Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
LO SVILUPPO DEL FIORE

2
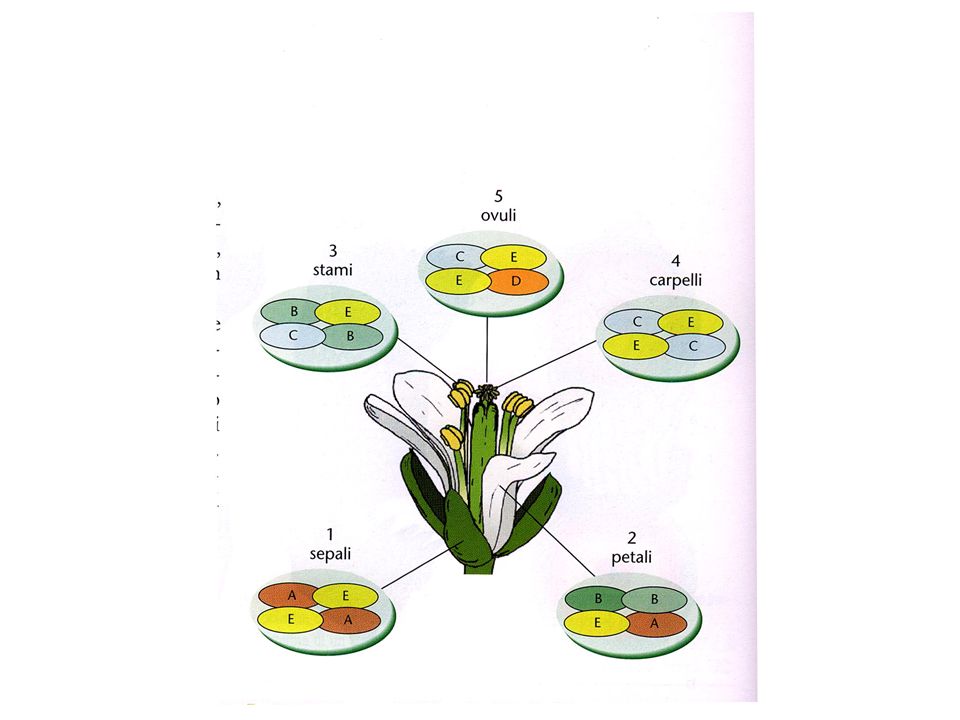
Struttura schematica di un fiore
(ermafrodita)
")
3
Fiore staminifero Fiore pistillifero

4
FIORE: complesso apparato di strutture funzionalmente specializzate e
radicalmente diverse dall’organismo vegetativo sia nella forma che nei tipi cellulari La transizione verso la fioritura implica cambiamenti radicali nel destino delle cellule dei meristemi apicali Alterazione del destino delle cellule meristematiche: cambiamento di fase L’insieme degli eventi che portano l’apice del germoglio a produrre fiori si indica come INDUZIONE FIORALE

5
INDUZIONE FIORALE

6
AUMENTO DEL TASSO DI DIVISIONE DELLE CELLULE CENTRALI
MERISTEMI FIORALI: distinguibili dai vegetativi perché di dimensioni maggiori Transizione allo stadio riproduttivo segnata da un aumento delle divisioni cellulari nella zona centrale del SAM. AUMENTO DEL TASSO DI DIVISIONE DELLE CELLULE CENTRALI

7
Sezione longitudinale della regione apicale di un germoglio vegetativo e
di un germoglio riproduttivo di Arabidopsis mf mi pf gf mg

8
Il meristema fiorale determina la formazione di quattro organi
organizzati in cerchi concentrici verticilli La formazione degli organi più interni (carpelli) esaurisce le cellule meristematiche nella zona apicale e rimangono solo i primordi degli organi fiorali In Arabidopsis dall’esterno: Quattro sepali (verdi) Quattro petali (bianchi) Sei stami (quattro più lunghi) Un pistillo (composto da due carpelli fusi a formare l’ovario; lo stilo e lo stigma)
 esaurisce le cellule meristematiche nella. zona apicale e rimangono solo i primordi degli organi fiorali. In Arabidopsis dall’esterno: Quattro sepali (verdi) Quattro petali (bianchi) Sei stami (quattro più lunghi) Un pistillo (composto da due carpelli fusi a formare l’ovario; lo stilo e lo stigma)")
9
Organi fiorali formati in sequenza dal meristema fiorale di Arabidopsis

10
Il fiore di arabidopsis (attinomorfo: simmetria raggiata)
")
11
Pistillo di Arabidopsis

12
Simmetria bilaterale Simmetria raggiata

13
Il fiore di Anthirrinum (zigomorfo: simmetria bilaterale)
Doppio mutante cycloidea/dichotoma

14
Fiore del riso: perianzio ridotto

15
Studi condotti principalmente su Arabidopsis
e Anthirrinum (bocca di leone) Identificazione di una rete di geni che controlla la morfogenesi del fiore
 Identificazione di una rete di geni che controlla la. morfogenesi del fiore.")
16
Identificate tre classi di geni che controllano lo sviluppo del fiore
Geni che determinano l’identità degli organi fiorali (fattori di trascrizione che controllano l’espressione di geni di differenziamento) Geni Catastali (regolatori spaziali della espressione dei geni di identità degli organi) Geni di identità meristematica (fattori di trascrizione necessari per l’induzione dei geni di identità degli organi fiorali)
 Geni Catastali. (regolatori spaziali della espressione dei geni di identità degli organi) Geni di identità meristematica. (fattori di trascrizione necessari per l’induzione dei geni di identità. degli organi fiorali)")
17
Lo studio di MUTANTI FIORALI OMEOTICI
ha portato alla scoperta dei geni che controllano l’identità degli organi fiorali

18
I geni che determinano l’identità degli organi fiorali sono definiti
OMEOTICI funzionalmente,in quanto mutazioni di questi geni comportano la formazione di un organo al posto di un altro (es: carpelli al posto di sepali). Tuttavia sono strutturalmente diversi dai geni omeotici in quanto non contengono il motivo HOMEOBOX ma appartengono alla classe di fattori MADS box MADS box è un motivo DNA binding conservato di 56 aa Inoltre è presente un motivo K box di interazione Proteina-proteina
. Tuttavia sono strutturalmente diversi dai geni omeotici in quanto non. contengono il motivo HOMEOBOX ma appartengono alla classe di. fattori MADS box. MADS box è un motivo DNA binding conservato. di 56 aa. Inoltre è presente un motivo K box di interazione. Proteina-proteina.")
19
MADS: MCM1(lievito), AGAMOUS (arabidopsis), DEFICIENS (anthirrinum), SRF (uomo)
In Arabidopsis oltre 100 geni MADS BOX Animali o funghi ne possiedono un numero molto minore (Drosophila 2) Nelle piante molti coinvolti nello sviluppo del fiore APETALA1, APETALA3, PISTILLATA, AGAMOUS, SEPALLATA1, SEPALLATA2, SEPALLATA3 5 classi:Ma, Mb, Mg, Md e MIKC
 Nelle piante molti coinvolti nello sviluppo del fiore. APETALA1, APETALA3, PISTILLATA, AGAMOUS, SEPALLATA1, SEPALLATA2, SEPALLATA3. 5 classi:Ma, Mb, Mg, Md e MIKC.")
20
GENI MADS box in specie diverse
Anthirrinum petunia arabidopsis pomodoro

21
MODELLO ABC Negli anni 90 E. Meyerowitz, E. Coen et al.
proposero il modello ABC per l’identità degli organi fiorali Basato sullo studio dei mutanti fiorali omeotici di arabidopsis e anthirrinum modello semplice e applicabile a molte angiosperme ancora oggi valido con integrazioni

22
wt apetala2-2 pistillata2 agamous1

23
Il modello ABC postula che l’identità d’organo nel fiore è controllata
dalla combinazione di tre funzioni geniche A B C nei verticilli fiorali A = SEPALI A+B = PETALI B+C = STAMI C = CARPELLI A e C si reprimono reciprocamente

24
Classe A: APETALA1 (AP1), APETALA 2 (AP2)
In Arabidopsis Classe A: APETALA1 (AP1), APETALA 2 (AP2) Classe B: APETALA3 (AP3), PISTILLATA (PI) Classe C: AGAMOUS (AG) In Anthirrinum Classe A: SQUAMOSA (SQUA) Classe B: DEFICIENS (DEF), GLOBOSA (GLO) Classe C: PLENA
, APETALA 2 (AP2) Classe B: APETALA3 (AP3), PISTILLATA (PI) Classe C: AGAMOUS (AG) In Anthirrinum. Classe A: SQUAMOSA (SQUA) Classe B: DEFICIENS (DEF), GLOBOSA (GLO) Classe C: PLENA.")
25
A (AP1, AP2) specifica SEPALI A e B (AP3, PI) specificano PETALI
A e C (AP1, AG) specificano STAMI C (AG) specifica CARPELLI A e C si reprimono reciprocamente C (AGAMOUS) controlla anche lo stato di crescita determinata del meristema. L’assenza di C fa si che il meristema continui a crescere e nel quarto verticillo si formi un nuovo fiore
 specificano STAMI. C (AG) specifica CARPELLI. A e C si reprimono reciprocamente. C (AGAMOUS) controlla anche lo stato di crescita determinata del. meristema. L’assenza di C fa si che il meristema continui a crescere e. nel quarto verticillo si formi un nuovo fiore.")
26
Fenotipi dei mutanti omeotici fiorali secondo il modello ABC
wt agamous1

27
apetala2 pistillata2

28
wt apetala2 pistillata2 agamous1

29
INTEGRAZIONE DEL MODELLO ABC
Il mancato sviluppo di organi nelle posizioni corrette nei mutanti ABC dimostra che i geni ABC sono NECESSARI per specificare l’identità degli organi fiorali Sono anche sufficienti? Come test di sufficienza STUDI DI ESPRESSIONE ECTOPICA DEI GENI A BC (promotore 35S)
")
30
Nel fiore: verticilli esterni = Petali verticilli interni = Stami
Esempio più significativo overespressione geni di classe B (AP3; PI) Nel fiore: verticilli esterni = Petali verticilli interni = Stami Sufficienti In foglie a rosetta = mancata conversione in petali In foglie caulinari = debole conversione in petali Insufficienti Altri componenti oltre ad AP3/PI al di fuori del fiore sono necessari per specificare la funzione B di identità di organo
 Nel fiore: verticilli esterni = Petali. verticilli interni = Stami. Sufficienti. In foglie a rosetta = mancata conversione in petali. In foglie caulinari = debole conversione in petali. Insufficienti. Altri componenti oltre ad AP3/PI al di fuori del fiore sono. necessari per specificare la funzione B di identità di organo.")
31
geni SEP sono NECESSARI per specificare l’identità degli organi fiorali appartengono alla famiglia dei geni MADS La maggior parte dei geni ABC sono membri della famiglia MADS box (AP3; PI;AP1; AG) Inizialmente geni MADS identificati sulla base dell’ omologia con AG Designati come AGAMOUS-LIKE (AGL) [MADS = Minichromosome maintenance 1, Deficiens, Agamous, Serum response]
 Inizialmente geni MADS identificati sulla base dell’ omologia con AG. Designati come AGAMOUS-LIKE (AGL) [MADS = Minichromosome maintenance 1, Deficiens, Agamous, Serum response]")
32
SEP1 (AGL2); SEP2 (AGL4); SEP3 (AGL9)
Sono simili nella sequenza ed esibiscono lo stesso pattern di espressione temporale durante le fasi iniziali e intermedia dello sviluppo del fiore SEP 1 e SEP2 espressi in tutti e quattro i verticilli SEP3 espresso soltanto nei verticilli 2, 3 ,4

33
Mutanti sep singoli o doppi non mostrano fenotipo
Tripli mutanti sep1 sep2 sep3 esibiscono un fenotipo simile a doppi mutanti BC Come ad esempio pi ag oppure ap3 ag Nei tripli mutanti il fiore è formato interamente da sepali e i fiori sono indeterminati

34
Tutti e tre i geni SEP sono necessari per lo sviluppo corretto
degli organi fiorali (in assenza dei geni SEP i tre verticilli interni si sviluppano tutti come sepali) I geni SEP sono ridondanti (necessari i tripli mutanti per la manifestazione del fenotipo)
 I geni SEP sono ridondanti (necessari i tripli mutanti per la. manifestazione del fenotipo)")
35
MODELLO ABC integrato con i geni SEP

36
Come test di “SUFFICIENZA” i geni SEP sono stati espressi
ectopicamente insieme ai geni ABC B B,A B,C

37
L’espressione ectopica dei geni SEP insieme ai geni
ABC converte foglie caulinari o foglie a rosetta in petali o stami petalo stami petaloide AP3,PI, SEP3 AP3,PI, SEP3, AP1 AP3,PI, SEP3, AG wt

38
l’espressione ectopica di AP1 può sostituire la funzione dei geni SEP
Forse SEP necessari quando l’espressione di AP1 è a livelli normali, non 35S::AP1 oppure AP1 e SEP evolutivamente vicini e distinti da altri gruppi di geni MADS come Quelli cui appartengono AG, AP3, PI.

39
Gli esperimenti di espressione ectopica suggeriscono
che nei tessuti vegetativi sia l’assenza di un fattore positivo (SEP3 , AP1) piuttosto che la presenza di un repressore a impedire la trasformazione completa delle foglie in petali
 piuttosto che la presenza di un repressore. a impedire la trasformazione completa delle foglie in petali.")
40
SEP4 sep1 sep2 sep3 : 1 verticillo sepali
altri verticilli sepali omeotici (indeterminato) sep1 sep2 sep3 sep4: tutti i verticilli foglie omeotiche I geni sepallata sono necessari per la conversione delle foglie in organi fiorali
 sep1 sep2 sep3 sep4: tutti i verticilli foglie omeotiche. I geni sepallata sono necessari per la conversione delle foglie in organi fiorali.")
41
Quadruplo mutante ap1, ap2, ap3/pi, ag

42
INTERAZIONI MOLECOLARI TRA I GENI SEP e ABC
Entrambi appartenenti alla classe MADS box e necessari per il corretto sviluppo degli organi fiorali Relazioni regolative a livello trascrizionale? (Ad esempio la trascrizione dei geni ABC può dipendere dai geni SEP o viceversa) SEP RNA si accumula in singoli mutanti ABC Espressione di ABC avviene nei tripli mutanti sep1 sep2 sep3
 SEP RNA si accumula in singoli mutanti ABC. Espressione di ABC avviene nei tripli mutanti sep1 sep2 sep3.")
43
Proteine MADS si legano in vitro al DNA come omo o etero dimeri
INTERAZIONE DIRETTA ABC /SEP ETERODIMERIZZAZIONE Proteine MADS si legano in vitro al DNA come omo o etero dimeri Si legano ad un motivo consenso detto CArG box 5’-CC[AT]6GG-3’ ma SEP (anche AP1) non forma eterodimeri con AP3 o PI eterodimerizzazione non spiega la formazione di petali e stami (SEP +B)
 non forma eterodimeri con AP3 o PI. eterodimerizzazione non spiega la formazione di petali e stami (SEP +B)")
44
SPECIFICITA’ FUNZIONALE DELLE PROTEINE MADS
La specificità funzionale delle proteine ABC non sembra dipendere da una diversa specificità o affinità di legame al DNA Tutte le proteine ABC hanno caratteristiche di binding in vitro al DNA in larga parte sovrapponibili Come si spiega la specificità funzionale?

45
FORMAZIONE DI COMPLESSI DI ORDINE SUPERIORE
quartetto petali 2 eterodimeri che interagiscono tra loro (AP1)
")
46
IL Modello ABC (DE) e il modello del QUARTETTO
E= SEP D= STK, SHT

48
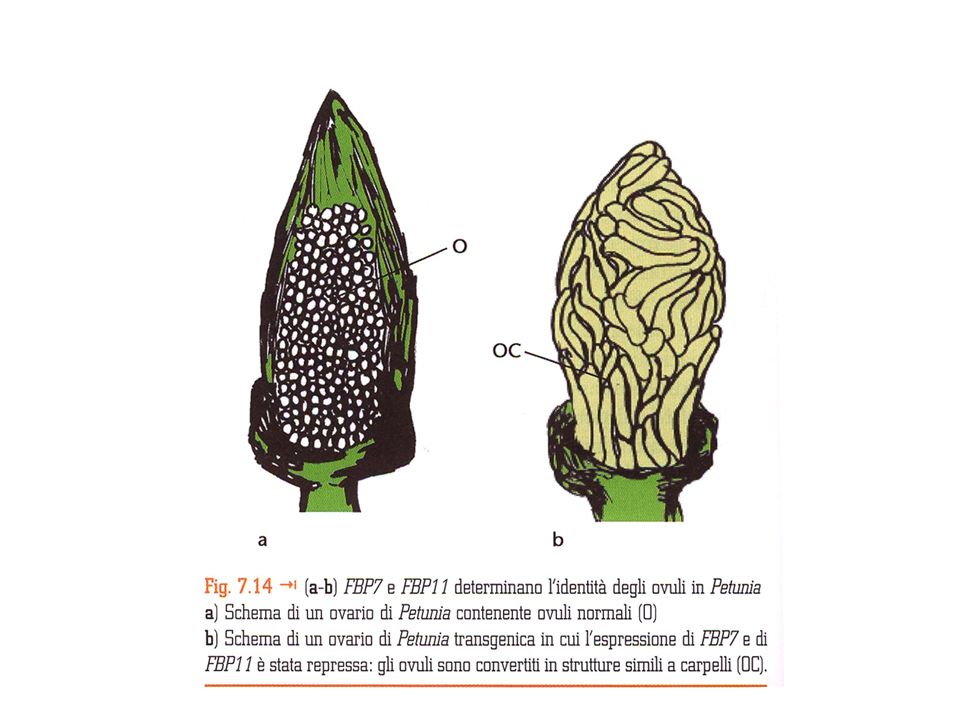
Da studi in petunia identificati i geni:
Geni di classe D Da studi in petunia identificati i geni: FLORAL BINDING PROTEIN7 (FBP7) FLORAL BINDING PROTEIN11 (FBP11) Che determinano l’identità dell’ovulo Il silenziamento di questi geni determina la conversione degli ovuli in strutture carpelloidi In Arabidopsis: SHP1, SHP2, , SKT
 FLORAL BINDING PROTEIN11 (FBP11) Che determinano l’identità dell’ovulo. Il silenziamento di questi geni determina la conversione. degli ovuli in strutture carpelloidi. In Arabidopsis: SHP1, SHP2, , SKT.")
50
PER UN CORRETTO SVILUPPO DEL FIORE NECESSARIA UNA
REGOLAZIONE SPAZIALE DELL’ESPRESSIONE DEI GENI OMEOTICI REGOLAZIONE TRASCRIZIONALE

51
UFO (UNUSUAL FLOWER ORGANS)
GENI CATASTALI agiscono come regolatori dell’espressione spaziale dei geni omeotici limitandone l’espressione nelle regioni appropriate Regolazione trascrizionale LFY (LEAFY) AP2 (APETALA2) SUP (SUPERMAN) UFO (UNUSUAL FLOWER ORGANS)
 AP2 (APETALA2) SUP (SUPERMAN) UFO (UNUSUAL FLOWER ORGANS)")
52
Regolazione dell’espressione di AG

53
Complesso LEUNIG/SEUSS (LUG/SEU)
Funziona da repressore dell’espressione di AG nel primo e secondo verticillo I singoli mutanti lug e seu mostrano sepali parzialmente convertiti in strutture petaloidi o carpelloidi nel primo verticillo e petali di dimensioni ridotte nel secondo; doppi mutanti hanno fenotipo più severo Espressione ectopica di AG e dei geni di classe B

54
AP2 Funziona come repressore di AG nel primo e secondo verticillo.
L’mRNA di AP2 è espresso in tutti e quattro i verticilli ma viene degradato mediante micro-RNA nel terzo e nel quarto; la proteina AP2, in grado di reprimere l’espressione di AG è dunque presente solo nel 1 e 2 verticillo

55
LFY Modulatore positivo dell’espressione di AG e dei geni di classe B (AP3; PI) Nei doppi mutanti lfy/ap1 il trascritto di AP3 è assente LFY+AP1 necessari per l’espressione di AP3

56
UFO Modulatore positivo di AP3 Mutanti ufo hanno un numero ridotto di petali e stami L’espressione ectopica di UFO provoca la conversione omeotica dei sepali in petali e dei carpelli in stami (come l’espressione ectopica di AP3 o PI) UFO è una proteina F box Attiva l’espressione di AP3 determinando la degradazione di un repressore
 UFO è una proteina F box. Attiva l’espressione di AP3 determinando la degradazione di un repressore.")
57
SUP regola negativamente l’espressione di
Mutazioni sup determinano la formazione di stami extra al posto dei carpelli In questi mutanti l’espressione di AP3 (B) normalmente ristretta al secondo e terzo verticillo si espande nel quarto (stami al posto di carpelli) SUP regola negativamente l’espressione di APETALA3 nel quarto verticillo
 normalmente ristretta. al secondo e terzo verticillo si espande nel quarto (stami al posto di carpelli) SUP regola negativamente l’espressione di. APETALA3 nel quarto verticillo.")
58
Regolazione dell’espressione di AP3

Presentazioni simili



















 Drosophila Small eye topo>")