Scaricare la presentazione
3
La S sta per unità Svedberg, l’unità che esprime il coefficiente di sedimentazione, ovvero la misura della velocità di sedimentazione di una particella sottoposta all’azione di un campo centrifugo. MAGGIORE E’ LA VELOCITA’ RAGGIUNTA DALLA PARTICELLA DURANTE LA CENTRIFUGAZIONE, MAGGIORE è IL SUO VALORE SVEDBERG. Il coefficiente di sedimentazione è una funzione del peso molecolare, della forma e del volume della particella. Particelle più pesanti e più compatte presentano un valore svedberg più elevato cioè sedimentano più in fretta.

4
Più sono "grandi" le particelle più sarà grande la velocità di sedimentazione e quindi avranno un più alto valore in unità svedberg. La velocità di sedimentazione non dipende solamente dalla grandezza (intesa come massa e volume totale) delle particelle (macromolecole biochimiche in genere). Quando due particelle si legano insieme vi è inevitabilmente una perdita di superficie totale da considerare. Il coefficiente di sedimentazione non è ADDITIVO, quindi, associando insieme n particelle la nuova particella avrà un valore in unità svedberg diverso dalla loro somma delle singole. In date condizioni fisiche (temperatura, pressione, viscosità del mezzo) il coefficiente di sedimentazione è influenzato soprattutto dalla forma del corpo che sedimenta -
 il coefficiente di sedimentazione è influenzato soprattutto dalla forma del corpo che sedimenta -")
5
ULTRACENTRIFUGA ANALITICA
determinazione della massa molecolare relativa analisi delle variazioni conformazionali studi sulla purezza del campione Velocità massima= g (campo centrifugo relativo) E’ costituita da: motore, rotore contenuto in una camera refrigerata in cui sia fatto il vuoto, sistema ottico di rivelamento del campione che sedimenta durante la centrifugazione
 E’ costituita da: motore, rotore contenuto in una camera refrigerata in cui sia fatto il vuoto, sistema ottico di rivelamento del campione che sedimenta durante la centrifugazione.")
9
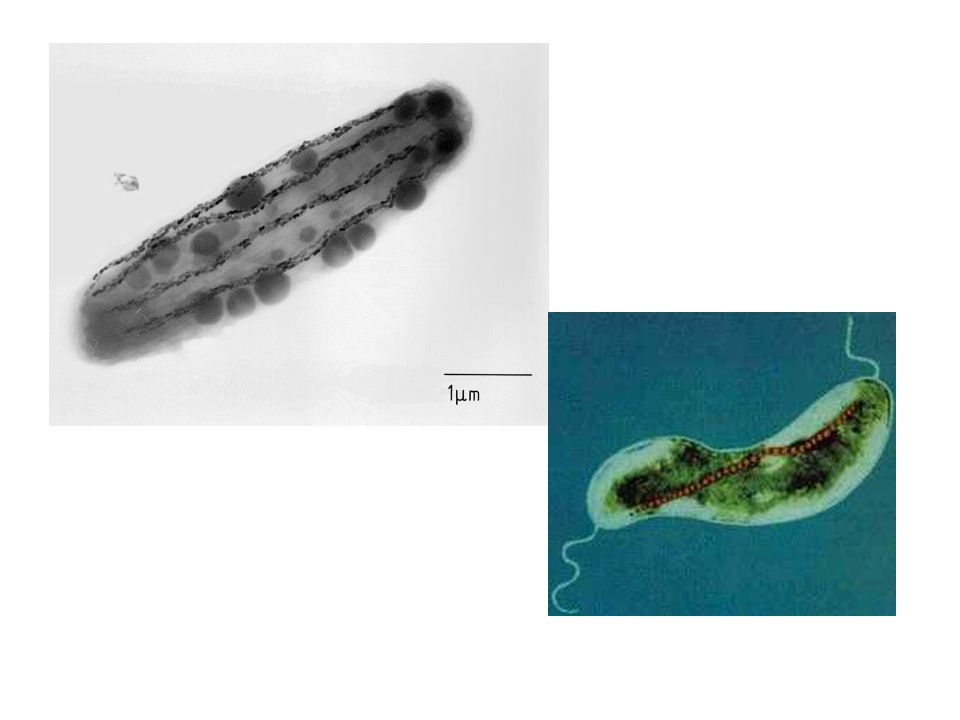
I Corpi d’inclusione (o inclusioni granulari) presenti nell’area citoplasmatica dei batteri
La matrice citoplasmatica dei batteri contiene un’ampia varietà di corpi di inclusione, ovvero granuli di sostanze organiche o inorganiche che spesso appaiono visibili al MO. In genere questi corpuscoli svolgono funzioni di deposito o riducono la pressione osmotica all’interno della cellula legando molecole. Alcuni corpi d’inclusione non sono delimitati da membrana e sono liberi nel citoplasma, altri sono circondati da una membrana monostratificata che può essere formata da proteine o lipidi

10
I corpi d’inclusione organici
Essi contengono soprattutto glicogeno (un polisaccaride formato da unità di glucosio) Amido Lipidi Nei cianobatteri e in alcune specie batteriche acquatiche sono interessanti i vacuoli gassosi. Le pareti di queste strutture non contengono lipidi, ma proteine.Esse sono impermeabili all’acqua e totalmente permeabili ai gas atmosferici.
 Amido. Lipidi. Nei cianobatteri e in alcune specie batteriche acquatiche sono interessanti i vacuoli gassosi. Le pareti di queste strutture non contengono lipidi, ma proteine.Esse sono impermeabili all’acqua e totalmente permeabili ai gas atmosferici.")
11
Le vescicole gassose sono state trovate nei cianobatteri,
che sono microrganismi fotosintetici e vivono in sistemi acquatici. Nei laghi e negli oceani, i cianobatteri possono controllare la loro posizione nella colonna d'acqua per ottenere la migliore quantità di luce e di nutrienti.

12
Magnetosomi • Alcuni batteri, quando messi in un campo magnetico, si orientano verso uno dei poli grazie alla presenza di magnetosomi (cristalli di magnetite (Fe3O4) disposti in forma di collana nel loro citoplasma). I magnetosomi orientano la cellula e ne determinano la direzione di avanzamento, per collocarsi in regioni favorevoli alla crescita. In genere i microrganismi che li possiedono sono acquatici e microaerofili o anaerobi
 disposti in forma di collana nel loro citoplasma). I magnetosomi orientano la cellula e ne determinano la direzione di avanzamento, per collocarsi in regioni favorevoli alla crescita. In genere i microrganismi che li possiedono sono acquatici e microaerofili o anaerobi.")
14
Granuli di fosfato e zolfo
• Molti organismi accumulano granuli di polifosfato (granuli di volutina o granuli metacromatici) in quanto esso è un nutriente limitato in natura. I granuli sono formati da lunghe catene di fosfato. • I batteri fotosintetici che non producono ossigeno spesso usano lo zolfo come fonte di elettroni e quindi alcuni di loro accumulano globuli di zolfo.
 in quanto esso è un nutriente limitato in natura. I granuli sono formati. da lunghe catene di fosfato. • I batteri fotosintetici che non producono ossigeno spesso usano lo zolfo come fonte di. elettroni e quindi alcuni di loro accumulano globuli di zolfo.")
15
Vacuoli gassosi I batteri dotati di v. gassosi possono regolare la propria profondità di galleggiamento, in modo che sia quella ottimale per soddisfare le loro esigenze d’intensità luminosa, concentraz. di O2 e quantità di nutrienti. Questi batteri possono scendere a fondo semplicemente collassando le vescicole o risalire in seguito alla sintesi di nuove strutture.

16
II materiale genetico procariotico è situato in una regione di forma irregolare, chiamata NUCLEOIDE.
La cellula procariotica di solito contiene una sola molecola di DNA, circolare a doppio filamento, ma in alcune specie il cromosoma è lineare. Recentemente si è scoperto che alcuni batteri come Vibrio cholerae possiedono più di un cromosoma. E’ possibile isolare nucleoidi purificati. La loro analisi chimica dimostra che sono composti per il 60% circa del peso di DNA, per il 30% di RNA e per il restante 10% di proteine. Il E. coli, il DNA, circolare misura approssimativamente 1400mm ed è circa da 230 a 700 volte la lunghezza della cellula.

17
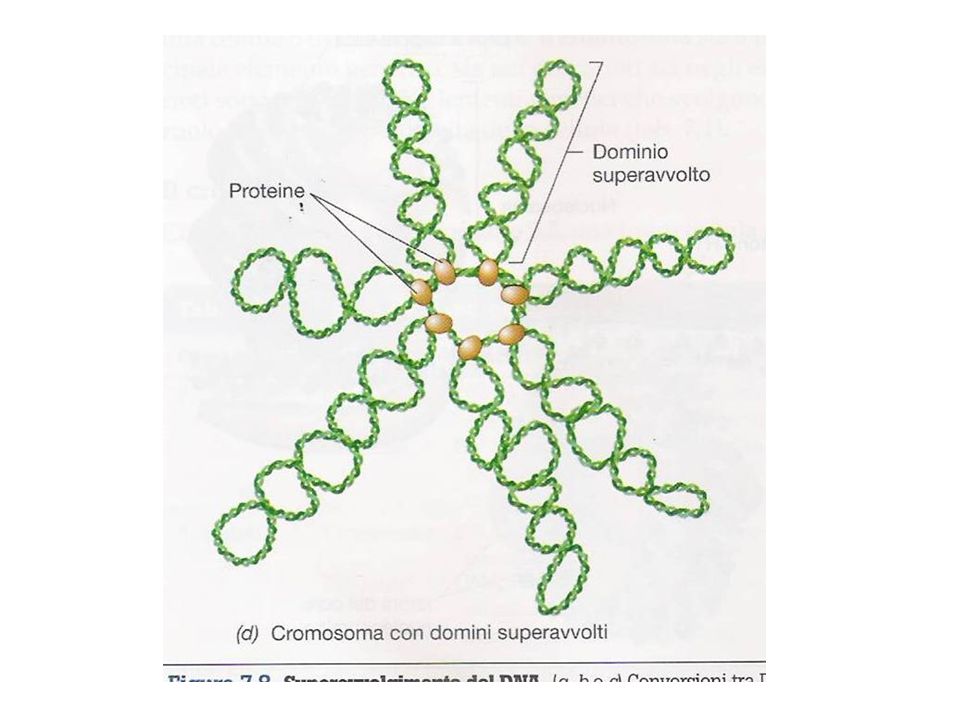
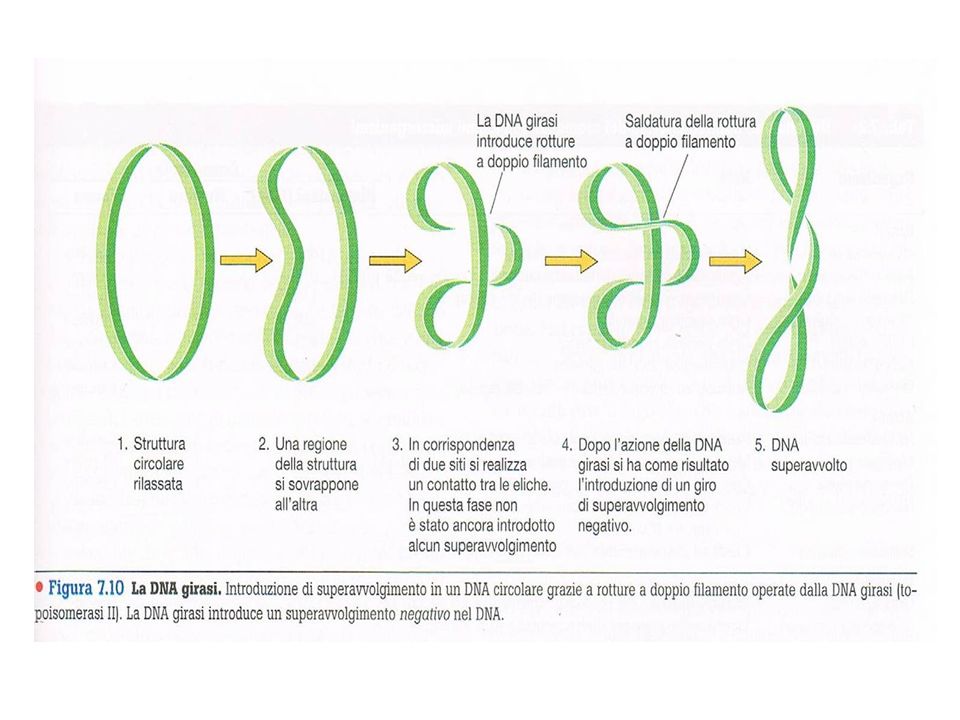
Un cromosoma batterico, quasi sempre di forma circolare, covalentemente chiuso, contenente geni essenziali (noti come geni housekeeping) per la vita dell’organismo, localizzato nel nucleoide. Al DNA sono associate proteine non istoniche che rendono stabile il superavvolgimento (supercoiling) del DNA. Il DNA deve essere superavvolto per poter essere impacchettato all’interno della cellula batterica. Nei Batteri (e nella maggior parte degli Archea) è presente un enzima, DNA girasi (topoisomerasi II), in grado di introdurre nel DNA superavvolgimenti negativi: il DNA si attorciglia attorno al suo asse in direzione opposta a quella della doppia elica destrorsa
 è presente un enzima, DNA girasi (topoisomerasi II), in grado di introdurre nel DNA superavvolgimenti negativi: il DNA si attorciglia attorno al suo asse in direzione opposta a quella della doppia elica destrorsa.")
18
Un altro enzima, topoisomerasi I, è in grado di rimuovere il superavvolgimento del DNA introducendoun taglio (nick) in un singolo filamento della doppia elica del DNA e provocando così la rotazione di un filamento attorno all’altro.Per evitare che ogni rottura provochi il passaggio dell’intero cromosoma dallo stato superavvolto a quello rilassato, il cromosoma batterico è suddiviso in domini di superavvolgimento, isolati da proteine. Un taglio di uno di questi domini non provoca il rilassamento degli altri. In alcuni Archea ipertermofili è presente una girasi inversa in grado di introdurre superavvolgimenti positivi, che avrebbero un ruolo importante nel proteggere il DNA dalla denaturazione.

19
Soltanto alla fine degli anni cinquanta alcuni studi suggerirono che in realtà il genoma batterico si presentasse anch'esso in uno stato condensato e in associazione con delle proteine, dette istoni-simili, implicate nell'organizzazione strutturale del DNA. Le principali sono: HU, H-NS, IHF E FIS.

22
La doppia elica è DESTRORSA, cioè si avvolge in senso orario intorno all’asse

23
Soltanto alla fine degli anni cinquanta alcuni studi suggerirono che in realtà il genoma batterico si presentasse anch'esso in uno stato condensato e in associazione con delle proteine, dette istoni-simili, implicate nell'organizzazione strutturale del DNA. Le principali sono: HU, H-NS, IHF E FIS.

24
DNA extracromosomico: I PLASMIDI

25
In aggiunta al materiale genetico presente nel nucleoide, molti procarioti (nonché alcuni lieviti e altri funghi) contengono molecole di DNA extracromosomico dette PLASMIDI

26
Caratteristiche dei Plasmidi:
I plasmidi sono Elementi genetici accessori: materiale genetico extracromosomico DNA bicatenario, generalmente di forma circolare, in forma superavvolta si trovano all’interno della cellula come “DNA libero”, con dimensioni: tra e bp (dimensioni tipiche di un cromosoma batterico: da 3 a bp contro circa i 3miliardi di nucleotidi in una cellula eucariotica). Contengono un limitato numero di geni (meno di 30) Non sono indispensabili, ma conferiscono alla cellula batterica ospite importanti vantaggi selettivi
. Contengono un limitato numero di geni (meno di 30) Non sono indispensabili, ma conferiscono alla cellula batterica ospite importanti vantaggi selettivi.")
27
Il genoma batterico medio
Genoma=insieme dei geni di un organismo 1 gene medio=1000 nucleotidi (regioni non-codificanti escluse)
")
28
PLASMIDI Dimensioni dei Plasmidi: tra e bp (DNA genomico: bp)
")
29
Caratteristiche dei Plasmidi:
Possono essere presenti da uno a centinaia di copie all’interno della cellula batterica capaci di replicazione indipendente. Gli enzimi coinvolti nella replicazione dei plasmidi sono quelli coinvolti nella replicazione del cromosoma, quindi il plasmide contiene solo i geni per il controllo dell’inizio della replicazione e non per la replicazione; alcuni plasmidi sono coniugativi, altri non coniugativi Alcune cellule batteriche possono contenere diversi tipi di plasmidi. Quando un plasmide viene trasferito in una cellula che già contiene un altro plasmide, a volte succede che il secondo plasmide viene perso durante la successiva replicazione della cellula: in questo caso i due plasmidi vengono detti incompatibili. Alcuni plasmidi, detti EPISOMI , sono in grado di integrarsi nel cromosoma batterico.

30
Principali tipi di plasmidi:
Plasmidi R : responsabili della resistenza agli antibiotici (enzimi inattivanti) Plasmidi Col : produzione di batteriocine, sostanze che inibiscono o uccidono ceppi della stessa specie o specie strettamente correlate Plasmidi di virulenza : produzione di tossine (emolisina ed enterotossina dell’E.coli) o fattori di colonizzazione (fimbrie) Plasmidi metabolici : produzione di enzimi degradativi Plasmidi F : per la fertilità, responsabili della formazione del pilo sessuale nella coniugazione
 Plasmidi Col : produzione di batteriocine, sostanze che inibiscono o uccidono ceppi della stessa specie o specie strettamente correlate. Plasmidi di virulenza : produzione di tossine (emolisina ed enterotossina dell’E.coli) o fattori di colonizzazione (fimbrie) Plasmidi metabolici : produzione di enzimi degradativi. Plasmidi F : per la fertilità, responsabili della formazione del pilo sessuale nella coniugazione.")
31
Le batteriocine sono molecole peptidiche di sintesi ribosomale che possono anche essere
associate a lipidi o carboidrati. Sebbene, alcune batteriocine prodotte da batteri Gram-positivi (Lactobacillus, Lactococcus) possiedono spettri di inibizione limitati ad alcuni ceppi appartenenti alla stessa specie del microrganismo produttore, la maggior parte di esse mostra un ampio spettro d’azione contro diverse specie batteriche sia Gram-positive che Gram-negative.
 possiedono spettri di inibizione limitati ad alcuni ceppi. appartenenti alla stessa specie del microrganismo produttore, la maggior parte di esse mostra un ampio spettro d’azione contro diverse specie batteriche sia Gram-positive che Gram-negative.")
32
Plasmide R

33
Plasmide R GENI PER LA RESISTENZA BATTERICA AGLI ANTIBIOTICI
Geni per il trasferimento

34
Plasmide F È costituito da circa 22 geni.
La formazione del pilo F è sotto il controllo di 12 geni Geni per il trasferimento Punto di origine del trasferimento

35
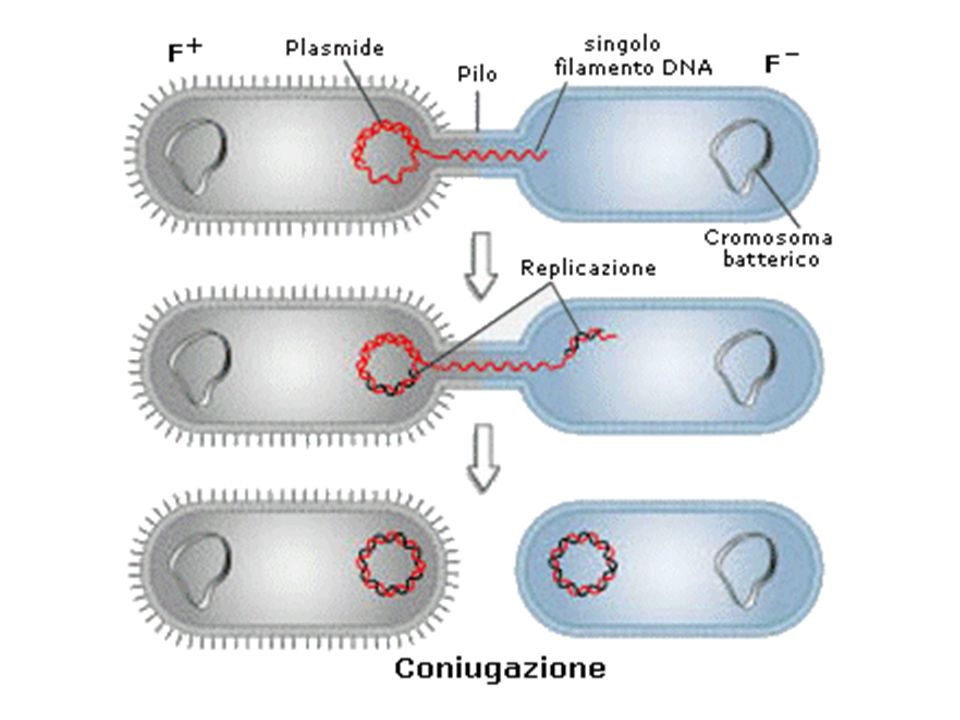
CONIUGAZIONE Processo di trasferimento di materiale genetico che comporta il contatto cellula-cellula Coinvolge una cellula, definita donatore, che contiene un plasmide coniugativo F (cellula F+) e una cellula ricevente che ne è priva (cellula F-) Nel plasmide sono presenti geni per la coniugazione tra cui quelli per la sintesi di una struttura di superficie, detta pilo sessuale i pili stabiliscono dei contatti con le cellule recipienti riconoscendo dei recettori specifici sulla loro superficie e poi si ritraggono, per depolimerizzazione, costringendo così le due cellule ad unirsi Dopo che il contatto si è stabilizzato, probabilmente per fusione delle membrane, il DNA viene trasferito da una cellula all’altra.
 e una cellula ricevente che ne è priva (cellula F-) Nel plasmide sono presenti geni per la coniugazione tra cui quelli per la sintesi di una struttura di superficie, detta pilo sessuale. i pili stabiliscono dei contatti con le cellule recipienti riconoscendo dei recettori specifici sulla loro superficie e poi si ritraggono, per depolimerizzazione, costringendo così le due cellule ad unirsi. Dopo che il contatto si è stabilizzato, probabilmente per fusione delle membrane, il DNA viene trasferito da una cellula all’altra.")
36
CONIUGAZIONE Richiede uno stretto contatto cellula –cellula Nella cellula donatrice sono presenti plasmidi coniugativi I plasmidi coniugativi presentano geni per il trasferimento e per la sintesi del pilo F

37
Coniugazione F+ - F-

38
1946: Lederberg e Tatum La coniugazione avviene attraverso il contatto fisico tra 2 cellule, la cellula F+ (donatrice)che presenta F libero nel citoplasma e la cellula F- (ricevente) priva di F
 priva di F.")
40
I pili (o fimbrie) sono strutture filamentose, formate da subunità proteiche (PILINA) organizzate in una struttura elicoidale: possono essere cavi (permettono il passaggio di DNA nei pili coniugativi) o non cavi (permettono l’ancoraggio a substrati nutritivi).
 o non cavi (permettono l’ancoraggio a substrati nutritivi).")
41
I pili F sono prodotti soltanto dalle cellule dotate di un fattore di fertilità, contenuto in genere nel cromosoma plasmidico. Il ruolo del pilo F o coniugativo è quello di fungere da veicolo per il trasferimento di geni da una cellula batterica ad un’altra. I pili per il loro limitato spessore sono visibili al M.E.

42
I termini fimbrie e pili sono spesso usati come sinonimi, ma in realtà è bene distinguere i due termini Le FIMBRIE consentono al microrganismo di aderire a superfici, come i tessuti ANIMALI nel caso dei batteri patogeni*, o di formare pellicole o biofilm sulle superfici. *Molto noto tra questi è il batterio patogeno Salmonella typhimurium. I

43
I PILI sono strutture simili alle fimbrie ma generalmente più lunghe e presenti sulla cellula in una o al massimo poche copie. Essi sono visibili al ME quando rivestiti da particelle virali che li utilizzano come recettori. Sebbene i pili possano essere coinvolti nei fenomeni di adesione come le fimbrie, i pili sono chiaramente implicati nel processo di coniugazione.

44
Batteri Gram negativi Batteri Gram-positivi FLAGELLO

45
In E. coli e nella maggioranza dei batteri Gram-, il corpo basale consiste di due coppie di anelli connesse ad un albero centrale. Gli anelli più esterni, L e P, sono associati rispettiv. con lo strato di lipopolisaccaridi e con quello di peptidoglicano. L’anello interno M prende contatto con la membrana plasmatica.Anche l’anello S prende contatti con la parete cellulare. Gram + Gram -

46
Nei batteri gram + il corpo basale consta di due soli anelli: uno interno collegato, M, collegato alla membrana plasmatica, ed uno esterno, S, probabilmente associato al peptidoglicano.

47
La mobilità può essere considerata un vantaggio selettivo
che favorisce la sopravvivenza. I batteri flagellati sono in grado di rispondere a stimoli chimici e luminosi. Tali comportamenti sono indicati coi termini rispettivamente di: CHEMIOTASSI FOTOTASSI


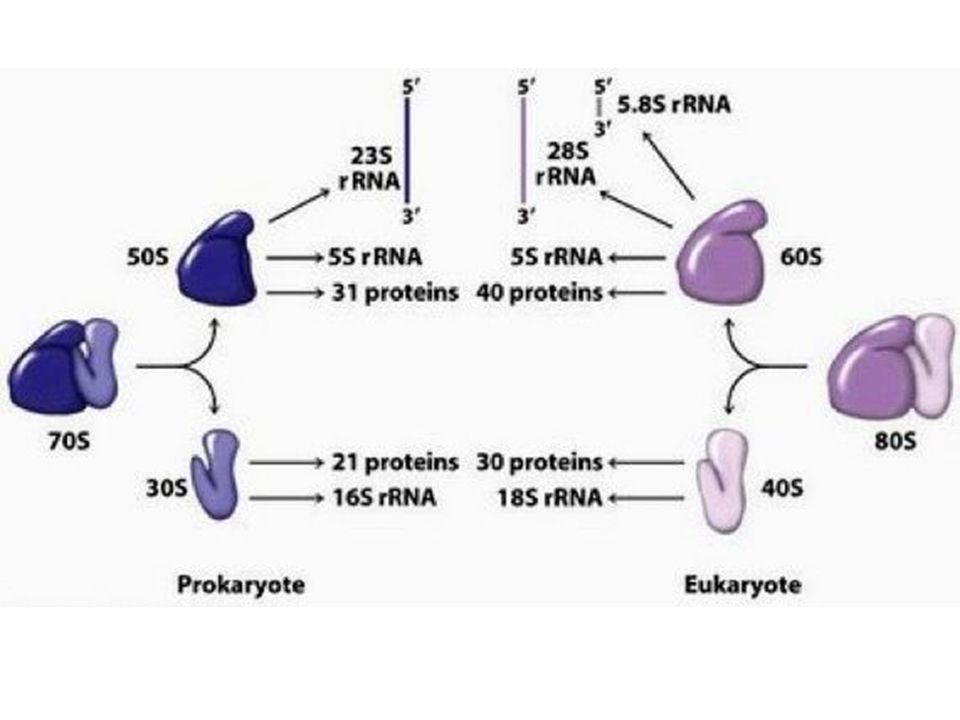
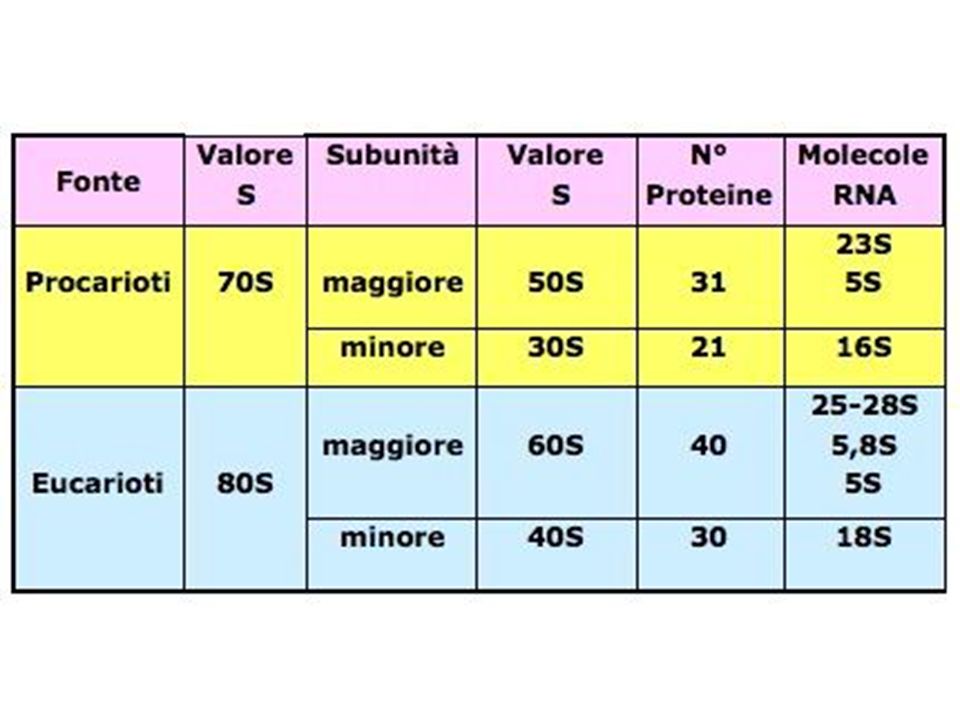
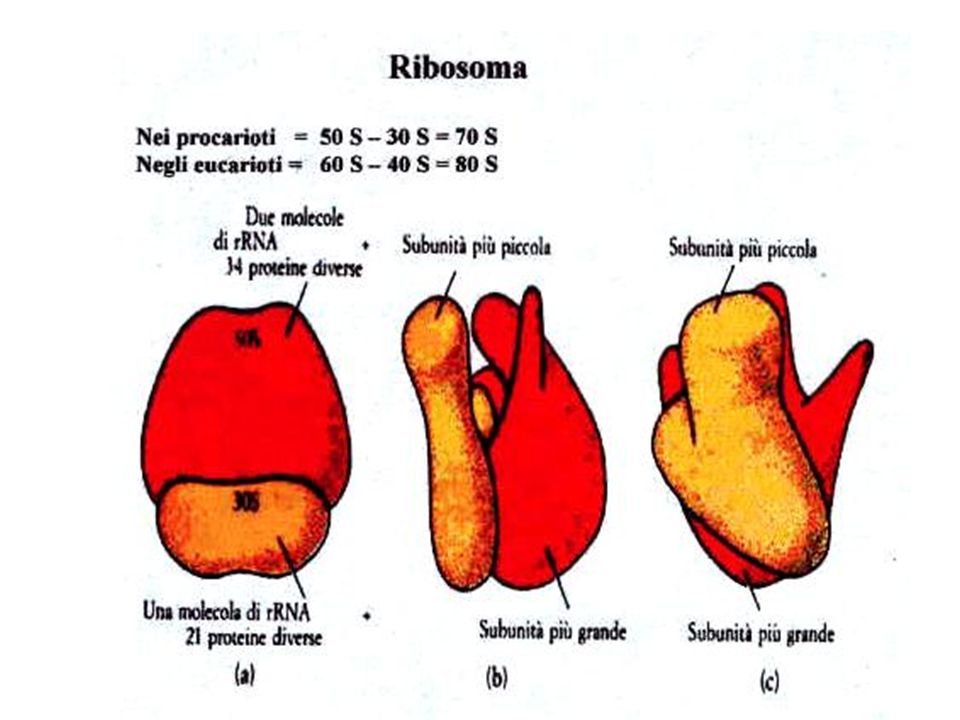
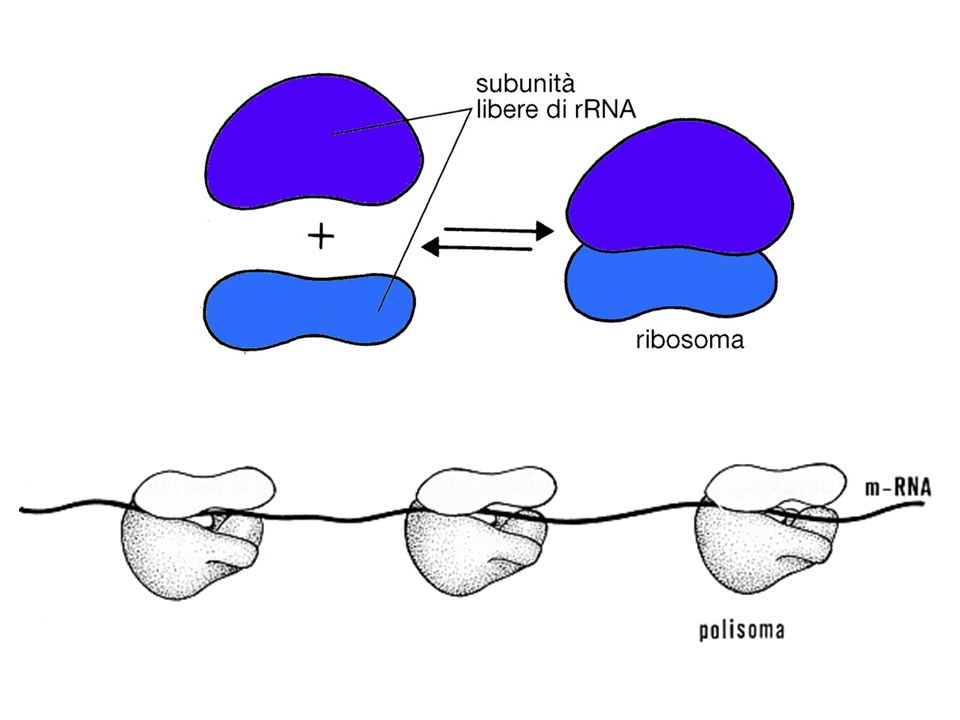


















>")
 è l'unità fondamentale di tutti gli organismi viventi, la più piccola struttura ad essere classificabile.>")