucosio e di amminoacidi con un aumento della secrezione di insulina e una diminuzione di quella del glucagone. L’elevato rapporto insulina/glucagone e l’immediata disponibilità di substrati in circolo che si ha nelle prime ore dopo un pasto porta a un aumento della sintesi dei triacilgliceroli, del glicogeno e delle proteine. Durante questa fase, praticamente tutti i tessuti utilizzano il glucosio come fonte di energia e la risposta metabolica dell’organismo è dominata da variazioni del metabolismo del fegato, del tessuto adiposo, dei muscoli e dell’encefalo. Introdurremo una mappa degli organi che individua gli spostamenti dei metaboliti tra i tessuti, permettendo di avere una visione complessiva e utile sul piano clinico del metabolismo corporeo.

3
Le variazioni delle attività enzimatiche nelle condizioni di apporto di nutrienti
Il flusso di intermedi lungo le vie metaboliche è controllato da quattro meccanismi che operano ciascuno secondo una scala temporale diversa, permettendo all’organismo di adeguarsi ad un’ampia varietà di situazioni fisiologiche: la disponibilità di substrati l’attivazione o l’inibizione allosterica di enzimi le modificazioni covalenti di enzimi l’induzione o la repressione della sintesi di enzimi In condizioni di apporto alimentare mediante tali meccanismi regolatori dai nutrienti disponibili si formano glicogeno, triacilgliceroli e proteine.

4
Le variazioni delle attività enzimatiche nelle
condizioni di apporto di nutrienti Gli effetti allosterici La regolazione degli enzimi mediante la modifica covalente L’induzione e repressione della sintesi degli enzimi legenda intermedi del metabolismo dei carboidrati intermedi del metabolismo dei lipidi

5
Il fegato: un centro di distribuzione dei nutrienti
Il fegato occupa una posizione strategica per l’elaborazione e la distribuzione dei nutrienti di provenienza alimentare, dal momento che il drenaggio venoso dell’intestino e del pancreas raccoglie le sostanze nutrienti nella vena porta epatica prima di entrare nella circolazione generale. Il sangue che il fegato riceve dopo un pasto è quindi ricco di nutrienti di origine alimentare e contiene un livello elevato di insulina, secreta dal pancreas. Durante il periodo dell’assorbimento intestinale, il fegato assume carboidrati, lipidi e la maggior parte degli amminoacidi. Questi nutrienti sono poi metabolizzati e conservati oppure inviati verso altri tessuti. Il fegato ha perciò il ruolo di attenuare le fluttuazioni, potenzialmente ampie, dei nutrienti a disposizione dei tessuti periferici.

6
Il metabolismo dei carboidrati
Normalmente il fegato produce glucosio, piuttosto che consumarlo. Tuttavia, dopo un pasto contenente carboidrati, il fegato trattiene circa 60 grammi di glucosio ogni 100 grammi di zucchero presenti nel sistema portale e li utilizza per tutte le sue attività. Il metabolismo epatico del glucosio aumenta attraverso i seguenti meccanismi: aumento della fosfolirazione del glucosio (1), aumento della sintesi del glicogeno (2), aumento dell’attività della via dell’esoso monofosfato (HMP) (3), aumento della glicolisi (4), diminuzione della gluconeogenesi
, aumento della sintesi del glicogeno (2), aumento dell’attività della via dell’esoso monofosfato (HMP) (3), aumento della glicolisi (4), diminuzione della gluconeogenesi.")
7
Il metabolismo dei grassi
aumento della sintesi degli acidi grassi ( 5) aumento della sintesi dei triacilgliceroli (6)
 aumento della sintesi dei triacilgliceroli (6)")
8
Il metabolismo degli amminoacidi
aumento della degradazione degli amminoacidi (7) aumento della sintesi proteica (8)
 aumento della sintesi proteica (8)")
9
Il tessuto adiposo: un deposito di scorte energetiche
La capacità del tessuto adiposo di distribuire molecole di combustibile è inferiore soltanto a quella del fegato. In un uomo di 70 kg il tessuto adiposo pesa, in media, 14 kg, circa la metà della massa muscolare complessiva. Negli individui obesi il tessuto adiposo può costituire fino al 70% della massa corporea. Lo spazio interno di un adipocita può essere occupato quasi completamente da una goccia di triacilglicerolo.

10
Il tessuto adiposo: un deposito di scorte energetiche
Il metabolismo dei carboidrati: aumento del trasporto del glucosio (1) aumento della glicolisi (2) aumento dell’attività dell’esoso monofosfato (HMP) (3)
 aumento della glicolisi (2) aumento dell’attività dell’esoso monofosfato (HMP) (3)")
11
Il tessuto adiposo: un deposito di scorte energetiche
Il metabolismo dei grassi: aumento della sintesi degli acidi grassi (4) aumento della sintesi triacilgliceroli (5) (6) diminuzione della degradazione dei triacilgliceroli
 aumento della sintesi triacilgliceroli (5) (6) diminuzione della degradazione dei triacilgliceroli.")
12
Il muscolo scheletrico a riposo
Il metabolismo energetico del muscolo scheletrico è esclusivo per la sua capacità di rispondere ai sostanziali cambiamenti nella richiesta di ATP che accompagnano la contrazione muscolare. A riposo il consumo di ossigeno è approssimativamente il 30% del consumo corporeo totale, ma può raggiungere il 90% durante un esercizio fisico intenso. Quindi il muscolo scheletrico, nonostante abbia la capacità potenziale di utilizzare, per un periodo transitorio, la glicolisi anaerobica, è un tessuto ossidativo. Il muscolo cardiaco differisce dal muscolo scheletrico per tre aspetti importanti: il cuore è in continua attività, mentre i muscoli scheletrici si contraggono in modo intermittente, in seguito ad una sollecitazione. il cuore ha un metabolismo completamente aerobico. il cuore contiene riserve energetiche ( glicogeno o lipidi) in quantità trascurabile. Ne consegue che l’interruzione del rifornimento ematico, quale quello che si verifica durante un infarto del miocardio, provoca rapidamente la morte delle cellule miocardiche. Le sostanze dalle quali il muscolo cardiaco ricava energia sono il glucosio libero, gli acidi grassi e i corpi chetonici.
 in quantità trascurabile. Ne consegue che l’interruzione del rifornimento ematico, quale quello che si verifica durante un infarto del miocardio, provoca rapidamente la morte delle cellule miocardiche. Le sostanze dalle quali il muscolo cardiaco ricava energia sono il glucosio libero, gli acidi grassi e i corpi chetonici.")
13
Il muscolo scheletrico a riposo
Il metabolismo dei carboidrati: aumento del trasporto del glucosio (1) aumento della sintesi del glicogeno (2) Il metabolismo degli acidi grassi: Gli acidi grassi si liberano dai chilomicroni e dalle VLDL per azione della lipoproteina lipasi. Tuttavia, nel periodo di apporto di nutrienti, gli acidi grassi sono combustibili di importanza secondaria per il muscolo, essendo invece il glucosio la fonte primaria. Il metabolismo degli amminoacidi: aumento della sintesi proteica (3) e (4) aumento dell’assunzione degli amminoacidi a catena ramificata leucina, isoleucina e valina (3).
 aumento della sintesi del glicogeno (2) Il metabolismo degli acidi grassi: Gli acidi grassi si liberano dai chilomicroni e dalle VLDL per azione della lipoproteina lipasi. Tuttavia, nel periodo di apporto di nutrienti, gli acidi grassi sono combustibili di importanza secondaria per il muscolo, essendo invece il glucosio la fonte primaria. Il metabolismo degli amminoacidi: aumento della sintesi proteica (3) e (4) aumento dell’assunzione degli amminoacidi a catena ramificata leucina, isoleucina e valina (3).")
14
L’encefalo Benché costituisca solo il 2% della massa corporea di un adulto, l’encefalo consuma il 20% a riposo dell’ossigeno utilizzato dall’organismo. L’encefalo consuma energia ad un ritmo costante.Poiché la sua attività è vitale per un corretto funzionamento dell’organismo, soddisfare il suo fabbisogno energetico diventa quindi prioritario. Per poter essere utilizzati, i substrati devono superare le cellule endoteliali che rivestono il lume dei vasi sanguigni encefalici ( barriera emato-encefalica). Normalmente il glucosio funge da combustibile primario, dal momento che, nel periodo di apporto di nutrienti, la concentrazione dei corpi chetonici è troppo bassa perché essi siano una fonte alternativa di energia. Se il glucosio ematico crolla ad un livello inferiore a circa 30 mg/100 mL ( livello normale mg/100 mL), la funzionalità cerebrale è compromessa. Può verificarsi un danno cerebrale grave e potenzialmente irreversibile anche qualora l’ipoglicemia perduri per un breve periodo. I corpi chetonici, in quanto combustibili, svolgono un ruolo significativo nei periodi di digiuno.
. Normalmente il glucosio funge da combustibile primario, dal momento che, nel periodo di apporto di nutrienti, la concentrazione dei corpi chetonici è troppo bassa perché essi siano una fonte alternativa di energia. Se il glucosio ematico crolla ad un livello inferiore a circa 30 mg/100 mL ( livello normale mg/100 mL), la funzionalità cerebrale è compromessa. Può verificarsi un danno cerebrale grave e potenzialmente irreversibile anche qualora l’ipoglicemia perduri per un breve periodo. I corpi chetonici, in quanto combustibili, svolgono un ruolo significativo nei periodi di digiuno.")
15
L’encefalo Il metabolismo dei carboidrati (1)
Il metabolismo dei grassi

16
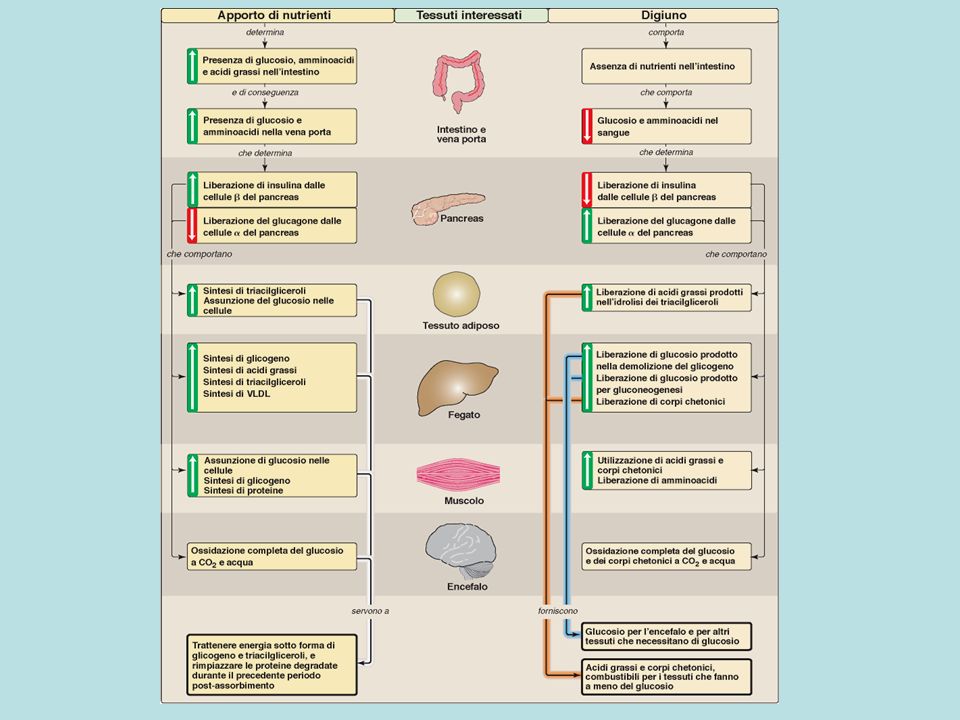
Relazioni tra tessuti durante il periodo di assorbimento

17
Il digiuno: uno sguardo d’insieme
Il digiuno può dipendere dall’impossibilità di reperire del cibo, dal desiderio di perdere peso rapidamente o da una situazione clinica che comporti l’impossibilità ad alimentarsi in seguito ad un trauma, ad un intervento chirurgico, a una neoplasia, a ustioni etc… In mancanza di cibo, i livelli plasmatici di glucosio, degli amminoacidi e dei triacilgliceroli tendono a diminuire; contemporaneamente la secrezione dell’insulina decresce mentre aumenta la liberazione del glucagone. La diminuzione del rapporto tra il livello dell’insulina e il livello del glucagone e la ridotta disponibilità di substrati circolanti rendono il periodo di privazioni di nutrienti un periodo catabolico, caratterizzato dalla degradazione di triacilgliceroli, glicogeno e proteine. Ciò mette in moto uno scambio di substrati tra il fegato, il tessuto adiposo, il tessuto muscolare e l’encefalo, sotto la spinta di due priorità: l’esigenza di mantenere un livello plasmatico del glucosio tale da sostenere il metabolismo energetico dell’encefalo e di altri tessuti che utilizzano questo zucchero la necessità di mobilizzare acidi grassi dal tessuto adiposo e di attivare la sintesi e la liberazione dei corpi chetonici da parte del fegato, per rifornire di energia gli altri tessuti.

18
Le scorte energetiche I combustibili metabolici presenti in un uomo di 70 kg all’esordio di un digiuno

19
I cambiamenti enzimatici nel digiuno
Anche nel digiuno il flusso di intermedi lungo le vie del metabolismo intermedio è controllato da quattro meccanismi: la disponibilità dei substrati l’attivazione o l’inibizione allosterica di enzimi la modificazione covalente di enzimi l’induzione o la repressione della sintesi di enzimi Enzimi: stato fosforilato (inattivi) Eccezioni: glicogeno fosforilasi, fruttosio bisfosfato fosfatasi-2 e lipasi sensibile agli ormoni del tessuto adiposo, sono inattive nello stato defosforilato Molte delle modificazioni osservate nel digiuno sono l’opposto di quelle che si verificano in condizione di apporto di nutrienti.
 Eccezioni: glicogeno fosforilasi, fruttosio bisfosfato fosfatasi-2 e lipasi sensibile agli ormoni del tessuto adiposo, sono inattive nello stato defosforilato. Molte delle modificazioni osservate nel digiuno sono l’opposto di quelle che si verificano in condizione di apporto di nutrienti.")
20
Il fegato durante il digiuno
Il ruolo primario del fegato nel metabolismo energetico durante il digiuno è la sintesi e la distribuzione di molecole di combustibile ad altri organi si parla quindi di “metabolismo epatico” e di “metabolismo extraepatico o periferico”. Il metabolismo dei carboidrati. Le fonti di glucosio ematico dopo l’assunzione di 100g di zucchero.

21
Il metabolismo dei carboidrati
Aumento della demolizione del glicogeno (1) Aumento della gluconeogenesi (2) Il metabolismo dei grassi Aumento dell’ossidazione degli acidi grassi (3) Aumento della sintesi dei corpi chetonici (4) (principalmente 3-idrossibutirrato)
 Aumento della gluconeogenesi (2) Il metabolismo dei grassi. Aumento dell’ossidazione degli acidi grassi (3) Aumento della sintesi dei corpi chetonici (4) (principalmente 3-idrossibutirrato)")
22
Aumento della sintesi dei corpi chetonici
Il fegato ha la capacità esclusiva di sintetizzare e liberare i corpi chetonici, principalmente il 3-idrossibutirrato, che i tessuti periferici possono usare come combustibile. Il fegato non può utilizzare i corpi chetonici come combustibile. La sintesi dei corpi chetonici è favorita quando la concentrazione dell’acetil CoA prodotto dal metabolismo degli acidi grassi eccede la capacità ossidativa del ciclo dei TCA. Una sintesi significativa di corpi chetonici ha inizio durante i primi giorni di digiuno. Contrariamente agli acidi grassi, i corpi chetonici sono idrosolubili e compaiono nel sangue e nell’urina a partire dal secondo giorno di digiuno. Nel digiuno la disponibilità di corpi chetonici in circolo è importante, perché la maggioranza dei tessuti può utilizzarli come combustibile, compreso l’encefalo, una volta che il loro livello ematico abbia raggiunto un valore sufficientemente elevato. Ciò riduce il bisogno di sintetizzare glucosio attraverso la gluconeogenesi a partire dagli scheletri carboniosi di amminoacidi, rallentando quindi la perdita di proteine essenziali. La concentrazione ematica degli acidi grassi e del 3-idrossibutirrato durante il digiuno

23
Il tessuto adiposo durante il digiuno
Il metabolismo dei carboidrati: il trasporto del glucosio dentro gli adipociti e il suo successivo metabolismo sono depressi a causa del basso livello di insulina in circolo. Ne consegue una diminuita sintesi di acidi grassi e triacilgliceroli. Il metabolismo dei grassi Aumento della degradazione dei triacilgliceroli: l’aumento dell’adrenalina e della noradrenalina, liberate dalle terminazioni simpatiche, stimola l’attività della lipasi sensibile agli ormoni con conseguente idrolisi dei triacilgliceroli depositati nell’adipocita (1) Aumento della liberazione degli acidi grassi: gli acidi grassi liberi prodotti dall’idrolisi dei triacilgliceroli di deposito sono rilasciati nel sangue e legati all’albumina raggiungono una molteplicità di tessuti e sono utilizzati per produrre energia. Il glicerolo è utilizzato dal fegato come precursore gluconeogenetico. Gli acidi grassi sono convertiti in acetil-CoA, che entra poi nel ciclo dei TCA liberando energia. Diminuzione dell’assunzione degli acidi grassi: nel digiuno l’attività della lipoproteina lipasi è bassa per cui i triacilgliceroli circolanti sotto forma di lipoproteine non sono disponibili per la sintesi dei triacilgliceroli nel tessuto adiposo.
 Aumento della liberazione degli acidi grassi: gli acidi grassi liberi prodotti dall’idrolisi dei triacilgliceroli di deposito sono rilasciati nel sangue e legati all’albumina raggiungono una molteplicità di tessuti e sono utilizzati per produrre energia. Il glicerolo è utilizzato dal fegato come precursore gluconeogenetico. Gli acidi grassi sono convertiti in acetil-CoA, che entra poi nel ciclo dei TCA liberando energia. Diminuzione dell’assunzione degli acidi grassi: nel digiuno l’attività della lipoproteina lipasi è bassa per cui i triacilgliceroli circolanti sotto forma di lipoproteine non sono disponibili per la sintesi dei triacilgliceroli nel tessuto adiposo.")
24
Il muscolo scheletrico a riposo durante il digiuno
Nel digiuno, il muscolo a riposo utilizza gli acidi grassi come fonte principale di energia. Al contrario il muscolo in esercizio inizialmente utilizza come fonte di combustibile le proprie scorte di glicogeno durante uno sforzo intenso, il glucosio 6-fosfato derivante dal glicogeno è convertito in lattato con la glicolisi anaerobica. Quando le scorte di glicogeno si esauriscono, gli acidi grassi liberi derivanti dalla mobilizzazione dei triacilgliceroli del tessuto adiposo diventano la fonte dominante di energia. Metabolismo dei carboidrati: il trasporto del glucosio all’interno delle fibre del muscolo scheletrico e il conseguente metabolismo sono depressi a causa del basso livello di insulina in circolo. Metabolismo dei lipidi: durante le prime due settimane di digiuno, il muscolo utilizza come combustibili gli acidi grassi derivanti dal tessuto adiposo e i corpi chetonici prodotti dal fegato (1 e 2). Dopo circa tre settimane di digiuno, l’utilizzo dei corpi chetonici diminuisce e il muscolo usa quasi esclusivamente gli acidi grassi, con ulteriore aumento dei corpi chetonici in circolo e loro maggior utilizzo da parte dell’encefalo. Metabolismo delle proteine: durante i primi giorni di digiuno si verifica una rapida demolizione delle proteine muscolari che forniscono al fegato amminoacidi (principalmente alanina e glutammina) per la gluconeogenesi (3). Dopo varie settimane di digiuno la proteolisi muscolare diminuisce perché l’encefalo ha un minore bisogno di glucosio perché utilizza, come fonte di energia, i corpi chetonici.
. Dopo circa tre settimane di digiuno, l’utilizzo dei corpi chetonici diminuisce e il muscolo usa quasi esclusivamente gli acidi grassi, con ulteriore aumento dei corpi chetonici in circolo e loro maggior utilizzo da parte dell’encefalo. Metabolismo delle proteine: durante i primi giorni di digiuno si verifica una rapida demolizione delle proteine muscolari che forniscono al fegato amminoacidi (principalmente alanina e glutammina) per la gluconeogenesi (3). Dopo varie settimane di digiuno la proteolisi muscolare diminuisce perché l’encefalo ha un minore bisogno di glucosio perché utilizza, come fonte di energia, i corpi chetonici.")
25
L’encefalo nel digiuno
Durante i primi giorni di digiuno, l’encefalo continua ad utilizzare come fonte di energia esclusivamente il glucosio (1). La gluconeogenesi epatica riesce a mantenere il livello ematico del glucosio utilizzando precursori glucogenici quali gli amminoacidi forniti dalla rapida demolizione della proteine muscolari. Nel digiuno protratto (2-3 settimane) i corpi chetonici raggiungono livelli significativamente elevati e l’encefalo li utilizza come combustibili in aggiunta al glucosio (2). Ciò riduce la richiesta del catabolismo proteico per alimentare la gluconeogenesi. I cambiamenti metabolici che si verificano durante il digiuno garantiscono a tutti i tessuti un apporto adeguato di molecole combustibili.
. La gluconeogenesi epatica riesce a mantenere il livello ematico del glucosio utilizzando precursori glucogenici quali gli amminoacidi forniti dalla rapida demolizione della proteine muscolari. Nel digiuno protratto (2-3 settimane) i corpi chetonici raggiungono livelli significativamente elevati e l’encefalo li utilizza come combustibili in aggiunta al glucosio (2). Ciò riduce la richiesta del catabolismo proteico per alimentare la gluconeogenesi. I cambiamenti metabolici che si verificano durante il digiuno garantiscono a tutti i tessuti un apporto adeguato di molecole combustibili.")
26
Relazioni metaboliche tra i tessuti durante il digiuno

Presentazioni simili