Scaricare la presentazione
1
Lez. 5 La ricombinazione sito-specifica La trasposizione del DNA

2
Tipi di ricombinazioni
Ricombinazione tra sequenze omologhe: (generalized o Homologous recombination) avviene prevalentemente nella meiosi. Avviene allo stadio di “four strand”. Ricombinazione tra paia di sequenze specifiche: site specific ricombination. Es. nell’integrazione dei fagi. Gli enzimi sono specifici per la sequenza Ricombinazione per trasposizione (Trasposizione): sequenze vengono inserite senza bisogno di omologia di sequenza
 avviene prevalentemente nella meiosi. Avviene allo stadio di four strand . Ricombinazione tra paia di sequenze specifiche: site specific ricombination. Es. nell’integrazione dei fagi. Gli enzimi sono specifici per la sequenza. Ricombinazione per trasposizione (Trasposizione): sequenze vengono inserite senza bisogno di omologia di sequenza.")
3
Ricombinazioni Ricombinazione conservativa sito-specifica (CSSR): tra due elementi di DNA definiti Ricombinazione per trasposizione (trasposizione): tra sequenze specifiche e siti non specifici Hanno in comune le ricombinasi e la formazione del complesso sinaptico
: tra sequenze specifiche. e siti non specifici. Hanno in comune le ricombinasi e la formazione del complesso sinaptico.")
4
CSSR (Ricombinazione conservativa sito-specifica)
Il segmento di DNA che viene spostato porta i siti di ricombinazione Esempio fago l un sito di ricombinazione sul DNA fagico e l’altro sul DNA batterico I siti di ricombinazione sono corti – 20 bp La CSSR può dar luogo a: Inserzione in un sito specifico (es. fago l) Delezione di un pezzo di DNA Inversione di un pezzo di DNA Ogni sito di ricombinazione ha: Due sequenze di riconoscimento della ricombinasi Una sequenza centrale: regione dello scambio
 Delezione di un pezzo di DNA. Inversione di un pezzo di DNA. Ogni sito di ricombinazione ha: Due sequenze di riconoscimento della ricombinasi. Una sequenza centrale: regione dello scambio.")
5
CSSR: tipi e meccanismo
I siti di ricombinazione sono composti da: sequenze di riconoscimento della ricombinasi e regione dello scambio Se hanno la stessa direzione (ripetizione diretta) permettono inserzione o delezione Se hanno direzione opposta (ripetizione invertita) permettono la inversione
 permettono inserzione o delezione. Se hanno direzione opposta (ripetizione invertita) permettono la inversione.")
6
Meccanismo delle ricombinasi: Ricombinasi a Serina
Una serina della ricombinasi attacca il P e libera il 3’OH. Il legame può essere riformato senza richiesta di energia (conservativo) Le ricombinasi a serina tagliano contemporaneamente i due filamenti di DNA prima dello scambio. Occorrono 4 subunità
 Le ricombinasi a serina tagliano contemporaneamente i due filamenti di DNA prima dello scambio. Occorrono 4 subunità.")
7
Ricombinasi a Tirosina
Le ricombinasi a tirosina tagliano ed uniscono due filamenti, e poi gli altri due. Si ha la formazione di una Holliday junction

8
Ricombinazione sito-specifica e toposomerasi
Le ricombinasi sono correlate alle topoisomerasi, e la reazione di ricombinazione somiglia quella di topoisomerase eccetto che gli strands da duplex diversi sono uniti insieme. La reazione conserva energia usando una tyrosine catalitica nell’enzima per rompere un legame fosfodiestere ed unire la terminazione al 3’. due unità enzimatiche si legano al sito di ricombinazione e i due dimeri formano un complesso in cui si ha il trasferimento.

9
Cre-Lox Un sistema semplice di ricombinasi a tirosina si trova nel batteriofago P1. La Cre recombinase codificata dal fago catalizza la ricombinazione tra due sequenze bersaglio. Le sequenze ricombinanti del fago P1 sono identiche, sono di 34 bp- chiamati loxP. La Cre ricombinase è sufficiente per la reazione; nessuna proteina accessoria è richiesta Per la sua semplicità ed efficienza, il sistema Cre/lox è stato adattato per le cellule eucariote, dove è diventata una tecnica standard per site-specific recombination Struttura di Cre

10
Cre-lox Ogni subunità di Cre si lega alla sequenza di riconoscimento.
Si ha un tetramero sul DNA cruciforme Cre ha due conformazioni quella verde può tagliare il DNA. Cambiando la coppia di subunità attive si taglia anche il secondo filamento

11
Ricombinazione fago l Il ciclo del fago lambda implica che:
Per essere lisogenico deve essere integrato nel DNA ospite Per entrare nel ciclo litico deve esser exciso dal cromosoma. I siti di integrazione (attP e attB) differiscono da quelli di excisione (attL e attR)
 differiscono da quelli di excisione (attL e attR)")
12
Integrasi di l Il passaggio tra lo stato lisogenico e la crescita litica del fago l richiede la integrazione o escissione del DNA fagico. L’integrasi l (lInt, a tirosina) catalizza la ricombinazione tra i siti attP (phage) e attB (batterio) attB di 30 bp ha i due siti di legame di lInt e la regione del crossing-over attP (240 bp) ha due braccia per il legame di lInt e del fattore di integrazione dell’ospite IHF (un fattore architettonico L’escissione richiede Xis
 catalizza la ricombinazione tra i siti attP (phage) e attB (batterio) attB di 30 bp ha i due siti di legame di lInt e la regione del crossing-over. attP (240 bp) ha due braccia per il legame di lInt e del fattore di integrazione dell’ospite IHF (un fattore architettonico. L’escissione richiede Xis.")
13
Ricombinasi Hin inverte il DNA
Nella Salmonella la ricombinasi Hin inverte una regione di 1000 bp fiancheggita da siti invertiti hixL e hixR In una posizione il promotore attiva i geni per la flagellina 2 e repressore per flagellina 1 Nell’altra induce flagellina 1 Il sistema necessita di un enhancer a DNA che lega la proteina Fis

14
Le resolvasi I DNA circolari dopo la ricombinazione omologa formano dimeri od oligomeri. Le ricombinasi chiamate resolvasi generano i monomeri In E. coli la resolvasi XerCD riconosce il sito dif Durante la replicazione cellulare è aiutata da FtsK, una ATPasi Xer è formato da 2 C e 2 D. C sono attive sempre, D solo in presenza di FtsK

15
La trasposizione Elementi trasponibili o trasposoni.
Il movimento avviene per un ricombinazione tra le estremità dell’elemento trasponibile e una sequenza di DNA della cellula. Tre classi Trasposoni a DNA Retrotrasposoni simili ai virus Retrotrasposoni poli-A

16
DNA mobile La seconda classe di DNA, interspersed DNA (moderately repeated DNA, Intermediate-repeat DNA). Fatta da un gran numero di poche famiglie di sequenze (45% hu genoma) Elementi mobili di DNA o elementi transposabili. Divisi in due categorie: DNA transposons e retrotransposons
 Elementi mobili di DNA o elementi transposabili. Divisi in due categorie: DNA transposons e retrotransposons.")
17
Trasposoni a DNA: taglia e incolla
Il DNA con ripetizioni invertite viene escisso, Il complesso sinaptico o trasposoma Nel DNA bersaglio viene fatta una nick, e si ha una trans-esterificazione: trasferimento del filamento di DNA.

18
Il transposoma La transposasi è codificata dal transposone
Occorrono almeno 2 subunità Ha il ruolo di riconoscere le estremità, unirle, tagliarle per fare il transposoma, ed inserire nel DNA bersaglio

19
Meccanismi di taglio del filamento non trasferito

20
IS911, a member of the IS3 family of bacterial insertion sequences, uses a two-step transposition mechanism. This entails formation of a circular intermediate which then undergoes integration. Complex A: the paired end complex (PEC) catalyses single-strand cleavage (hydrolysis) at one IS end and strand transfer to the other (trans-esterification). The closed circular copy is generated from this by replication and subsequently undergoes integration into a target DNA molecule using the highly recombinogenic IRR–IRL junction. The final integration event occurs within a second type of complex, synaptic complex B, which must be formed between the covalently joined inverted transposon ends and the target DNA. Formation of synaptic complex A requires the 44 kDa IS911 transposase, OrfAB, produced as a fusion protein. OrfAB contains the catalytic site in its C-terminal domain and alone, is capable of generating the bridged figure eight form. Pouget, N. et al. Nucl. Acids Res : ; doi: /nar/gkl420 Copyright restrictions may apply.
 catalyses single-strand cleavage (hydrolysis) at one IS end and strand transfer to the other (trans-esterification). The closed circular copy is generated from this by replication and subsequently undergoes integration into a target DNA molecule using the highly recombinogenic IRR–IRL junction. The final integration event occurs within a second type of complex, synaptic complex B, which must be formed between the covalently joined inverted transposon ends and the target DNA. Formation of synaptic complex A requires the 44 kDa IS911 transposase, OrfAB, produced as a fusion protein. OrfAB contains the catalytic site in its C-terminal domain and alone, is capable of generating the bridged figure eight form. Pouget, N. et al. Nucl. Acids Res : ; doi: /nar/gkl420. Copyright restrictions may apply.")
21
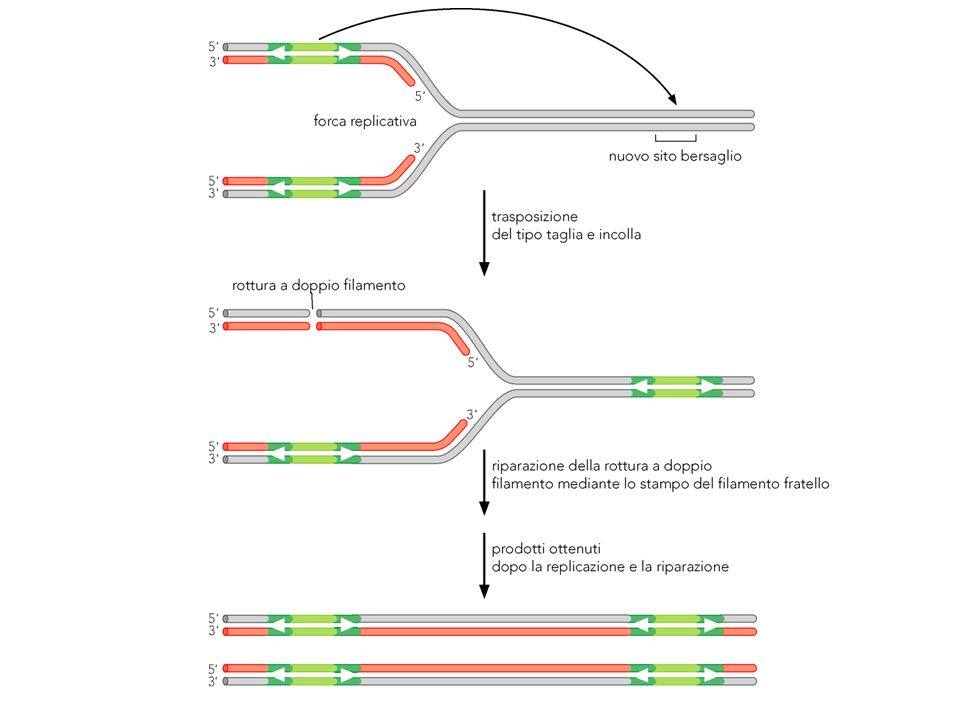
La transposizione a DNA con meccanismo replicativo
Dopo il trasferimento del filamento si forma una struttura con due ramificazioni, che può fungere da forca replicativa. La replicazione termina sulla seconda forca e produce due copie del transposone animazione

22
Retrotrasposoni simili ai virus
Hanno un RNA intermedio Il promotore è su LTR ed l’RNA viene copiato in cDNA Un’integrasi riconosce le estremità e dirige il trasposoma al bersaglio. Una trascrittasi inversa sintetizza il DNA

23
Modello dell retrotrascrizione di RNA retrovirale genomico in DNA

24
Retrotrasposoni poli-A
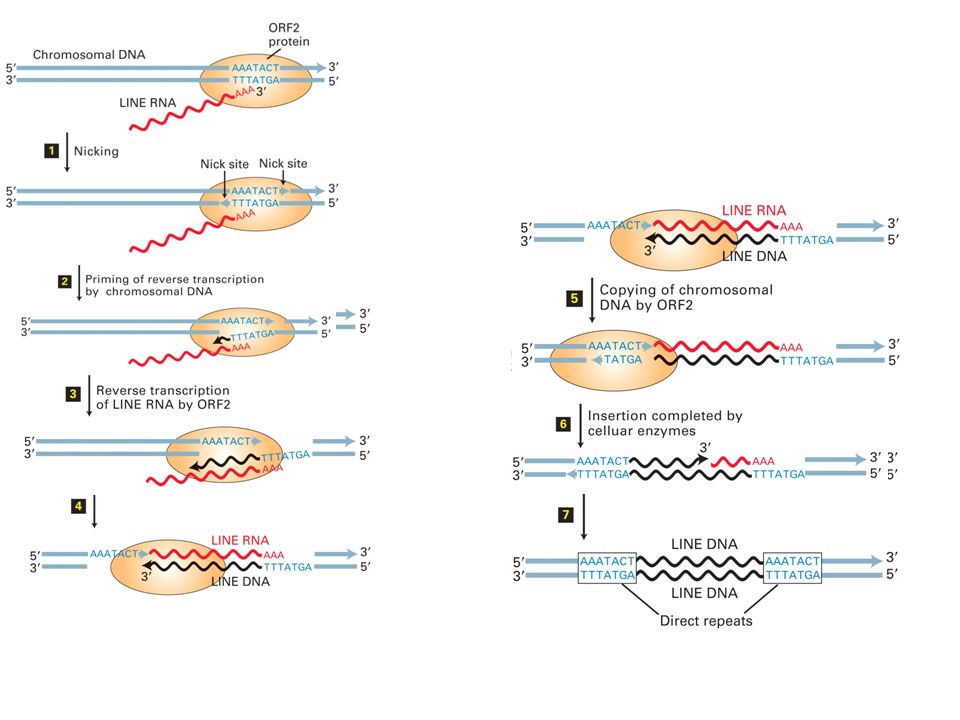
Usano un meccanismo chiamato trascrizione inversa innescata dal sito bersaglio Il RNA codifica per due proteine: ORF1 lega l’RNA, ORF2 ha attività di trascrittasi inversa e endonucleasi. Portano l’RNA sul bersaglio Dopo il taglio il 3’OH del DNA fa da primer e RNA da templato.

25
Esempio di DNA transposons:
Bacterial Insertion Sequences (IS elements) 1-2 kb. La parte centrale codifica una transposase
 1-2 kb. La parte centrale codifica una transposase.")
26
Modello di trasposizione di IS batteriche
Caratteristiche: Inverted repeats (50 b) Direct repeats (5-11 b)
 Direct repeats (5-11 b)")
27
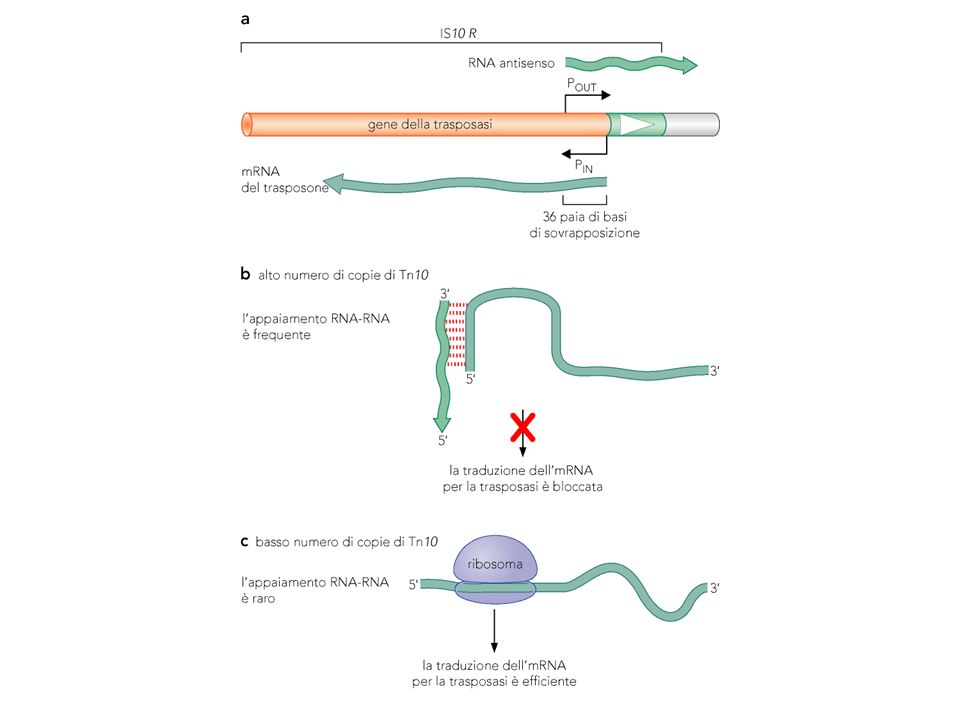
Tn10 batterico Appartiene alla famiglia IS4, con sistema taglia e incolla Di 9 kb, contiene la trasposasi e la resistenza alla tetraciclina Alle estremità ha IS10R (con la trasposasi) e IS10L. Usa RNA antisenso per regolare la espressione della trasposasi
 e IS10L. Usa RNA antisenso per regolare la espressione della trasposasi.")
30
Tc1/mariner Sono tra i più semplici trasposoni autonomi, Tc1 è di C. elegans, mariner è di Drosophila Sono di 1,5 – 2,5 kb e contengono le sequenze terminali ed il gene per una trasposasi. Non necessitano di proteine accessorie, usano sistema taglia e incolla La loro trasposizione non è ben regolata, e nel genoma ve ne sono molti “morti” Sleeping beauty è una versione ingegnerizzata ad alta attività. Usato in biotecnologia

31
LTR retrotransposons Alcuni contengono long terminal repeats (LTR, bp) Codificano le proteine tipiche dei retrovirus, eccetto quelle dell’envelope. Codificano reverse transcriptase e integrase Nell’uomo la maggior parte deriva dal retrovirus endogeno ERV

32
Retrotrasposoni senza LTR
Chiamati anche retrotransposoni non virali. Formano due classi: Long interspersed elements (LINE) (6 kb), promuovono la loro mobilità e forniscono le proteine per la mobilità di SINE Short interspersed elements (SINE) (300 bp) Hanno sequenze simili ai geni: un promotore e poliA. Spesso si presentano come pseudogeni processati, senza il 5’
 (6 kb), promuovono la loro mobilità e forniscono le proteine per la mobilità di SINE. Short interspersed elements (SINE) (300 bp) Hanno sequenze simili ai geni: un promotore e poliA. Spesso si presentano come pseudogeni processati, senza il 5’")
33
LINE Tre maggiori famiglie: L1, L2 ed L3
L1 è la più abbondante (21% DNA totale) Codifica: RNA-binding protein (ORF1) e rev transcripatase/DNA endonuclease (ORF2).
 Codifica: RNA-binding protein (ORF1) e rev transcripatase/DNA endonuclease (ORF2).")
35
SINE 13% DNA umano 100-400 bp. Non codificano proteine.
Hanno una sequenza ricca in A/T, come LINE. 1.6 milioni di siti, di cui 1.1 sono elementi Alu

36
Ricombinazione V(D)J Adiacenti ai geni della ricombinazione V(D)J vi sono le sequenze segnale per la ricombinazione: l’eptamero e il nonamero. Le ricombinasi sono RAG1 e RAG2.

37
Exon shuffling per ricombinazione tra interspersed repeats omologhi

38
Exon shuffling per transposizione