Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore


1
Si può dimostrare che la capacità portante vale
Qualunque sia il valore iniziale della popolazione, a lungo termine essa si assesterà alla capacità portante

2
I PUNTI DI EQUILIBRIO DEL MODELLO DI BEVERTON-HOLT

3
Punti di equilibrio del modello di Beverton-Holt
instabile È equilibrio stabile

4
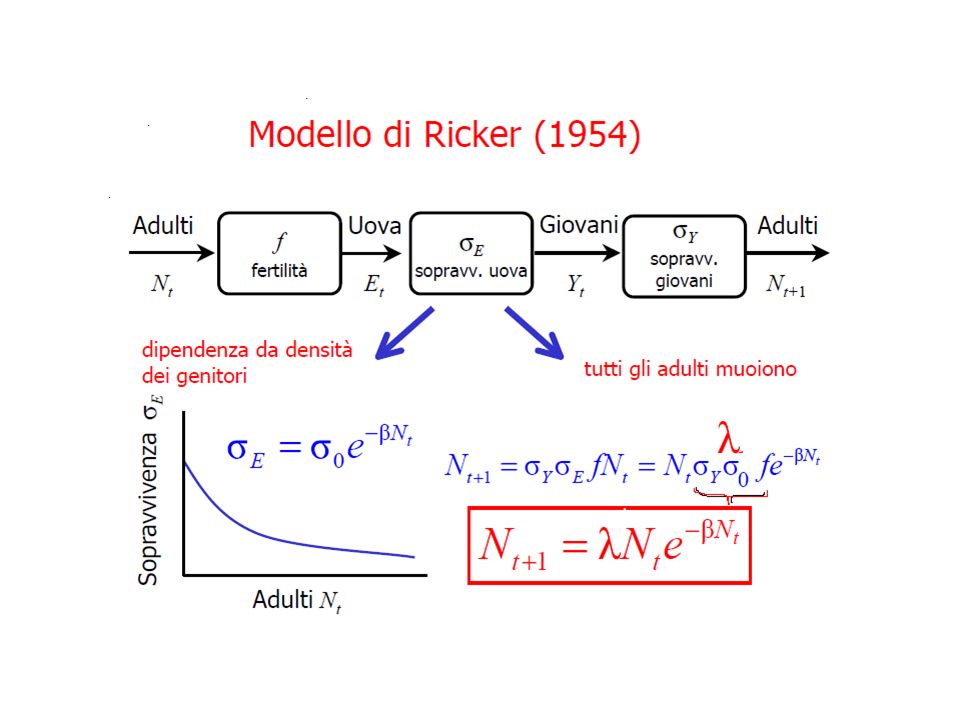
IL MODELLO DI RICKER (1954)
")
5
Pacifico Il salmone del Oncorhynchus Durante il ciclo riproduttivo, le
femmine di salmone ricercano un luogo nel greto del fiume dove deporre le uova, in attesa che vengano fecondate da un maschio. Al crescere della densità degli adulti, cresce la probabilità che più femmine scelgano lo stesso luogo. Oncorhynchus distruzione accidentale di alcune delle uova con conseguente decomposizione proliferazione di diversi agenti patogeni che infestano le uova deposte Inoltre in alcuni casi, si sono anche osservati fenomeni di cannibalismo degli adulti nei riguardi delle proprie uova. è ragionevole pensare che la sopravvivenza dei nuovi nati decresca con il numero di adulti riproduttivi non in modo lineare

7
COMPORTAMENTO DEL MODELLO DI RICKER
( DINAMICHE CAOTICHE )
")
8
Dopo un breve periodo, la popolazione si assesta al valore di equilibrio
non banale

9
Se si ha un andamento ciclico (di periodo 2).
Si alternano anni a popolazione scarsa con anni a popolazione abbondante L’equilibrio non banale è instabile: partendo da densità vicine ad esso, si tende ad allontanarsi. Ci si avvicina invece ciclicamente ad altri 2 punti (non di equilibrio)
")
10
Dopo un breve transitorio, la popolazione oscilla con periodicità tra 4 valori diversi. Ciclo di periodo 4 Ad ogni anno di elevata abbondanza segue regolarmente un anno di bassa densità, ma di valore non costante. I minimi e i massimi assumono alternativamente valori diversi.

11
Comportamento caotico
L’abbondanza della popolazione ora fluttua in maniera caotica. (caos deterministico)
")
12
Valori iniziali diversi :
Il caos deterministico determina una sensibile dipendenza della soluzione dai dati iniziali. I valori previsti dal modello si differenziano sempre di più e addirittura sono in contro-fase dopo il trentesimo passo (max e min scambiati)
")
13
Le aringhe del Mare del Nord
APPLICAZIONE DEL MODELLO DI RICKER Le aringhe del Mare del Nord

14
La relazione stock-reclutamento per questo tipo di popolazione ha prodotto la tabella:
0.69 0.00 10.03 25.39 16.36 37.16 22.41 46.’02 29.00 53.56 36.43 59.12 44.81 61.76 52.10 61.63 59.79 59.25 81.79 48.50 105.57 33.20 119.86 24.20 136.50 16.00 163.99 7.67

15
Tramite una procedura di fitting ai minimi quadrati è possibile risalire alla funzione relativa al modello di Ricker capace di descrivere la dinamica di questa popolazione clear all global Nt Nt0 Nt1 % Numero di aringhe (10^3 tonellate)al tempo t Nt=[ ]; % Numero di aringhe (10^3 tonellate)al tempo t+1 Nt1=[ ]; plot(Nt,Nt1,'o') title('Stock-Reclutamento per una popolazione di aringhe del Mare del nord') xlabel('Nt [ 10^3 tonnellate]') ylabel('Nt+1 [ 10^3 tonnellate] ')
al tempo t. Nt=[ ]; % Numero di aringhe (10^3 tonellate)al tempo t+1. Nt1=[ ]; plot(Nt,Nt1, o ) title( Stock-Reclutamento per una popolazione di aringhe del Mare del nord ) xlabel( Nt [ 10^3 tonnellate] ) ylabel( Nt+1 [ 10^3 tonnellate] )")
16
% Approssimazione ai minimi quadrati con il modello di Richer
% N(t+1)= lambda* N(t)*exp(-K*N(t)) % p = [ lambda, K ]: parametri da identificare % ======================================== Nt0= Nt(1); p=[ ]; % valori iniziali dei parametri da identificare options=optimset('TolX',0.001); [p,fmin,exit,out]=fminsearch('Rerr',p,options); % Rappresentazione grafica dei risultati % y dati calcolati con il modello % Nt1 dati misurati for i=1:length(Nt) y(i)= ricker(p,Nt(i)); end
= lambda* N(t)*exp(-K*N(t)) % p = [ lambda, K ]: parametri da identificare. % ======================================== Nt0= Nt(1); p=[ ]; % valori iniziali dei parametri da identificare. options=optimset( TolX ,0.001); [p,fmin,exit,out]=fminsearch( Rerr ,p,options); % Rappresentazione grafica dei risultati. % y dati calcolati con il modello. % Nt1 dati misurati. for i=1:length(Nt) y(i)= ricker(p,Nt(i)); end.")
17
%%%%%%%%%%%%%%%%%%%%%%%%%%%% % Calcolo dell'errore
% tra i valori misurati e quelli calcolati (modello di Ricker) % % ( Errore = Distanza euclidea) %%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%% function z=Rerr(p) global Nt Nt0 Nt1 len = length(Nt); for i=1:len y(i)= ricker(p,Nt(i)); end z=norm(y-Nt1); %%%%%%%%%%%%%%%%%%%% % Definizione del MODELLO DI % RICKER % % N(t+1)= lambda* N(t)*exp(-K*N(t)) function y =ricker(param,N) global Nt Nt0 Nt1 lambda=param(1); K=param(2); y=lambda*N*exp(-K*N);
 % % ( Errore = Distanza euclidea) %%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%% function z=Rerr(p) global Nt Nt0 Nt1. len = length(Nt); for i=1:len. y(i)= ricker(p,Nt(i)); end. z=norm(y-Nt1); %%%%%%%%%%%%%%%%%%%% % Definizione del MODELLO DI. % RICKER. % % N(t+1)= lambda* N(t)*exp(-K*N(t)) function y =ricker(param,N) global Nt Nt0 Nt1. lambda=param(1); K=param(2); y=lambda*N*exp(-K*N);")
18
figure(2) plot(Nt,Nt1,'o',Nt,y) title( 'Modello di Ricker per una popolazione di aringhe') xlabel('Nt [ 10^3 tonnellate]') ylabel('Nt+1 [ 10^3 tonnellate]') legend('Dati misurati','Modello di Ricker') 3.7331 0.0234
![figure(2) plot(Nt,Nt1, o ,Nt,y) title( Modello di Ricker per una popolazione di aringhe ) xlabel( Nt [ 10^3 tonnellate] )](http://slideplayer.it/slide/534693/1/images/18/figure%282%29+plot%28Nt%2CNt1%2C+o+%2CNt%2Cy%29+title%28+Modello+di+Ricker+per+una+popolazione+di+aringhe+%29+xlabel%28+Nt+%5B+10%5E3+tonnellate%5D+%29.jpg "ylabel( Nt+1 [ 10^3 tonnellate] ) legend( Dati misurati , Modello di Ricker )")
19
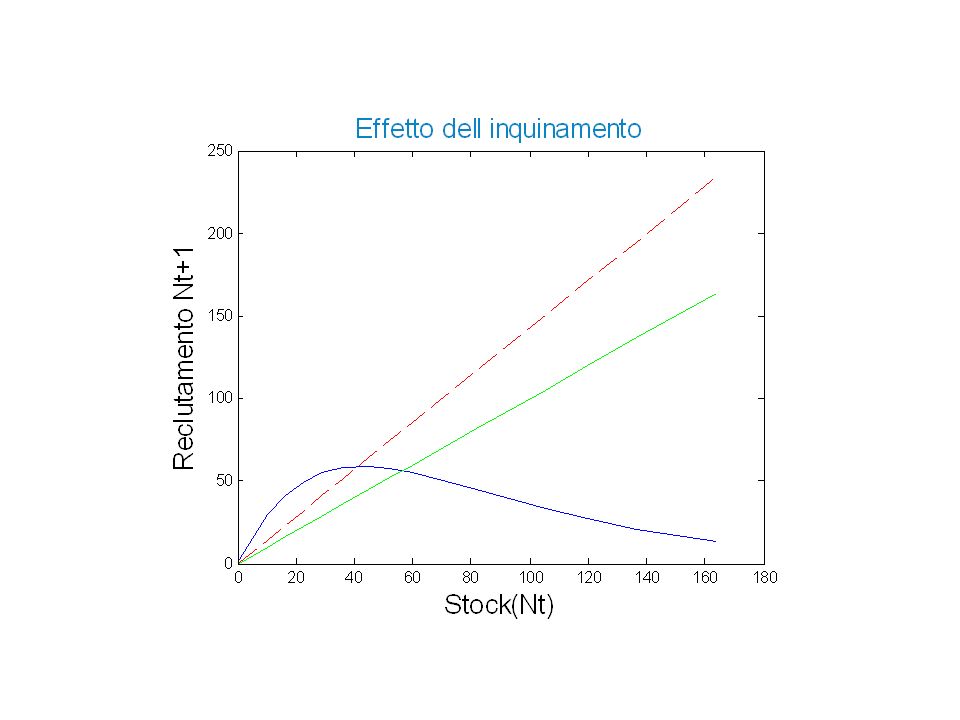
In una certa annata si è verificato un peggioramento della qualità delle acque dovuto all’inquinamento La mortalità delle aringhe è aumentata del 30% ogni anno Variante del Modello di Ricker che prende in considerazione l’effetto dell’inquinamento Ricker Inquinamento

20
Punti di equilibrio in assenza e con inquinamento
Senza inquinamento: Intersezione della curva con la bisettrice Con inquinamento Intersezione della curva con la retta di coefficiente angolare 1\0.7 (bisettrice ruotata)
")
22
%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%
% Equilibrio in caso di inquinamento (morte del 30% delle aringhe) % % f(Nt+1) = f(Nt) -0.3*f(Nt) % % ==> equilibrio: f(Nt)[1-0.3] = Nt % ==> intersezione di f(Nt) con la retta: Nt / [1-0.3] %%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%% figure(3) equi=Nt; inqui=1/(1-0.3); y2=inqui*Nt; plot(Nt,y,Nt,equi,'g',Nt,y2,'--r') title('Effetto dell inquinamento') xlabel('Stock(Nt)') ylabel('Reclutamento Nt+1')
 % % f(Nt+1) = f(Nt) -0.3*f(Nt) % % ==> equilibrio: f(Nt)[1-0.3] = Nt. % ==> intersezione di f(Nt) con la retta: Nt / [1-0.3] %%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%%% figure(3) equi=Nt; inqui=1/(1-0.3); y2=inqui*Nt; plot(Nt,y,Nt,equi, g ,Nt,y2, --r ) title( Effetto dell inquinamento ) xlabel( Stock(Nt) ) ylabel( Reclutamento Nt+1 )")
23
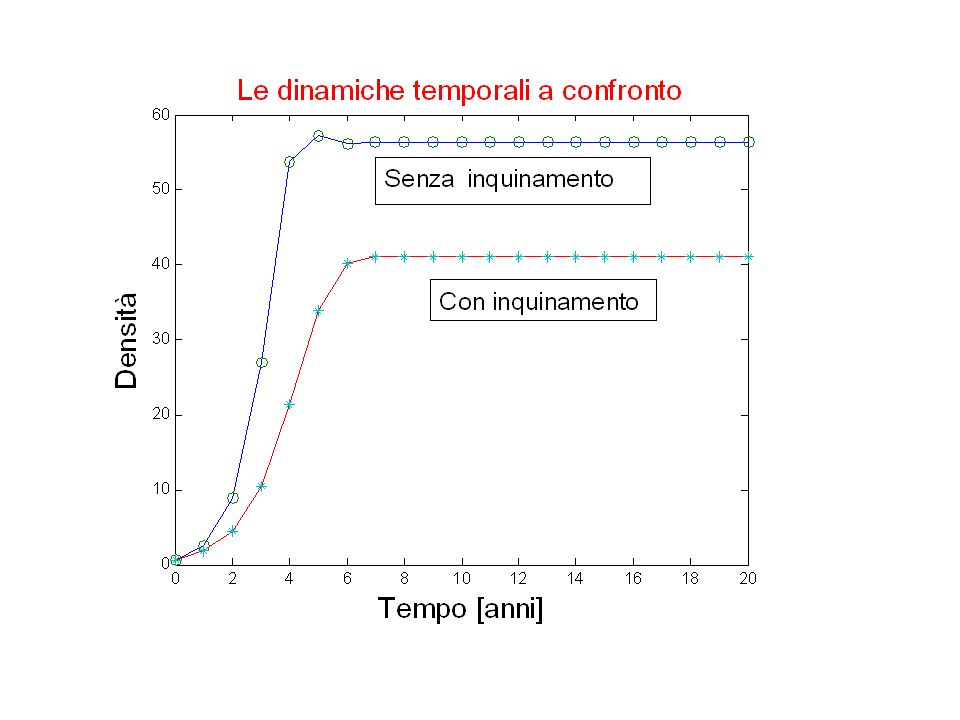
Dinamiche temporali a confronto
a1(1)=Nt0; a2(1)=a1(1); for i=1:length(T)-1 a1(i+1)=ricker(p,a1(i)); a2(i+1)=ricker(p,a2(i)); a2(i+1)=a2(i+1)-0.3*a2(i+1); end plot(T,a1,T,a1,'o',T,a2,T,a2,'*') title('Le dinamiche temporali a confronto') xlabel('Tempo [anni]') ylabel('Densità')
=Nt0; a2(1)=a1(1); for i=1:length(T)-1. a1(i+1)=ricker(p,a1(i)); a2(i+1)=ricker(p,a2(i)); a2(i+1)=a2(i+1)-0.3*a2(i+1); end. plot(T,a1,T,a1, o ,T,a2,T,a2, * ) title( Le dinamiche temporali a confronto ) xlabel( Tempo [anni] ) ylabel( Densità )")
25
EFFETTO ALLEE Warder Clyde Allee Biologo ecologista USA 1885-1955
Il tasso di accrescimento dei modelli di Beverton-Holl e di Ricker decresce all’aumentare della popolazione In alcune popolazioni e in certe condizioni ambientali il tasso di accrescimento è massimo ad una densità di individui intermedia tra la minima e la massima. Per valori della popolazione non così grandi da rendere rilevanti l’effetto della competizione intraspecifica, l’aumento della popolazione provoca un aumento del tasso di accrescimento

26
Ciò è vero per popolazioni dotate di una struttura sociale (lupi), per le quali l’aumento della numerosità significa maggiore aiuto, sicurezza e protezione del singolo individuo. Si vuole costruire una funzione che per valori di P piccoli risulti crescente, mentre per valori grandi di P risulti decrescente Questo tipo di dipendenza da densità prende il nome di DEPENSAZIONE. Una parabola rivolta verso il basso soddisfa le ipotesi che abbiamo posto su r(P).
.")
27
Due situazioni possibili :
Pd < 0 Pd > 0 Per valori di P molto piccoli, r(P) può essere negativo (depensazione non critica) oppure positivo (depensazione critica) Pd = -100 Pd = 100
 può essere negativo (depensazione non critica) oppure positivo (depensazione critica) Pd = Pd = 100.")
28
con effetto Allee Evoluzione della popolazione
c è una costante legata al tasso malthusiano di crescita Depensazione non critica Depensazione critica

29
con effetto Allee PUNTI DI EQUILIBRIO
I due casi di depensazione critica e non critica si rivelano molto diversi nella ricerca dei punti di equilibrio. Depensazione critica Depensazione non critica

30
Depensazione non critica:
K capacità portante Depensazione non critica: Depensazione critica La stabilità dei punti di equilibrio dipende dai valori numerici che specificano la parabola, cioè dalla pendenza della curva Depensazione non critica: P = 0 instabile P = K dipende dai parametri P = stabile possibile estinzione P = Pd instabile P = K dipende dai parametri Depensazione critica:

31
UN ESEMPIO Il branco di lupi
Si consideri una popolazione di lupi con capacità portante K=1000 e tasso di crescita descritto dalla legge di depensazione. Supponiamo c=10^(-6) e Pd = 100 (depensazione critica) o Pd = -100 (depensazione non critica). Simuliamo l’evoluzione della popolazione su 50 anni
 e Pd = 100 (depensazione critica) o. Pd = -100 (depensazione non critica). Simuliamo l’evoluzione della popolazione su 50 anni.")
32
Nd = -100 Nd = 100

33
Nel caso critico una popolazione di lupi con numerosità inferiore a 100 (Nd) si estingue
 si estingue")
34
Si supponga che venga concessa la caccia.
E’ consentita l’abbattimento di 200 esemplari ogni anno a partire dall’undicesimo anno. caccia L’effetto del prelievo è evidente, ma alla fine del periodo venatorio la popolazione è in grado di recuperare.

35
Anche in questo caso l’effetto della caccia si fa sentire, ma la popolazione è in grado di recuperare. Ma se la caccia è permessa anche all’anno 31, alla fine del periodo venatorio la popolazione si trova sotto il valore Nd, ciò provoca l’inesorabile estinzione anche se la caccia viene sospesa.

36
La regolamentazione di una popolazione con depensazione critica è molto delicata !
NON E’ NECESSARIO STERMINARE TUTTI GLI INDIVIDUI PER INDURRE UNA POPOLAZIONE ALL’ESTINZIONE …

Presentazioni simili













>")





