Scaricare la presentazione
1
Studio della metabolismo ossidativo della Dopamina

2
Neuromelanin formation
Neuromelanin formation. The pathway of the reactions converting dopamine to neuromelanin is a normal process in the substantia nigra, which it is tought to involve several steps:1) dopamine oxidation to dopamine o-quinone catalyzed by metals, oxygen, peroxynitrite or peroxidative activity of several enzymes (PX), such as prostaglandin H synthase, cytochrome P450, xantine oxidase 2) cyclization of dopamine o-quinone to dopaminochrome via an addition at physiological pH values, leading to the formation of unstable leukodopaminochrome and oxidation of leukodopaminochrome to dopaminochrome and polymerization of dopaminochrome to neuromelanin. (see the text)
 dopamine oxidation to dopamine o-quinone catalyzed by metals, oxygen, peroxynitrite or peroxidative activity of several enzymes (PX), such as prostaglandin H synthase, cytochrome P450, xantine oxidase 2) cyclization of dopamine o-quinone to dopaminochrome via an addition at physiological pH values, leading to the formation of unstable leukodopaminochrome and oxidation of leukodopaminochrome to dopaminochrome and polymerization of dopaminochrome to neuromelanin. (see the text)")
3
LOCALIZZAZIONE ATTIVITA’ DOPAMINO PEROSSIDASICA IN CERVELLO DI RATTO
E UMANO * * *

4
Studio in vitro della formazione del dopaminocromo
La formazione di dopaminocromo è catalizzata in maniera simile da perossidasi purificate (lattoperossidasi, mieloperossidasi, perossidasi di rafano) e da una frazione di cervello di ratto, in presenza di perossido d’idrogeno. La formazione di un intermedio radicalico, dopamino-o-semichinone, è comune nelle reazioni catalizzate da ciascuna delle perossidasi e dalla frazione di cervello di ratto. (Galzigna et al. Biochim Biophys Acta 1999)
 e da una frazione di cervello di ratto, in presenza di perossido d’idrogeno. La formazione di un intermedio radicalico, dopamino-o-semichinone, è comune nelle reazioni catalizzate da ciascuna delle perossidasi e dalla frazione di cervello di ratto. (Galzigna et al. Biochim Biophys Acta 1999)")
5
Procedures Preparation of protein mixture: The midbrain fractions were homogenized and centrifuged. The supernatant, obtained after two successive centrifugations, was spectrophotometrically assayed for enzymatic activity. striato s.nigra Assay of peroxidizing activity Dopamine peroxidizing activity was in vitro followed spectrophometrically, as increasing absorbance at 475nm, corresponding to the peak of maximum absorption of the dopaminochrome formed from dopamine and hydrogen peroxyde. This interest comes from the demonstration of an enzymatic activity catalyzing the formation of dopaminochrome - from dopamine and hydrogen peroxide - firstly described in a rat brain fraction (11) and then found by us in human midbrain (12). Subsequentely, we observed an increased dopamine peroxidation in the midbrain and basal ganglia of Parkinsonian brain, obtained at autopsy (13). In order to isolate the fraction of brain homogenate containing the DA peroxidizing activity, we have investigated non-denaturing polyacrylamide gel electrophoresis conditions, mimicking the characteristic colour in vitro reaction, resulting from cyclic oxidation of DA
 and then found by us in human midbrain (12). Subsequentely, we observed an increased dopamine peroxidation in the midbrain and basal ganglia of Parkinsonian brain, obtained at autopsy (13). In order to isolate the fraction of brain homogenate containing the DA peroxidizing activity, we have investigated non-denaturing polyacrylamide gel electrophoresis conditions, mimicking the characteristic colour in vitro reaction, resulting from cyclic oxidation of DA.")
6
Come si determina l’attività enzimatica
ΔA475 rappresenta la variazione di assorbanza misurata a 475 nm; ε è il coefficiente di estinzione molare del substrato cromoforo = 1175 M-1cm-1 La determinazione è stata effettuata in un il volume totale della miscela (omogenato e tampone di lettura) di 10-3 L (Galzigna et al., 1999)
 di 10-3 L (Galzigna et al., 1999)")
7
Attività dopamino perossidasica in mesencefalo
ATTIVITA’ DOPAMINO PEROSSIDASICA IN MESENCEFALO E GANGLI DELLA BASE DA REPERTI AUTOPTICI PARKINSONIANI E CONTROLLI Attività dopamino perossidasica in mesencefalo Controlli (media dei valori) = 249 (SD+/-191) mmol dopaminocromo/min/mg proteina PD (media dei valori) = 789 (SD+/-149) mmol dopaminocromo/min/mg proteina Attività dopamino perossidasica nei gangli della base Controlli (media dei valori) = 0 mmol dopaminocromo/min/mg proteina PD (media dei valori) = 529 mmol dopaminocromo/min/mg proteina *Significativamente differente dai controlli, p<0,001 con t di Student
 = 249 (SD+/-191) mmol dopaminocromo/min/mg proteina. PD (media dei valori) = 789 (SD+/-149) mmol dopaminocromo/min/mg proteina. Attività dopamino perossidasica nei gangli della base. Controlli (media dei valori) = 0 mmol dopaminocromo/min/mg proteina. PD (media dei valori) = 529 mmol dopaminocromo/min/mg proteina. *Significativamente differente dai controlli, p<0,001 con t di Student.")
8
CONCENTRAZIONE DELLE CATECOLAMINE IN MESENCEFALO E GANGLI DELLA BASE
Cconcentrazione della dopamina nel mesencefalo Ccontrolli (media dei valori) = 62 nmol/L (SD+/-37) PD (media dei valori) = 31 nmol/L (SD+/-26) Cconcentrazione della dopamina nei gangli della base Ccontrolli (media dei valori) = 59 nmol/L (SD+/-35) PD (media dei valori) = 43 nmol/L (SD+/-22) *Significativamente differenti dal controllo : p< 0.05 con t di Student.
 = 62 nmol/L (SD+/-37) PD (media dei valori) = 31 nmol/L (SD+/-26) Cconcentrazione della dopamina nei gangli della base. Ccontrolli (media dei valori) = 59 nmol/L (SD+/-35) PD (media dei valori) = 43 nmol/L (SD+/-22) *Significativamente differenti dal controllo : p< 0.05 con t di Student.")
9
PROCEDURE ELETTROFORETICHE

10
Procedure elettroforetiche
Gel di poliacrilamide non denaturante: è utilizzato per il frazionamento di miscele proteiche che mantengono la loro conformazione nativa. Esso non permette di distinguere gli effetti delle dimensioni, della struttura e della carica elettrica netta di ciascuna proteina sulla mobilità elettroforetica. E’ utilizzato soprattutto quando si voglia studiare l’attività biologica di una proteina.

11
Procedure elettroforetiche
Gel di poliacrilamide denaturante in presenza di sodio dodecilsolfato (SDS-PAGE). Prevede la denaturazione delle miscele proteiche al calore e il trattamento con agenti riducenti (2-α-mercaptoetanolo), che rompe i ponti disolfuri intra ed intercatena, causando lo “srotolamento” della struttura ripiegata della catena. La presenza dell’SDS assicura prolungata denaturazione, rompendo i legami covalenti, e, avvolgendo la proteina, le conferisce carica negativa, indipendentemente dalla sua carica netta. Tutte le proteine della miscela migrano verso l’anodo, risolte in bande discrete, con una velocità inversamente proporzionale al logaritmo del loro peso molecolare. Si attribuisce peso molecolare alle bande discrete mediante confronto della loro mobilità elettroforetica con quella di alcune proteine a peso molecolare noto
. Prevede la denaturazione delle miscele proteiche al calore e il trattamento con agenti riducenti (2-α-mercaptoetanolo), che rompe i ponti disolfuri intra ed intercatena, causando lo srotolamento della struttura ripiegata della catena. La presenza dell’SDS assicura prolungata denaturazione, rompendo i legami covalenti, e, avvolgendo la proteina, le conferisce carica negativa, indipendentemente dalla sua carica netta. Tutte le proteine della miscela migrano verso l’anodo, risolte in bande discrete, con una velocità inversamente proporzionale al logaritmo del loro peso molecolare. Si attribuisce peso molecolare alle bande discrete mediante confronto della loro mobilità elettroforetica con quella di alcune proteine a peso molecolare noto.")
12
Q-mass spectrometry analysis
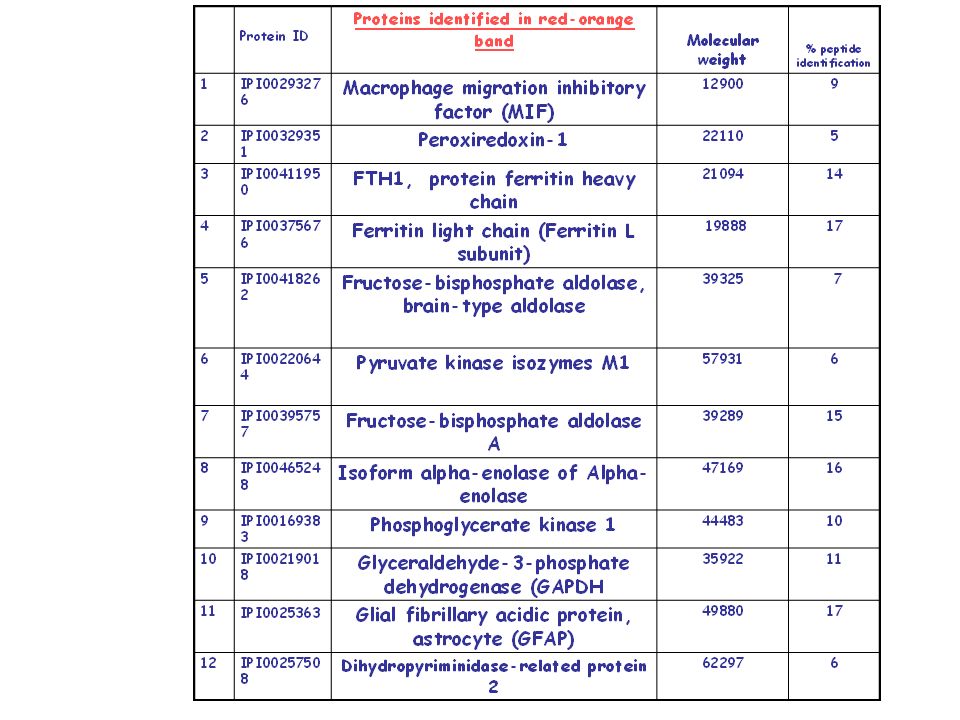
Protein mixtures of midbrain tissues homogenates from four different specimens were separated by non-denaturing polyacrylamide gel electrophoresis, in two corresponding gel sets The protein mixtures were separated along with horse radish peroxidase, which was present as peroxidatic actvity control. After electrophoresis running, one gel set was stained with the substrate solution (2mM DA, 30mM H2O2) and the second control gel set was stained with Coomassie Blue. Separation of protein mixture Q-mass spectrometry analysis A red/orange band diplaying peroxidatic activity and the corresponding band stained with Coomassie Blue were analysed by mass spectrometry.
 and the second control gel set was stained with Coomassie Blue. Separation of protein mixture. Q-mass spectrometry analysis. A red/orange band diplaying peroxidatic activity and the corresponding band stained with Coomassie Blue were analysed by mass spectrometry.")
13
DOPAMINE PEROXIDIZING ACTIVITY: DETECTION ON
NON-DENATURING POLYACRYLAMIDE GEL A A A’ HRP HRP 1’ ‘ 3’ 4’ HRP

15
Proteins identified in colorless, non-reactive band
Protein ID Protein name Molecular Weight % Peptide identification 1 IPI Malate dehydrogenase, cytoplasmic 36426 7 2 IPI Isoform 1 of Alpha-adducin 80955 8 3 IPI Retinal dehydrogenase- 1 54862 11 4 IPI IGKV1-5 protein 26234 18

16
Conclusions We have developed a method for detecting on native gel a dopamine peroxidizing activity from human midbrain. This method appears to be the first report of a peroxidatic activity in gel detection using dopamine and hydrogen peroxide as substrates. Q-mass spectrometry analysis of the activity bands revealed the presence, among the others, of two proteins: macrophage migration inhibitory factor (MIF) and Peroxiredoxin-1, highlighting a possible functional link among dopamine/dopaminochrome redox cycle and protein metabolism.
 and Peroxiredoxin-1, highlighting a possible functional link among dopamine/dopaminochrome redox cycle and protein metabolism.")
17
PERSPECTIVES Our findings, revealing a possible functional link among oxidative species and protein metabolism, are consistent with the latest studies on the pathogenic mechanism of PD. The properties of MIF and Peroxiredoxin-1 present in the activity bands are consistent with their role in the maintenance of redox potential within cells. New experimental approaches are under study in order to define the role of proteins found in the gel activity band, in particular of MIF and Peroxiredoxin-1, in oxidative metabolism of dopamine and in PD. Recently, MIF has been reported to possess a thiol-protein oxidoreductase activity and to play a role in cellular redox signalling. Moreover, dopachrome tautomerase activity, which converts D-dopachrome methylester to 5,6 dihydroxyndole-2-carboxymethylester, was described for MIF. The Peroxiredoxins members of a new expanding family of thiol-specific antioxidant proteins, using peroxidase-like mechanism, appear to be responsible for the redox regulation of diverse metabolic processes. Moreover, specific protein complex between MIF and PAG, a peroxiredoxin was shown. The evidence for the occurrence of oxidative stress in PD is overwhelming, and there is little doubt that oxidative stress leads to an increase in oxidative damage in the substantia nigra. Investigation of oxidative species and protein metabolism shows a complex cycle of events occurs between these two processes that appear to be inevitably linked and integrated. Nevertheless, the conundrum remains that is not currently possible to separate out oxidative stress from the other components of the cascade of events leading to the dopaminergic cell degeneration.

18
Cos’è la PROTEOMICA ? La proteomica è una disciplina scientifica
che permette lo studio del PROTEOMA, cioè delle PROTeine espresse da un genOMA in una particolare cellula, tessuto o organismo, sia esso animale o vegetale

21
Cos’è la SPETTROMETRIA DI MASSA
E’una tecnica analitica che permette di determinare la massa molecolare di un composto chimico. Ha origine nella prima metà del 1900, grazie agli studi di J. Thompson, il quale osservò che, in un tubo sotto vuoto a cui venga applicata una differenza di potenziale, si formano elettroni e radiazioni positive. Uno spettrometro di massa è uno strumento che misura la massa molecolare di una molecola, dopo che gli sia stata impartita una carica elettrica. Esso è, infatti, in grado di separare gli ioni molecolari in base al loro rapporto massa/carica.

22
MALDI MALDI-Tof MALDI, matrix.assisted laser desorption/ionization desorbimento: è un processo in cui una miscela viene evaporata da una superficie e ionizzata. Piastra maldi è un supporto d’acciaio: il campione dopo dogestione con tripsina e immerso in una matrice (acido organico) che funziona da solvente eminimizza le interazioni molecolari, assorbe parte dell’energia che viene impartita, ed ha un ruolo attivo anche nella formazione dell’evaporazione e formazione degli ioni per protonazione, cioè assorbono una carica positiva. Il campione depositatato sulla piastra viene lasciato cristalllizzare e bombardata con fotoni ad alta energia, proveneienti da raggio laser pulsato, quinid vengiono diretti verso l’analizzatore tof, che è un tubo, di lunghezza nota’in cui viene fatto il vuoto. Gli ioni vengono emessi ad un’energia cinetica costante ed indirizzati verso il tubo. E= i\e mv2, dove m è la massa dello ione e v la sua velocità. Minore sarà il rapporo massa\carica maggiore sarà la sua velocità.
 che funziona da solvente eminimizza le interazioni molecolari, assorbe parte dell’energia che viene impartita, ed ha un ruolo attivo anche nella formazione dell’evaporazione e formazione degli ioni per protonazione, cioè assorbono una carica positiva. Il campione depositatato sulla piastra viene lasciato cristalllizzare e bombardata con fotoni ad alta energia, proveneienti da raggio laser pulsato, quinid vengiono diretti verso l’analizzatore tof, che è un tubo, di lunghezza nota’in cui viene fatto il vuoto. Gli ioni vengono emessi ad un’energia cinetica costante ed indirizzati verso il tubo. E= i\e mv2, dove m è la massa dello ione e v la sua velocità. Minore sarà il rapporo massa\carica maggiore sarà la sua velocità.")
23
Spettrometria di massa
La spettrometria di massa permette di determinare la massa molecolare di un composto chimico.( ha origine da Thompson nel 1900: in un tubo in cui venga fatto il vuoto ed al quale venga applicata una differenza di potenziale si generano elettroni e radiazioni positive) Lo spettrometro di massa è uno strumento analitico in grado di misurare la massa molecolare di una molecola, dopo che sia stata ionizzata, dopo cioè che gli sia stata impartita una carica elettrica. Esso è infatti in grado di separare gli ioni molecolari in base al rapporto di massa/carica. Prima dell’immisione nello spettrometro di massa il campione una volat doveva passare in fase gassosa. , oggi può essere in fase liquida, solida o gassosa ed esser introdotto in una camera dove si fa il vuoto. MALDI0 matrix.assisted laser desorption/ionization desorbimento: è un processo in cui una miscela viene evaporata da una superficie e ionizzata. Piastra maldi è un supporto d’acciaio: il campione dopo dogestione con tripsina e immerso in una matrice (acido organico) che funziona da solvente eminimizza le interazioni molecolari, assorbe parte dell’energia che viene impartita, ed ha un ruolo attivo anche nella formazione dell’evaporazione e formazione degli ioni per protonazione, cioè assorbono una carica positiva. Il campione depositatato sulla piastra viene lasciato cristalllizzare e bombardata con fotoni ad alta energia, proveneienti da raggio laser pulsato, quinid vengiono diretti verso l’analizzatore tof, che è un tubo, di lunghezza nota’in cui viene fatto il vuoto. Gli ioni vengono emessi ad un’energia cinetica costante ed indirizzati verso il tubo. E= i\e mv2, dove m è la massa dello ione e v la sua velocità. Minore sarà il rapporo massa\carica maggiore sarà la sua velocità.
 Lo spettrometro di massa è uno strumento analitico in grado di misurare la massa molecolare di una molecola, dopo che sia stata ionizzata, dopo cioè che gli sia stata impartita una carica elettrica. Esso è infatti in grado di separare gli ioni molecolari in base al rapporto di massa/carica. Prima dell’immisione nello spettrometro di massa il campione una volat doveva passare in fase gassosa. , oggi può essere in fase liquida, solida o gassosa ed esser introdotto in una camera dove si fa il vuoto. MALDI0 matrix.assisted laser desorption/ionization desorbimento: è un processo in cui una miscela viene evaporata da una superficie e ionizzata. Piastra maldi è un supporto d’acciaio: il campione dopo dogestione con tripsina e immerso in una matrice (acido organico) che funziona da solvente eminimizza le interazioni molecolari, assorbe parte dell’energia che viene impartita, ed ha un ruolo attivo anche nella formazione dell’evaporazione e formazione degli ioni per protonazione, cioè assorbono una carica positiva. Il campione depositatato sulla piastra viene lasciato cristalllizzare e bombardata con fotoni ad alta energia, proveneienti da raggio laser pulsato, quinid vengiono diretti verso l’analizzatore tof, che è un tubo, di lunghezza nota’in cui viene fatto il vuoto. Gli ioni vengono emessi ad un’energia cinetica costante ed indirizzati verso il tubo. E= i\e mv2, dove m è la massa dello ione e v la sua velocità. Minore sarà il rapporo massa\carica maggiore sarà la sua velocità.")
24
Fragment fingerprinting del peptide selezionato: la sequenza parziale, insieme con la massa, costituisce un “tag” di sequenza che può essere utilizzato come probe, altamente specifico, per identificare proteine nei database proteici. Lo spettro di frammentazione può anche essere analizzato automaticamente per mezzo di SEQUEST, un programma che consente la correlazione dei dati sperimentali con spettri teorici, generati da sequenze proteiche note presenti nei database.

25
Procedure elettroforetiche
L’ elettroforesi BIDIMENSIONALE prevede due fasi: IEF: Separazione delle proteine in base alle differenze nella loro carica netta mediante l’isoelettrofocalizzazione SDS-PAGE: Separazione delle proteine focalizzate in base alla loro massa molecolare, in gel denaturante

26
Modello di ratto emiparkinsoniano
La somministrazione di 6-OHDA riproduce, nel ratto, fenomeni di stress ossidativo determinati dalla riduzione dei complessi I e IV della catena respiratoria con un’iperproduzione mitocondriale di radicali liberi, quali lo ione superossido, di radicali idrossilici e di perossido d’idrogeno. Questa riduzione dell’attività dei complessi I e IV mitocondriali è probabilmente alla base della perdita di neuroni che si realizza, a livello della substantia nigra trattata, nel giro di due settimane. (A e B). A: denervazione dello striato ipsolaterale B: danno neuronale in s.nigra, dopo iniezione di 6-idrossidopamina (6-OHDA)
. A: denervazione dello striato ipsolaterale. B: danno. neuronale in s.nigra, dopo iniezione di 6-idrossidopamina. (6-OHDA)")
27
CONCLUSIONI IL protocollo di estrazione e solubilizzazione delle proteine, messo a punto per il tessuto cerebrale, associato ad un programma mirato di isoelettrofocalizzazione, ci ha permesso di ottenere una soddisfacente risoluzione del corredo proteico delle sezioni di substantia nigra e di striato mediante separazione in doppia dimensione. Significative variazioni di espressione di alcune proteine sono emerse dal confronto, effettuato mediante software d’immagine, dei profili proteici delle sezioni di tessuto neurodegenerato rispetto al controllo. Le proteine, individuate come significativamente differenti nei tessuti patologici rispetto ai controlli, sono state sequenziate in spettrometria di massa MALDI-TOF. Nel campione di tessuto ottenuto dallo striato due spot che appaiono piu’ intensamente espressi nel profilo proteico dello striato indotto alla neurodegenerazione, rispetto al controllo, corrispondono alla b-actina. Lo spot proteico presente nel campione di substantia nigra trattato con 6-OHDA e non nel rispettivo controllo corrisponde ad un’enzima: l’a-enolasi ( 2-fosfo-D-glicerato idrolasi) (non neuronal-enolasi) (NNE). L’ aumento dei livelli di espressione dell’a-enolasi e della b-actina, nei tessuti neurodegenerati rispetto al tessuto di controllo, potrebbe essere risultato dei fenomeni di ossidazione indotti dalla 6-OHDA. Entrambe le proteine appaiono coinvolte nei processi di neurodegenerazione ed è testimoniata l’ossidazione che esse subiscono nella malattia di Alzheimer. ( Castagna et al.,2002). Il coinvolgimento di fenomeni di stress ossidativo nella patogenesi del morbo di Parkinson è infatti largamente dimostrato (Jenner et al. AnnNeurol, 2003; 53 suppl. S26-S38). I nostri risultati sono pertanto ricollocabili all’interno di tale scenario. Se l’a-enolasi e la b-actina possono essere considerate bersagli del processo di ossidazione che avviene a carico di molteplici proteine, nelle malattie neurodegenerative quali il morbo di Parkinson e la malattia d’Alzheimer, potrebbero rappresentare importanti indicatori diagnostici, sebbene occorra stabilire in quale fase della malattia insorgano le alterazioni osservate, per far luce sul fattore o i fattori d’innesco della neurodegenerazione. Solo in questo modo, infatti, sarà possibile risolvere il nesso di causalità della malattia, ancora purtroppo oscuro. Le mutazioni geniche associate alla malattia hanno fornito importanti indicazioni sul probabile meccanismo molecolare coinvolto nella patogenesi della neurodegenerazione, avvalorando le ipotesi riguardanti lo stress ossidativo e le disfunzioni mitocondriali, basate sulle osservazioni epidemiologiche e sperimentali. La presenza dell’a-enolasi e della b-actina, nella nostra frazione proteica proveniente rispettivamente dal tessuto di substantia nigra e da quello dello striato, trova spiegazione in letteratura. Entrambe le proteine appaiono coinvolte nei processi di neurodegenerazione ed è testimoniata l’ossidazione che esse subiscono nella malattia di Alzheimer. ( Castagna et al.,2002). L’a-enolasi è una delle subunità che compongono l’enzima enolasi, che catalizza la reazione di rimozione di una molecola di acqua dal 2-fosfoglicerato, producendo fosfoenoilpiruvato nella via metabolica della glicolisi. L’enzima attivo è un dimero che comprende tre differenti subunità, denominate a, b e g. Gli omodimeri aa, bb e g e gli eterodimeri ag e ab sono stati trovati nei tessuti umani. Le isoforme ag e gg sono i dimeri predominanti nel cervello e rappresentano le enolasi specifiche dei neuroni, mentre le altre isoforme sono presenti soprattutto nel cuore e nel muscolo scheletrico. La subunità g è localizzata nei neuroni, mentre la subunità a è localizzata nelle cellule gliali. Studi sulla distribuzione dell’attività enolasica nel cervello umano non hanno mostrato differenze significative nei livelli delle specifiche isoforme, rispetto all’età, sebbene sia stata dimostrata la trasformazione dell’isoforma a in quella g, durante lo sviluppo cerebrale. Uno studio proteomico quantitativo della malattia di Alzheimer ha dimostrato un aumento dei livelli della subunità a nel cervello umano malato rispetto a quello sano ( Schonberger et al., 2001).In tale studio, infatti, è risultato come l’a-enolasi sia ossidata nel tessuto malato rispetto a quello sano e questo ha suggerito un possibile legame tra alterazione degli enzimi che presiedono alla glicolisi e diminuito metabolismo del glucosio a livello cerebrale nella malattia di Alzheimer, con conseguente accumulo degli enzimi glicolitici nel cervello ( Butterflie DA 2001). L’accumulo dell’a-enolsi, che aumenta nella malattia di Alzheimer, potrebbe essere il risultato di una modificazione ossidativa degli amminoacidi, con perdita della funzionalità della proteina stessa e ridotta capacità di eliminazione della medesima da parte delle proteinasi. La presenza di una sovraespressione di b-actina nel profilo dello striato lesionato trova anch’essa spiegazione in letteratura. Nel sistema nervoso centrale l’actina è largamente distribuita nei neuroni, negli astrociti e nei vasi sanguigni. Alcuni studi dimostrano la sua presenza soprattutto nelle porzioni periferiche del corpo neuronale , comprese le proiezioni assonali ( Pannese et al., Anat. Embryol. 1996; 194, ). Anche nel caso della b-actina l’incremento di espressione potrebbe essere il risultato di un’ossidazione della stessa, già riscontrata in altre malattie neurodegenerative (Castegna et al., J Neurochem. 2002; 82, ) . E’stato osservato, infatti, che fenomeni di stress ossidativo inattivano la b-actina. (Milzani et al., Biochem Biophys. 1997; 339, ; Milzani et al., Fasej.1997Mar;11(4): ). La conseguenza è un non corretto assemblaggio dei filamenti di actina e, pertanto, la perdita, da parte del citoscheletro, della sua struttura. Le mutazioni geniche associate alla malattia hanno fornito importanti indicazioni sul probabile meccanismo molecolare coinvolto nella patogenesi della neurodegenerazione, avvalorando le ipotesi riguardanti lo stress ossidativo e le disfunzioni mitocondriali, basate sulle osservazioni epidemiologiche e sperimentali. Nel morbo di Parkinson i fenomeni di stress ossidativo e le alterazioni mitocondriali possono influenzare l’espressione delle proteine ed i meccanismi implicati nelle mutazioni geniche che causano la malattia, così le mutazioni geniche, d’altra parte, possono portare allo scompenso delle funzioni mitocondriali ed allo stress ossidativo. L’analisi degli stessi profili proteici, dopo trattamento degli animali con molecole antiossidanti, sarebbe un interessante, conseguente studio, a quello condotto nel corso di questa tesi. L’osservazione, infatti, di una non mutata espressione delle proteine a-enolasi e b-actina nei tessuti trattati con 6-OHDA, conforterebbe la nostra ipotesiLo stesso studio, mediante approccio proteomico, potrebbe essere condotto nell’uomo, utilizzando marcatori periferici. La proteomica potrebbe essere il piu’ valido strumento di identificazione degli stessi nel liquido cerebrospinale ad esempio, o nel siero.
 (non neuronal-enolasi) (NNE). L’ aumento dei livelli di espressione dell’a-enolasi e della b-actina, nei tessuti neurodegenerati rispetto al tessuto di controllo, potrebbe essere risultato dei fenomeni di ossidazione indotti dalla 6-OHDA. Entrambe le proteine appaiono coinvolte nei processi di neurodegenerazione ed è testimoniata l’ossidazione che esse subiscono nella malattia di Alzheimer. ( Castagna et al.,2002). Il coinvolgimento di fenomeni di stress ossidativo nella patogenesi del morbo di Parkinson è infatti largamente dimostrato (Jenner et al. AnnNeurol, 2003; 53 suppl. S26-S38). I nostri risultati sono pertanto ricollocabili all’interno di tale scenario. Se l’a-enolasi e la b-actina possono essere considerate bersagli del processo di ossidazione che avviene a carico di molteplici proteine, nelle malattie neurodegenerative quali il morbo di Parkinson e la malattia d’Alzheimer, potrebbero rappresentare importanti indicatori diagnostici, sebbene occorra stabilire in quale fase della malattia insorgano le alterazioni osservate, per far luce sul fattore o i fattori d’innesco della neurodegenerazione. Solo in questo modo, infatti, sarà possibile risolvere il nesso di causalità della malattia, ancora purtroppo oscuro. Le mutazioni geniche associate alla malattia hanno fornito importanti indicazioni sul probabile meccanismo molecolare coinvolto nella patogenesi della neurodegenerazione, avvalorando le ipotesi riguardanti lo stress ossidativo e le disfunzioni mitocondriali, basate sulle osservazioni epidemiologiche e sperimentali. La presenza dell’a-enolasi e della b-actina, nella nostra frazione proteica proveniente rispettivamente dal tessuto di substantia nigra e da quello dello striato, trova spiegazione in letteratura. Entrambe le proteine appaiono coinvolte nei processi di neurodegenerazione ed è testimoniata l’ossidazione che esse subiscono nella malattia di Alzheimer. ( Castagna et al.,2002). L’a-enolasi è una delle subunità che compongono l’enzima enolasi, che catalizza la reazione di rimozione di una molecola di acqua dal 2-fosfoglicerato, producendo fosfoenoilpiruvato nella via metabolica della glicolisi. L’enzima attivo è un dimero che comprende tre differenti subunità, denominate a, b e g. Gli omodimeri aa, bb e g e gli eterodimeri ag e ab sono stati trovati nei tessuti umani. Le isoforme ag e gg sono i dimeri predominanti nel cervello e rappresentano le enolasi specifiche dei neuroni, mentre le altre isoforme sono presenti soprattutto nel cuore e nel muscolo scheletrico. La subunità g è localizzata nei neuroni, mentre la subunità a è localizzata nelle cellule gliali. Studi sulla distribuzione dell’attività enolasica nel cervello umano non hanno mostrato differenze significative nei livelli delle specifiche isoforme, rispetto all’età, sebbene sia stata dimostrata la trasformazione dell’isoforma a in quella g, durante lo sviluppo cerebrale. Uno studio proteomico quantitativo della malattia di Alzheimer ha dimostrato un aumento dei livelli della subunità a nel cervello umano malato rispetto a quello sano ( Schonberger et al., 2001).In tale studio, infatti, è risultato come l’a-enolasi sia ossidata nel tessuto malato rispetto a quello sano e questo ha suggerito un possibile legame tra alterazione degli enzimi che presiedono alla glicolisi e diminuito metabolismo del glucosio a livello cerebrale nella malattia di Alzheimer, con conseguente accumulo degli enzimi glicolitici nel cervello ( Butterflie DA 2001). L’accumulo dell’a-enolsi, che aumenta nella malattia di Alzheimer, potrebbe essere il risultato di una modificazione ossidativa degli amminoacidi, con perdita della funzionalità della proteina stessa e ridotta capacità di eliminazione della medesima da parte delle proteinasi. La presenza di una sovraespressione di b-actina nel profilo dello striato lesionato trova anch’essa spiegazione in letteratura. Nel sistema nervoso centrale l’actina è largamente distribuita nei neuroni, negli astrociti e nei vasi sanguigni. Alcuni studi dimostrano la sua presenza soprattutto nelle porzioni periferiche del corpo neuronale , comprese le proiezioni assonali ( Pannese et al., Anat. Embryol. 1996; 194, ). Anche nel caso della b-actina l’incremento di espressione potrebbe essere il risultato di un’ossidazione della stessa, già riscontrata in altre malattie neurodegenerative (Castegna et al., J Neurochem. 2002; 82, ) . E’stato osservato, infatti, che fenomeni di stress ossidativo inattivano la b-actina. (Milzani et al., Biochem Biophys. 1997; 339, ; Milzani et al., Fasej.1997Mar;11(4): ). La conseguenza è un non corretto assemblaggio dei filamenti di actina e, pertanto, la perdita, da parte del citoscheletro, della sua struttura. Le mutazioni geniche associate alla malattia hanno fornito importanti indicazioni sul probabile meccanismo molecolare coinvolto nella patogenesi della neurodegenerazione, avvalorando le ipotesi riguardanti lo stress ossidativo e le disfunzioni mitocondriali, basate sulle osservazioni epidemiologiche e sperimentali. Nel morbo di Parkinson i fenomeni di stress ossidativo e le alterazioni mitocondriali possono influenzare l’espressione delle proteine ed i meccanismi implicati nelle mutazioni geniche che causano la malattia, così le mutazioni geniche, d’altra parte, possono portare allo scompenso delle funzioni mitocondriali ed allo stress ossidativo. L’analisi degli stessi profili proteici, dopo trattamento degli animali con molecole antiossidanti, sarebbe un interessante, conseguente studio, a quello condotto nel corso di questa tesi. L’osservazione, infatti, di una non mutata espressione delle proteine a-enolasi e b-actina nei tessuti trattati con 6-OHDA, conforterebbe la nostra ipotesiLo stesso studio, mediante approccio proteomico, potrebbe essere condotto nell’uomo, utilizzando marcatori periferici. La proteomica potrebbe essere il piu’ valido strumento di identificazione degli stessi nel liquido cerebrospinale ad esempio, o nel siero.")
28
DOPAMINA

29
6-IDROSSIDOPAMINA

30
Confronto tra i profili proteici ottenuti da sezioni di tessuto
8 pH 5 striato Substantia nigra b-actina A: Neostriato controllo Neostriato trattato con 6-OHDA 7 pH 4 Il modello animale di morbo di Parkinson è stato ottenuto iniettando stereotassicamente la substantia nigra destra di ratti con 6-idrossidopamina (6-OHDA) tossina che determina stress ossidativo e pertanto neurodegenerazione, mantenendo la substantia nigra controlaterale come controllo (ratti emiparkinsionani). Campioni di tessuto di substantia nigra e di striato sono stati dissezionati da 5 ratti emiparkinsoniani, risultati positivi al test comportamentale con apomorfina, e successivamente omogenati. La miscela di proteine, presente nell’omogenato è stata sottoposta a separazione in doppia dimensione, mediante ief, e poi sds page. Abbaimo così ottenuto i profili proteici dei tessuti in neurodegenarzione edei rispettivi controlli.La separazione in prima dimensione del tessuto di striato è stata condotta in un gradiente di ph compreso tra 5 ed 8, quello dellas,nigra secondo un gradiente compreso tra 4 e 7. è.. L’analisi è stata effettuata mediante il software di analisi d’immagine PDQuest (Bio-Rad). Che ci ha permesso di osservare come proteine presenti nel profilo dello striato di cont. Risultavano maggiormente spresse nel tessuto trattato. Così alcune delle proteine nel profilo di nigra di controllo sembravano addirittutra scomparire nel ytrattato, ma al pari del profilo dello strato in neurodegenarzione anche in quello della nigra trattata deue spot proteici risultavano pienamente epsrisolti rispetto al controllo. La nostra cusriosità avviamente rsi è soofremata sul le oproteine maggiormente esperesse nei tesstu in neurodegenarzione e duno spot prioteico prelevato dal profilo delllo striato sequenzitato i spettrometrai di amssa maldi tof è risultatao corrispondente alla b,actina, quello dèprelevato dalla nigra tratttata è rilsutata corrisponedete allaalfa enolasi.alcunecon le frecce di colore rosso, che non compaiono nel tessuto trattato. idrolasi) (non neuronal-enolasi) (NNE). a-enolasi B: S. nigra controllo S. nigra trattata con 6-OHDA
 tossina che determina stress ossidativo e pertanto neurodegenerazione, mantenendo la substantia nigra controlaterale come controllo (ratti emiparkinsionani). Campioni di tessuto di substantia nigra e di striato sono stati dissezionati da 5 ratti emiparkinsoniani, risultati positivi al test comportamentale con apomorfina, e successivamente omogenati. La miscela di proteine, presente nell’omogenato è stata sottoposta a separazione in doppia dimensione, mediante ief, e poi sds page. Abbaimo così ottenuto i profili proteici dei tessuti in neurodegenarzione edei rispettivi controlli.La separazione in prima dimensione del tessuto di striato è stata condotta in un gradiente di ph compreso tra 5 ed 8, quello dellas,nigra secondo un gradiente compreso tra 4 e 7. è.. L’analisi è stata effettuata mediante il software di analisi d’immagine PDQuest (Bio-Rad). Che ci ha permesso di osservare come proteine presenti nel profilo dello striato di cont. Risultavano maggiormente spresse nel tessuto trattato. Così alcune delle proteine nel profilo di nigra di controllo sembravano addirittutra scomparire nel ytrattato, ma al pari del profilo dello strato in neurodegenarzione anche in quello della nigra trattata deue spot proteici risultavano pienamente epsrisolti rispetto al controllo. La nostra cusriosità avviamente rsi è soofremata sul le oproteine maggiormente esperesse nei tesstu in neurodegenarzione e duno spot prioteico prelevato dal profilo delllo striato sequenzitato i spettrometrai di amssa maldi tof è risultatao corrispondente alla b,actina, quello dèprelevato dalla nigra tratttata è rilsutata corrisponedete allaalfa enolasi.alcunecon le frecce di colore rosso, che non compaiono nel tessuto trattato. idrolasi) (non neuronal-enolasi) (NNE). a-enolasi. B: S. nigra controllo S. nigra trattata con 6-OHDA.")
31
pH pH pH pH7 Separazione miscela proteica di substantia nigra di ratto emiparkinsoniano mediante elettroforesi bidimensionale
















 www.javascript.com www.iboost.com/build/programming/js/tutorial/885.h tm www.javascriptsearch.com www.a1javascripts.com.>")

 è un enzima citoplasmatico con un'ampia distribuzione nei tessuti, dove catalizza.>")

>")


 E’ la morte cellulare programmata (origina dal nucleo), la ritroviamo in numerosi meccanismi fisiologici e patologici: - eliminazione di.>")