Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
Le funzioni dei geni STM e WUS sono interconnesse
a vie di signaling ormonali CITOCHININE GIBBERELLINE

2
Diversi studi indicano che esiste una correlazione positiva tra
l’espressione/funzione dei geni KNOX e le CITOCHININE (CK) CK promuovono la divisione cellulare e la morfogenesi Piante overesprimenti KNOX hanno elevati livelli di CK Piante con elevati livelli di CK overesprimono geni KNOX e una correlazione negativa con le GIBBERELLINE (GA) Le GA promuovono l’espansione cellulare e la crescita degli organi, incompatibile con il mantenimento dell’omeostasi meristematica STM reprime la sintesi di gibberelline nel SAM
 CK promuovono la divisione cellulare e la morfogenesi. Piante overesprimenti KNOX hanno elevati livelli di CK. Piante con elevati livelli di CK overesprimono geni KNOX. e una correlazione negativa con le GIBBERELLINE (GA) Le GA promuovono l’espansione cellulare e la crescita degli organi, incompatibile con il mantenimento dell’omeostasi meristematica. STM reprime la sintesi di gibberelline nel SAM.")
3
inibiscono la crescita del meristema
Wt spy S::AtCKX3 spy-1,35S::AtCKX3 GA CK inibiscono la crescita del meristema spy-1,35S::AtCKX3 spindly: attivazione costitutiva GA signaling mutanti spindly con ridotti livelli di CK (35S::AtCKX3) hanno bassa attività meristematica spy-1 Spy-1,35S::AtCKX3 crescita determinata del meristema della infiorescenza spy-1 wt spy-1;AtCKX3
 hanno bassa attività. meristematica. spy-1. Spy-1,35S::AtCKX3. crescita determinata del meristema della infiorescenza. spy-1. wt. spy-1;AtCKX3.")
4
GR= promotore del recettore dei glucocorticoidi DEX= dexametasone
Piante transgeniche di arabidopsis overesprimenti GR::STM (promotore inducibile) GR= promotore del recettore dei glucocorticoidi DEX= dexametasone RTPCR 10gg 1mM DEX STM nell’apice induce la sintesi di geni della biosintesi delle CK (IPT7) e la sintesi di geni di della via di signaling (ARR5)
 GR= promotore del recettore dei glucocorticoidi. DEX= dexametasone. RTPCR. 10gg 1mM DEX. STM nell’apice induce la sintesi di geni della biosintesi delle CK (IPT7) e la sintesi di geni di della via di signaling (ARR5)")
5
Piante transgeniche di arabidopsis overesprimenti STM (GR::STM) RTPCR
STM nell’apice induce l’espressione di geni (GA2 ossidasi) che inattivano le GA (da studi di espressione di GA2 ossidasi in mutanti wol: effetto non diretto ma mediato da CK )
 che inattivano le GA. (da studi di espressione di GA2 ossidasi in mutanti wol: effetto non diretto ma mediato da CK )")
6
In tabacco dimostrato che geni KNOX (STM) agiscono come repressori
trascrizionali del gene della GA20 ossidasi La GA20 ossidasi è un enzima necessario alla di GA attive

7
Biosintesi delle Gibberelline (dalla GA12- aldeide)
GA20 ox

8
Via di Biosintesi delle GA

9
STM nel SAM attiva la biosintesi delle CK e reprime sintesi della GA20ox e quindi la biosintesi delle GA e promuove l’attività meristematica. CK attivano l’espressione di GA2ox stimolando l’inattivazione delle GA. Il risultato è che le GA attive vengono confinate nelle foglie cotiledonari

10
GA2ox controls GA movement into the shoot meristem
Before the transition to flowering, OsGA2ox1 is expressed just below the shoot meristem, selectively excluding GA1 and GA4. GA4 GA5 It has been shown in Lolium that GA5 (which is resistant to deactivation by GA2ox) can move into the meristem despite GA2ox activity. OsGA2ox1 mRNA In the cartoon on the right the position of the buried meristem in the plant is indicated in yellow. Beside it in , the ring of GA2ox activity is shown in blue in the cartoon meristem. The image on the left shows a cross-section through a rice apex with the expression pattern of OsGA2ox1 shown in purple (by in situ hybridization). Sakamoto, T., Kobayashi, M., Itoh, H., Tagiri, A., Kayano, T., Tanaka, H., Iwahori, S., and Matsuoka, M. (2001). Expression of a gibberellin 2-oxidase gene around the shoot apex is related to phase transition in rice. Plant Physiol. 125:
 can move into the meristem despite GA2ox activity. OsGA2ox1 mRNA. In the cartoon on the right the position of the buried meristem in the plant is indicated in yellow. Beside it in , the ring of GA2ox activity is shown in blue in the cartoon meristem. The image on the left shows a cross-section through a rice apex with the expression pattern of OsGA2ox1 shown in purple (by in situ hybridization). Sakamoto, T., Kobayashi, M., Itoh, H., Tagiri, A., Kayano, T., Tanaka, H., Iwahori, S., and Matsuoka, M. (2001). Expression of a gibberellin 2-oxidase gene around the shoot apex is related to phase transition in rice. Plant Physiol. 125:")
11
FUNZIONE DEL GENE WUSCHEL

12
Piante di arabidopsis overesprimenti WUS con promotore inducibile da etanolo
Espressione ectopica di AG::GUS In risposta all’induzione di WUS (AGAMOUS= bersaglio di wuschel) Microarray STM WUS Alc::GUS Alc::CLV3 t ARR5 ARR6 ARR7 ARR15 RT-PCR RT PCR
 Microarray. STM WUS. Alc::GUS. Alc::CLV3. t. ARR5 ARR6 ARR7 ARR15. RT-PCR. RT PCR.")
13
WUSCHEL Reprime la trascrizione dei geni ARR5, ARR6, ARR7 e ARR15 che codificano per regolatori di risposta nel sistema a due componenti attivato dalle citokinine ARR5, ARR6, ARR7, e ARR15 agiscono come regolatori negativi della risposta alle citokinine

14
MECCANISMO DI AZIONE DELLE CK

15
NETWORK REGOLATIVI PER IL MANTENIMENTO DEL SAM

16
NETWORK REGOLATIVO DI WUS
STIMPY (STIP/WOX9): necessario per l’aumento delle dimensioni del meristema nel mutante clv3-2 ULTRAPETALA HD-ZIP HANABA TARANU AP2 Regolatori negativi di WUS SPLAYED (SYD) : regolatore diretto di WUS (fattore di rimodellamento della cromatina)
: necessario per l’aumento delle dimensioni del meristema. nel mutante clv3-2. ULTRAPETALA. HD-ZIP. HANABA TARANU. AP2. Regolatori negativi di WUS. SPLAYED (SYD) : regolatore diretto di WUS. (fattore di rimodellamento della cromatina)")
17
INTERAZIONE TRA FATTORI DI TRASCRIZIONE E ORMONI NELL’APICE
MERISTEMATICO

18
FORMAZIONE DEL MERISTEMA APICALE
DELLA RADICE

19
L'ipofisi, allo stadio “a cuore” (=emersione degli abbozzi dei cotiledoni) si divide in due cellule, dando una cellula superiore a forma di lente, e una cellula inferiore che va incontro subito ad una divisione longitudinale. La cellula lenticolare si dividerà ulteriormente e formerà la porzione interna del RAM, ovvero le 4 cellule del centro quiescente. Le due cellule inferiori si divideranno sia longitudinalmente sia trasversalmente, formando le iniziali della porzione apicale della cuffia radicale (o columella). L'ipofisi, nello stadio globulare tardivo si divide in due cellule, dando una cellula superiore a forma di lente, e una cellula inferiore che va incontro subito ad una divisione longitudinale. La cellula lenticolare si dividerà ulteriormente e formerà la porzione interna del RAM, ovvero le 4 cellule del centro quiescente. Le due cellule inferiori si divideranno sia longitudinalmente sia trasversalmente, formando le iniziali della porzione apicale della cuffia radicale (o columella).
.")
21
Apice radicale di arabidopsis

22
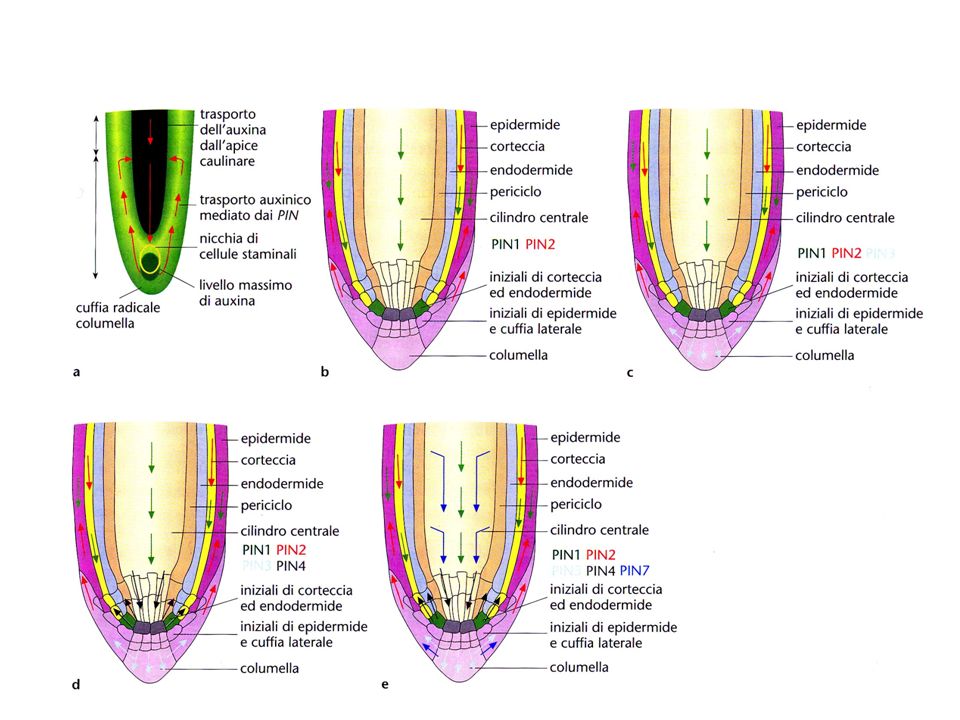
Il segnale di formazione del meristema apicale radicale è generato da
alti livelli di IAA nella regione basale del proembrione Elevati livelli di IAA sono causati dal trasporto polare di IAA mediato dalle proteine PIN

23
Espressione geni PIN Espressi con diversa localizzazione in tempi diversi dello sviluppo dell’embrione; La sequenza di espressione, regolata temporalmente e spazialmente, è responsabile della variazione della direzione del flusso di IAA durante l’embriogenesi.

25
Trasporto di IAA nella radice
Massima concentrazione di IAA nelle iniziali della columella

26
Proteine PIN PIN1 parte basale delle cellule del cilindro vascolare;
PIN2 estremità basale delle cellule corticali e all’estremità apicale delle cellule epidermiche e della cuffia laterale; PIN3 espressa, senza polarità apparente, in due file sovrapposte di cellule della columella e nel periciclo; PIN4 localizzazione non polarizzata nelle cellule del centro quiescente e nelle cellule che lo circondano, nonché all’estremità basale nelle cellule provascolari; PIN7 nelle membrane laterali e basali delle cellule provascolari, nella zona di distensione e nelle cellule della columella.

29
CENTRO QUIESCENTE (QC)
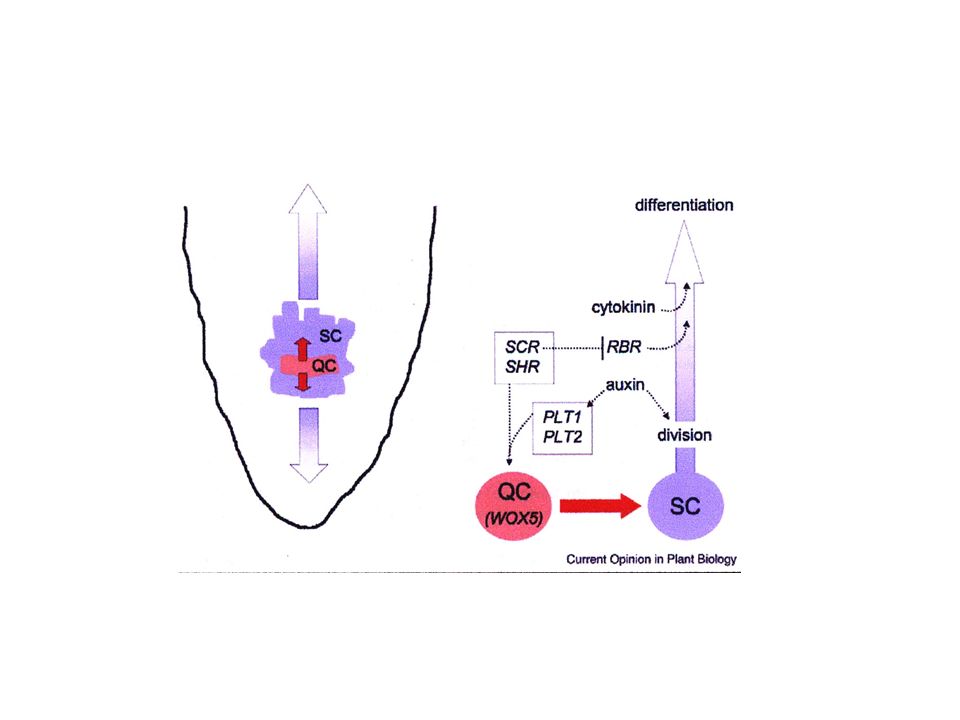
Nei meristemi del germoglio (SAM) e della radice (RAM) la popolazione di cellule staminali è mantenuta grazie all’attività di un centro organizzatore SAM: centro organizzatore specificato dal gene WUSCHEL RAM: centro quiescente (QC); l’identità di QC è specificata dai geni SHORT ROOT e SCARECROW è richiesta anche l’espressione dei geni PLETHORA1 e PLETHORA2 Codificano per fattori di trascrizione di tipo AP2 Sono espressi inizialmente nella regione basale dell’embrione, poi nel primordio della radice e infine nel centro quiescente
 e della radice (RAM) la popolazione. di cellule staminali è mantenuta grazie all’attività di un centro organizzatore. SAM: centro organizzatore specificato dal gene WUSCHEL. RAM: centro quiescente (QC); l’identità di QC è specificata dai geni. SHORT ROOT e SCARECROW. è richiesta anche l’espressione dei geni PLETHORA1 e PLETHORA2. Codificano per fattori di trascrizione di tipo AP2. Sono espressi inizialmente nella regione basale dell’embrione, poi nel primordio. della radice e infine nel centro quiescente.")
30
CENTRO QUIESCENTE Localizzazione determinata da massimo di concentrazione di auxina Espressione dei geni SHORT ROOT e SCARECROW Espressione dei geni PLETHORA1 e PLETHORA2

31
SHORT-ROOT (SHR) e SCARECROW (SCR)
Le proteine SHR sono prodotte nella stele e traslocano nelle cellule adiacenti Necessarie per la formazione ed il mantenimento delle cellule staminali del CQ. Attivazione espressione SRC

32
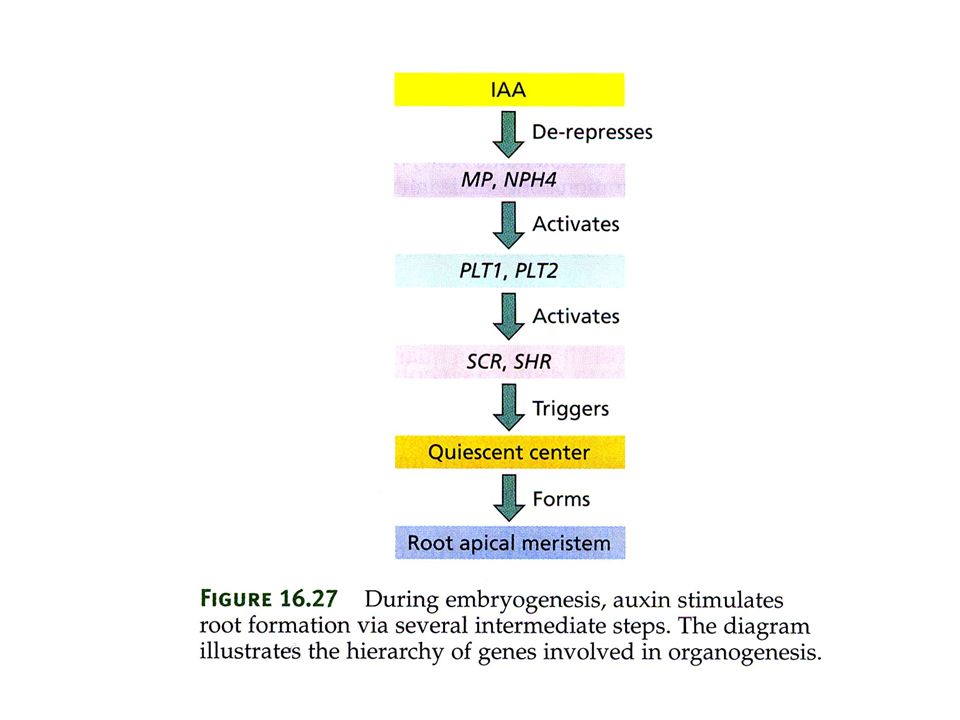
Geni PLETHORA (PLT) Codificano per fattori di trascrizione di tipo AP2; Richiesti per specificare l’identità delle cellule del QC e per il corretto pattern di divisione delle cellule circostanti; La loro espressione è controllata da MONOPTEROS; L’espressione dei geni PLT è modulata e modula l’espressione dei geni PIN. Le proteine PIN restringono l’espressione di PLT nella regione basale dell’embrione per consentire la formazione del primordio della radice (embrione di 8/16 cellule); In uno stato successivo dell’embriogenesi, PLT mantiene la trascrizione dei geni PIN e definisce la posizione della nicchia delle cellule staminali; Ad una determinata distanza dal punto di massima [IAA], PLT promuove l’espressione di PIN3 e PIN7, che rinforzano il flusso dell’IAA in direzione acropeta, regolando la divisione cellulare (DIV) nella zona meristematica e l’espansione cellulare nella zona di elongazione (EL).
; In uno stato successivo dell’embriogenesi, PLT mantiene la trascrizione dei geni PIN e definisce la posizione della nicchia delle cellule staminali; Ad una determinata distanza dal punto di massima [IAA], PLT promuove l’espressione di PIN3 e PIN7, che rinforzano il flusso dell’IAA in direzione acropeta, regolando la divisione cellulare (DIV) nella zona meristematica e l’espansione cellulare nella zona di elongazione (EL).")
33
PLETHORA Mutanti plt1 e plt2 hanno alterazioni nel pattern di divisioni cellulari nel QC e nella columella Columella WT: 4 file di cellule la prima di staminali le altre differenziate (accumulo granuli di amido) Columella plt1: ulteriori file di cellule dovute ad aumento di divisioni nello strato 1 QC WT plt1-4 plt2-2 plt1-4 plt2-2
 Columella plt1: ulteriori file di cellule dovute ad aumento di divisioni nello strato 1. QC. WT. plt1-4. plt2-2. plt1-4 plt2-2.")
34
Mutazioni dei geni PLT riducono il numero di cellule meristematiche
e la lunghezza della radice La columella contiene più cellule , la struttura a strati è alterata; si accumulano granuli di amido anche negli strati che dovrebbero essere occupati da cellule staminali La crescita della radice è ridotta Il numero delle cellule meristematiche è molto ridotto L’espressione del gene reporter cyclin-GUS (marcatore fase G2/M) è limitata Sviluppo postembrionale Zona meristematica plt1-4 plt2-2
 è limitata. Sviluppo postembrionale. Zona meristematica. plt1-4 plt2-2.")
35
I geni PLT sono espressi nella regione basale dell’embrione nello
stadio globulare a 8 cellule sono richiesti per specificare l’identità delle cellule del QC cellula progenitrice del QC Ibridazione in situ PLT1

36
Espressione di marcatori del centro quiescente nello stadio a cuore
(QC25) (QC46) WT plt1-4 plt2-2 WT plt1-4 plt2-2 La funzione PLT è richiesta nello stadio iniziale della determinazione dell’identità del QC e per il corretto pattern di divisione delle cellule circostanti
 (QC46) WT. plt1-4 plt2-2. WT. plt1-4 plt2-2. La funzione PLT è richiesta nello stadio iniziale della determinazione dell’identità del QC. e per il corretto pattern di divisione delle cellule circostanti.")
37
PLT1 e PLT2 agiscono in parallelo con i geni SHORT ROOT/SCARECROW
nel pattern di specificazione del centro quiescente SCR e SHR necessari per l’espressione dei marcatori del CQ (QC 25; QC46) I domini di espressione di PLT1 PLT2 e SHR/SCR si stabiliscono in maniera Indipendente wt plt1-4 plt2-2 wt plt1-4 plt2-2 Espressione di SHR::GFP Espressione di SCR::YFP
 I domini di espressione di PLT1 PLT2 e SHR/SCR si stabiliscono in maniera. Indipendente. wt. plt1-4 plt2-2. wt. plt1-4 plt2-2. Espressione di SHR::GFP. Espressione di SCR::YFP.")
38
Ibridazione in situ PLT1 wt scr-1 shr-1 3dpg

39
e dipende dai fattori di risposta all’auxina (ARF)
L’espressione dei geni PLT risponde a cambiamenti di distribuzione dell’auxina e dipende dai fattori di risposta all’auxina (ARF) Embrione nello stato di transizione doppio mutante mp-G12 nph4-1 Ibridazione in situ assenza di segnale per PLT1 Necessario doppio mutante per ridondanza proteine ARF
 Embrione nello stato di transizione. doppio mutante mp-G12 nph4-1. Ibridazione in situ. assenza di segnale per PLT1. Necessario doppio mutante per ridondanza proteine ARF.")
40
L’espressione di PLT è controllata dal gradiente di IAA determinato dalla
localizzazione di PIN e da MP; Le proteine SHR sono prodotte nella stele e traslocano nelle cellule adiacenti, dove attivano SRC; Il centro quiescente è specificato nella zona d’intersezione dei domini d’espressione dei geni PLT, SHR e SCR; I pattern d’espressione di SHR, SCR e PLT definiscono le differenti regioni; In presenza dei geni PLT → SHR e SCR sono coinvolti nella specificazione delle cellule staminali; in assenza di PLT → SHR regola il differenziamento delle cellule derivate dalle staminali.

41
MODELLO Le cellule che esprimono PLT SCR e SHR diventano QC
QC segnala alle cellule circostanti per mantenere l’identità meristematica

42
Esperimenti di ablazione con laser del centro quiescente
alterazione dei flussi auxinici rimodulazione della espressione dei geni PLT (SHR, SCR, PIN) rispecificazione del centro quiescente qc ablati DR5::GFP SHR SCR
 rispecificazione del centro quiescente. qc. ablati. DR5::GFP. SHR. SCR.")
43
ablati qc PIN1 PIN2

44
Ablazione: shift gradiente auxinico; espressione PLT; inibizione espressione PIN;
Loc nucleare di SHR; promozione espressione SCR

46
qc

47
Network regolativo nel meristema della radice
WOX5 è un omologo di WUSCHEL: contribuisce a mantenere le cellule del CQ in srtato indifferenziato

48
CK nel meristema radicale
controllano la velocità di differenziamento delle cellule meristematiche della radice e quindi le dimensioni del meristema

49
CK esogene riducono il numero di
Dimensioni del Meristema: N° di 1 fila di cellule del cortex dal QC alla TZ CK esogene riducono il numero di cellule meristematiche senza influenzare Le dimensioni del QC e della Columella quindi senza influire sul loro potenziale meristematico

50
Mutanti biosintetici per le CK o mutanti nella via di signaling hanno meristemi
più grandi ahk3

51
Recettori per le CK (sistema a due componenti Istidina kinasi)
AHK4/CREI1/WOL AHK3 AHK2 AHK3 è l’unico espresso in tutti i tessuti della zona di transizione AHK3 ARR1 ARR12 ARR1 e ARR12 sono espressi solo nella TZ

52
UN SISTEMA A DUE COMPONENTI DI SIGNALING PER LE CK BASATO SU
MODELLO UN SISTEMA A DUE COMPONENTI DI SIGNALING PER LE CK BASATO SU AHK3/ARR1, AHK3/ARR12 MEDIA IL CONTROLLO DEL NUMERO DELLE CELLULE MERITEMATICHE NELLA ZONA DI TRANSIZIONE LE CK AGISCONO ANTAGONIZZANDO UN SEGNALE DI DIVISIONE CELLULARE (AUXINA) Applicazioni di auxina esogena alla radice incrementano le dimensioni del meristema La riduzione di livelli di CK in tripli mutanti pin non ha effetto sullle dimensioni del meristema
 Applicazioni di auxina esogena alla radice incrementano le dimensioni del meristema. La riduzione di livelli di CK in tripli mutanti pin non ha effetto sullle dimensioni del meristema.")
53
CK attivano SHY2 che media la repressione dell’espressione delle proteine PIN
IAA media la degradazione tramite proteosoma di SHY2 IAA è il segnale per la divisione cellulare meristematica nella radice

54
SHY2 è una proteina AUXIAA
Controlla la biosintesi delle CK Attraverso la IPT5 Gli effetti antagonistici tra IAA e CK Sono spiegati in base al conrollo negativo da parte di CK sulla distribuzione PIN-dipendente di IAA

56
RBR: PROTEINA DEL RETINOBLASTOMA (antagonizza nei mammiferi la
funzione meristematica) Mutanti di arabidopsis rbr loss of function mostrano divisioni cellulari in più e ritardo del differenziamento della radice L’overespressione di RBR causa differenziamento precoce RBR proteina regolativa per il differenziamento nella radice Non sono alterati i pattern di espressione di PLT, SCR, SHR RBR agisce a valle di SCR/SHR; PLT mutanti scr hanno livelli più alti di RBR
 Mutanti di arabidopsis rbr loss of function mostrano divisioni cellulari in più e ritardo. del differenziamento della radice. L’overespressione di RBR causa differenziamento precoce. RBR proteina regolativa per il differenziamento nella radice. Non sono alterati i pattern di espressione di PLT, SCR, SHR. RBR agisce a valle di SCR/SHR; PLT. mutanti scr hanno livelli più alti di RBR.")
57
SAM/RAM Le CK promuovono il mantenimento del meristema del germoglio
ma hanno la funzione opposta nel meristema radicale dove stimolano il differenziamento nella zona di transizione L’auxina è necessaria per la formazione del meristema radicale ma nel meristema del germoglio promuove l’organogenesi Sia nel germoglio che nella radice il mantenimento del meristema dipende dal bilancio tra CK e auxina ma le risposte sono diverse nei due meristemi Quali funzioni cellulari sono controllate dai due ormoni nei meristemi del germoglio e della radice?

58
DIFFERENZIAMENTO DI CELLULE EPIDERMICHE NELLA RADICE
Le cellule epidermiche confinanti con lo spazio tra due corticali diventano TRICOBLASTI

59
SE UN TRICOBLASTO VIENE RIMOSSO (ablazione laser)
L’ A TRICOBLASTO VICINO PRENDE IL SUO POSTO ( e viceversa) IL DIFFERENZIAMENTO E’ DIRETTO DALLA INFORMAZIONE POSIZIONALE
 IL DIFFERENZIAMENTO E’ DIRETTO DALLA INFORMAZIONE POSIZIONALE.")
60
Mediante l’analisi di mutanti identificati geni regolatori
per il differenziamento DEI TRICOBLASTI WEREWOLF (WER) (myb) regolatore negativo dell’identità tricoblastica CAPRICE (CPC) (myb) CPC regolatore negativo dell’identità atricoblastica
 (myb) regolatore negativo dell’identità tricoblastica. CAPRICE (CPC) (myb) CPC regolatore negativo dell’identità atricoblastica.")
61
TTG1: TRANSPARENT TESTA GLABA1
GL3 : GLABRA3 (2) (1) EGL3: ENHANCER OF GLABRA3 SCM: SCRAMBLED CAPRICE: CPC
 (1) EGL3: ENHANCER OF GLABRA3. SCM: SCRAMBLED. CAPRICE: CPC.")
62
Modello regolativo per il differenziamento dei tricoblasti nella radice
LRR-RLK CPC è espresso nelle cellule N nonostante il suo effetto inibitorio sul differenziamento azione non-cell-autonomous del gene CPC

63
Gl2 (+) WER ATRICOBLASTO Gl2 (-) TRICOBLASTO CPC
 WER ATRICOBLASTO Gl2 (-) TRICOBLASTO CPC")
Presentazioni simili