Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
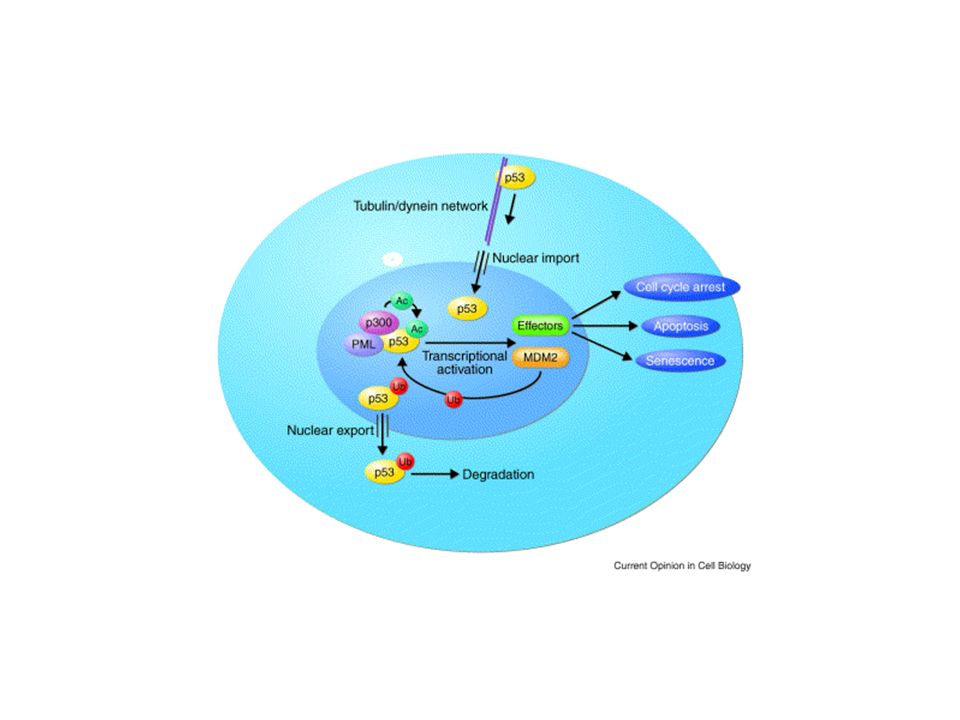
p53 has a key role in integrating the cellular responses (pink boxes) to different types of stress (blue boxes). Activation of p53 can result in a number of cellular responses, and it is possible that different responses are induced by different stress signals. There is evidence that p53 can play a part in determining which response is induced through differential activation of target-gene expression. Although the importance of these responses to tumour suppression is clear, previously unanticipated contributions of these responses to other aspects of human health and disease are being uncovered. The role of p53 in tumour suppression, development and ageing is likely to depend on which cellular response is activated and on the context in which the activation occurs.

2
p53 activation. p53 is activated by several types of stress and induces the expression of several proteins that are involved in the inhibition of cell proliferation or in promoting cell death by apoptosis. In tumour cells, the growth-arresting and apoptotic effects of p53 might be lost owing to the mutation of genes downstream of p53, to mutations in the p53 gene, or to mutations or alterations in the levels of proteins that modulate the p53 function

3
Regulation of p53, a stress-regulated transcription factor that co-ordinately induces or represses sets of gene products in response to changes in the cellular microenvironment. P53 co-operates with the transcriptional activator p300 to induce sets of gene products implicated in growth control (left panel). P53 co-operates with de-acetylases to repress gene expression of proteins implicated in proliferation and survival (right panel). P53 is often mutated in human cancers leading to a protein that can not respond to signalling cues and that can not regulate gene products that orchestrate the stress response. The mutation in p53 unfolds the protein, stabilizes it in the nucleus where it can be bound by molecular chaperones, and creates a protein that often has an oncogenic gain-of-function property.
. P53 co-operates with de-acetylases to repress gene expression of proteins implicated in proliferation and survival (right panel). P53 is often mutated in human cancers leading to a protein that can not respond to signalling cues and that can not regulate gene products that orchestrate the stress response. The mutation in p53 unfolds the protein, stabilizes it in the nucleus where it can be bound by molecular chaperones, and creates a protein that often has an oncogenic gain-of-function property..")
4
miR-34 is a direct transcriptional target of p53, which in turn downregulates genes required for proliferation and survival. Along with other p53 targets, such as p21 and BAX, miR-34-family miRNAs promote growth arrest and cell death in response to cancer related stress. ATM, ataxia talangiectasia mutated; ATR, ataxia telengiectasia and RAD3-related; CDK, cyclin-dependent kinase; CHK, checkpoint kinase

5
PUMA NOXA

6
b | Classes of ubiquitin ligases: single RING-finger E3s; HECT E3s; and multi-subunit RING-finger E3s, exemplified by the SCF complexes. Only HECT-domain E3s form a covalent bond with ubiquitin during polyubiquitylation of their target proteins. Specific E3s discussed in this review are indicated.

8
INACTIVATION OF THE p53 PATHWAY
Dominant-negative missense p53 gene mutation Upstream tumour suppressor gene mutation (ATM) Oncogene amplification (MDM2) Viral oncogene amplification (HPV E6)
 Oncogene amplification (MDM2) Viral oncogene amplification (HPV E6)")
9
The major functional domains of the p53 protein are shown, including the N-terminal transactivation domains, the central sequence-specific DNA-binding domain and the C-terminal regulatory domain. p53 is subject to numerous post-transcriptional modifications, including phosphorylation, acetylation, methylation and modification with ubiquitin-like proteins, that can affect the function and stability of p53. Phosphatases, de-acetylases and de-ubiquitylating enzymes have been identified that can reverse most of these modifications. Most of the point mutations found in naturally occurring cancers occur in the central DNA-binding domain, and the position of the hotspots for these mutations are indicated by the orange lightning bolts. The p53-related proteins p63 and p73 show a similar overall structure, although some isoforms of these p53 relatives also contain a C-terminal sterile -motif (SAM) domain. Multiple isoforms of each of these proteins have now been described16. Download file

10
Structure and expression of p53 family members
Structure and expression of p53 family members. (a) Structure of p53, p63 and p73 transcription units. Numbered boxes indicate exons, and black shading denotes untranslated sequences. The approximate regions encoding the transactivation (TA) domain (light blue), N-specific region (green), DNA-binding domain (red), oligomerization domain (yellow), sterile alpha motif (SAM, grey), and transactivational inhibitory domain (TID, orange) are indicated. Distinct transcription start sites are indicated by arrows. N-terminal alternative splicing for p53 and p73 are indicated by dotted lines, and C-terminal splicing events for all p53 family members are indicated by solid lines and Greek letter designation. (b) Protein domains of p53 family members. All three family members share a homologous DNA-binding domain and oligomerization domain (oligo). The TA domain is shared by p53, TAp63, and TAp73 isoforms. TAp63 /TAp73 isoforms most closely resemble p53. N-terminally truncated N isoforms possess unique N-terminal sequences. Alpha isoforms of p63 and p73 possess a C-terminal SAM domain followed by a transactivational inhibitory domain (TID). Other isoforms of p53, p63 and p73 are not shown.
 Structure of p53, p63 and p73 transcription units. Numbered boxes indicate exons, and black shading denotes untranslated sequences. The approximate regions encoding the transactivation (TA) domain (light blue), N-specific region (green), DNA-binding domain (red), oligomerization domain (yellow), sterile alpha motif (SAM, grey), and transactivational inhibitory domain (TID, orange) are indicated. Distinct transcription start sites are indicated by arrows. N-terminal alternative splicing for p53 and p73 are indicated by dotted lines, and C-terminal splicing events for all p53 family members are indicated by solid lines and Greek letter designation. (b) Protein domains of p53 family members. All three family members share a homologous DNA-binding domain and oligomerization domain (oligo). The TA domain is shared by p53, TAp63, and TAp73 isoforms. TAp63 /TAp73 isoforms most closely resemble p53. N-terminally truncated N isoforms possess unique N-terminal sequences. Alpha isoforms of p63 and p73 possess a C-terminal SAM domain followed by a transactivational inhibitory domain (TID). Other isoforms of p53, p63 and p73 are not shown.")
11
Structure and expression of p53 family members
Structure and expression of p53 family members. (a) Structure of p53, p63 and p73 transcription units. Numbered boxes indicate exons, and black shading denotes untranslated sequences. The approximate regions encoding the transactivation (TA) domain (light blue), N-specific region (green), DNA-binding domain (red), oligomerization domain (yellow), sterile alpha motif (SAM, grey), and transactivational inhibitory domain (TID, orange) are indicated. Distinct transcription start sites are indicated by arrows. N-terminal alternative splicing for p53 and p73 are indicated by dotted lines, and C-terminal splicing events for all p53 family members are indicated by solid lines and Greek letter designation. (b) Protein domains of p53 family members. All three family members share a homologous DNA-binding domain and oligomerization domain (oligo). The TA domain is shared by p53, TAp63, and TAp73 isoforms. TAp63 /TAp73 isoforms most closely resemble p53. N-terminally truncated N isoforms possess unique N-terminal sequences. Alpha isoforms of p63 and p73 possess a C-terminal SAM domain followed by a transactivational inhibitory domain (TID). Other isoforms of p53, p63 and p73 are not shown.
 Structure of p53, p63 and p73 transcription units. Numbered boxes indicate exons, and black shading denotes untranslated sequences. The approximate regions encoding the transactivation (TA) domain (light blue), N-specific region (green), DNA-binding domain (red), oligomerization domain (yellow), sterile alpha motif (SAM, grey), and transactivational inhibitory domain (TID, orange) are indicated. Distinct transcription start sites are indicated by arrows. N-terminal alternative splicing for p53 and p73 are indicated by dotted lines, and C-terminal splicing events for all p53 family members are indicated by solid lines and Greek letter designation. (b) Protein domains of p53 family members. All three family members share a homologous DNA-binding domain and oligomerization domain (oligo). The TA domain is shared by p53, TAp63, and TAp73 isoforms. TAp63 /TAp73 isoforms most closely resemble p53. N-terminally truncated N isoforms possess unique N-terminal sequences. Alpha isoforms of p63 and p73 possess a C-terminal SAM domain followed by a transactivational inhibitory domain (TID). Other isoforms of p53, p63 and p73 are not shown.")
12
The E3 ligase ITCH regulates p73 stability
The E3 ligase ITCH regulates p73 stability. (A) The PR domain of p73 physically interacts with the WW domain of the E3 ligase Itch. Itch is an ubiquitin E3 ligase, part of the Nedd4 family, containing an HECT domain. Itch protein includes several domains: the C2 domain is a calcium-dependent region able to interact also with lipids, the WW domain is a protein-protein interacting region, able to bind PY motif, such as the PR region of p73, the HECT domain is the catalytic domain transferring ubiquitin to the specific substrate, in this case p73. Itch degrades p73, p63 and also c-Jun, Jun-B, Notch and Flip, therefore suggesting a relevant role in cancer development and chemosensitivity. A natural mouse knockout of Itch is available, the 18H aguty mice, that shows specific immune defects and atrophin-related defects. (B) Itch specifically binds (left panel), ubiquitylates (central panel) and degrades (right panel) p73. These data are extracted from Rossi et al. (2005); relevant controls are omitted to facilitate the presentation.
 The PR domain of p73 physically interacts with the WW domain of the E3 ligase Itch. Itch is an ubiquitin E3 ligase, part of the Nedd4 family, containing an HECT domain. Itch protein includes several domains: the C2 domain is a calcium-dependent region able to interact also with lipids, the WW domain is a protein-protein interacting region, able to bind PY motif, such as the PR region of p73, the HECT domain is the catalytic domain transferring ubiquitin to the specific substrate, in this case p73. Itch degrades p73, p63 and also c-Jun, Jun-B, Notch and Flip, therefore suggesting a relevant role in cancer development and chemosensitivity. A natural mouse knockout of Itch is available, the 18H aguty mice, that shows specific immune defects and atrophin-related defects. (B) Itch specifically binds (left panel), ubiquitylates (central panel) and degrades (right panel) p73. These data are extracted from Rossi et al. (2005); relevant controls are omitted to facilitate the presentation.")
13
Acetylation Fig. 7. Regulation of p73 stability and transcriptional activation. (A) The p73 protein is degraded via the ubiquitinin-proteasome degradation pathway, regulated by the E3 ligase Itch. Additional E3 ligases are also involved, even though their identification and specificity is still under evaluation. The PML protein is able to attenuate protein degradation by diverting the protein to the transcription pathway. (B) The transcription pathway of p73 follows a very strict regulation involving several steps: (i) phosphorylation by a specific kinase such as MAP p38 and c-Abl, (ii) prolyl-isomerization by the enzyme Pin-1, (iii) recruitment and activation by PML into the PML-nuclear body and (iv) acethylation by the CBP/p300 complex. The protein that is phosphorylated, pro-isomerised and acethylated is transcriptionally competent. During this pathway the protein seems less sensitive to the degradation process.
 The p73 protein is degraded via the ubiquitinin-proteasome degradation pathway, regulated by the E3 ligase Itch. Additional E3 ligases are also involved, even though their identification and specificity is still under evaluation. The PML protein is able to attenuate protein degradation by diverting the protein to the transcription pathway. (B) The transcription pathway of p73 follows a very strict regulation involving several steps: (i) phosphorylation by a specific kinase such as MAP p38 and c-Abl, (ii) prolyl-isomerization by the enzyme Pin-1, (iii) recruitment and activation by PML into the PML-nuclear body and (iv) acethylation by the CBP/p300 complex. The protein that is phosphorylated, pro-isomerised and acethylated is transcriptionally competent. During this pathway the protein seems less sensitive to the degradation process.")
14
b | Classes of ubiquitin ligases: single RING-finger E3s; HECT E3s; and multi-subunit RING-finger E3s, exemplified by the SCF complexes. Only HECT-domain E3s form a covalent bond with ubiquitin during polyubiquitylation of their target proteins. Specific E3s discussed in this review are indicated.

15
Figure 1. p63 deficiency induces cellular senescence
Figure 1. p63 deficiency induces cellular senescence. p63 deficiency induces the expression p16Ink4a and PML.8 The enhanced p16Ink4a and PML promote SAHF formation and senescence. Given the finding that ΔNp63α inhibits GSK3β activity,55 p63 deficiency potentially enhances GSK3β activity. The increased activity of GSK3β can regulate p53 and the chromatin regulator HIRA, thus regulating senescence. On the other hand, p63 can inhibit the transactivation of cell cycle inhibitors p21.39 Thus, p63 deficiency can cause upregulation of p21. p21 has been proposed to be a key effector of p53-mediated senescence by inhibiting Cyclin E-Cdk2.26

16
Figure 2. ΔNp63α promotes cellular proliferation
Figure 2. ΔNp63α promotes cellular proliferation. ΔNp63α can inhibit GSK3β activity through interacting with and inhibiting the protein phosphotase PP2A. The decreased GSK3β activity leads to β-catenin nuclear accumulation, thus its pro-proliferative function.55 ΔNp63α is induced by EGF through the PI3K pathway.52 ΔNp63α can promote cellular proliferation by inducing oncogenic proteins such as Hsp70.51 On the other hand, ΔNp63α can inhibit the activity of p539 and the transactivation of cell cycle inhibitors such as p21 and14-3-3σ,39 thus enhancing cellular proliferation.

17
Figure 3. Model for p63 as a tumor suppressor
Figure 3. Model for p63 as a tumor suppressor. (A) Down regulation or loss of TAp63 and/ or overexpression of ΔNp63 leads to inhibition of the functions of TAp63, p53, and TAp73 leading to the development of an invasive and metastatic tumor. (B) Mutant p53 binds to TAp63 and TAp73 inhibiting their function leading to the development of an invasive and metastatic tumor.
 Down regulation or loss of TAp63 and/ or overexpression of ΔNp63 leads to inhibition of the functions of TAp63, p53, and TAp73 leading to the development of an invasive and metastatic tumor. (B) Mutant p53 binds to TAp63 and TAp73 inhibiting their function leading to the development of an invasive and metastatic tumor.")
18
Pathways involving the p53 family functional network in squamous cell carcinoma. DNA damage and oncogenic stress (through p14arf) trigger p53 activation, leading to selective pressure for its mutation or loss to avoid apoptosis. Similarly, DNA damage and potentially oncogenic stress induce TAp73 expression and activation. TAp73 activity can be suppressed through overexpression of Np63 or certain missense p53 mutants (mtp53). Alternatively, apoptosis can be averted in Np63 -negative tumors through overexpression of bcl-2, which inhibits the proapoptotic function of TAp73 target genes.
 trigger p53 activation, leading to selective pressure for its mutation or loss to avoid apoptosis. Similarly, DNA damage and potentially oncogenic stress induce TAp73 expression and activation. TAp73 activity can be suppressed through overexpression of Np63 or certain missense p53 mutants (mtp53). Alternatively, apoptosis can be averted in Np63 -negative tumors through overexpression of bcl-2, which inhibits the proapoptotic function of TAp73 target genes..")
19
p63/p73 pathway mediates cisplatin sensitivity in squamous carcinoma cells. In proliferating tumor cells, Np63 inhibits the proapoptotic transcriptional activity of TAp73 through both direct physical interaction and through direct binding to the promoters of TAp73 target genes. Cisplatin treatment induces Np63 downregulation and TAp73 stabilization, thereby activating the TAp73-dependent apoptotic program. Phosphorylation of TAp73 by c-Abl and potentially other kinases is important for its activation following DNA damage. Phosphorylation may also contribute to downregulation of Np63 in this context. Note that proteins are shown schematically as dimers but in fact are thought to bind DNA as tetramers.

20
421 421 240 240 1620 1620 (A) wildtype or (B) structural mutants
DNA contact mutants (B) structural mutants 421 421 240 240 1620 1620 1620 421 240 1620 421 240
 structural mutants")
21
Figure 1. Mutant p53 is stabilized by unknown mechanisms in tumors
Figure 1. Mutant p53 is stabilized by unknown mechanisms in tumors. Sufficient mutant p53 is then available to bind and inactivate the functions of wild type p53, p63 and p73.

22
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO MUTAZIONI DI p53 terapia genica con p53 wildtype

23
pRb E1A p53 E1B 55kDa E1B E1B 19kDa E1A E1B ADENOVIRUS
ARRESTO del CICLO CELLULARE pRb E1A ADENOVIRUS p53 APOPTOSI DEGRADAZIONE E1B 55kDa E1B E1B 19kDa APOPTOSI E1A E1B rAd-p53 SOSTITUITI DA p53wt

24
PROBLEMI CONNESSI CON L’USO DI ADENOVIRUS
COME VETTORI PER LA TERAPIA GENICA espressione genica transiente (14-21 gg) prevalenza di anticorpi neutralizzanti accessibilità alle cellule tumorali
 prevalenza di anticorpi neutralizzanti. accessibilità alle cellule tumorali.")
25
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO MUTAZIONI DI p53 terapia genica con p53 wildtype virus oncolitici a replicazione selettiva (Onyx-015)
")
26
pRb E1A p53 E1B 55kDa E1B E1B 19kDa pRb E1A E1B DEGRADAZIONE
ADENOVIRUS E1B 19kDa APOPTOSI ONYX-015 E1A pRb E1B

27
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO MUTAZIONI DI p53 terapia genica con p53 wildtype virus oncolitici a replicazione selettiva (Onyx-015) imitazione dei prodotti di geni regolati da p53 (p21waf1; Bax)
 imitazione dei prodotti di geni regolati da p53 (p21waf1; Bax)")
28
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO MUTAZIONI DI p53 terapia genica con p53 wildtype virus oncolitici a replicazione selettiva (Onyx-015) imitazione dei prodotti di geni regolati da p53 recupero farmacologico della proteina p53 mutata: inibitori di Hsp90 altri
 imitazione dei prodotti di geni regolati da p53. recupero farmacologico della proteina p53 mutata: inibitori di Hsp90. altri.")
29
Correcting mutant p53 to perform wild-type functions
Correcting mutant p53 to perform wild-type functions. Specific p53 mutants maybe altered to perform wild type-mediated growth inhibition through exposure to the activating molecules (Section 3 and subsections within). Haupt S Semin Cancer Biol 2004
. Haupt S Semin Cancer Biol")
30
(p53 reactivation and induction of massive apoptosis)
PRIMA-1 (p53 reactivation and induction of massive apoptosis)
")
31
CP does not bind the core domain of recombinant wt or mutant p53 in vitro. It has no effect on the interaction between p53 and Mdm2. Hypothesis: CP is proposed to interact with newly synthetized p53 in vivo, blocking its ubiquitination and degradation

32
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO p53 wildtype attivazione della risposta di p53

33
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO p53 wildtype attivazione della risposta di p53 blocco dell’espressione di E6

34
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO p53 wildtype attivazione della risposta di p53 blocco dell’espressione di E6 inibizione dell’espressione di Mdm2 inibizione della funzione di Mdm2

35
Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).
![Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).](http://slideplayer.it/slide/579113/2/images/35/Structural+formula+of+PRIMA-1+%282%2C2-bis%28hydroxy-methyl%29-1-azabicyclo%5B2%2C2%2C2%5Doctan-3-one%29..jpg "Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).")
36
Regulation of p53 by MDM2. p53 and MDM2 form an auto-regulatory feedback loop. p53 stimulates the expression of MDM2; MDM2 inhibits p53 activity because it blocks its transcriptional activity, favours its nuclear export and stimulates its degradation. Different cellular signals, such as DNA-damage or oncogene activation, induce p53 activation. DNA damage favours p53 phosphorylation, preventing its association with MDM2. Activated oncogenes activate the ARF protein, which prevents the MDM2-mediated degradation of p53. Similarly, inhibitors of the p53–MDM2 interaction should activate p53 tumour-suppressor activity in tumour cells that express wild-type p53. These compounds, because they bind to MDM2, could also affect the p53-independent activities of MDM2.

37
Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).
![Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).](http://slideplayer.it/slide/579113/2/images/37/Structural+formula+of+PRIMA-1+%282%2C2-bis%28hydroxy-methyl%29-1-azabicyclo%5B2%2C2%2C2%5Doctan-3-one%29..jpg "Structural formula of PRIMA-1 (2,2-bis(hydroxy-methyl)-1-azabicyclo[2,2,2]octan-3-one).")
39
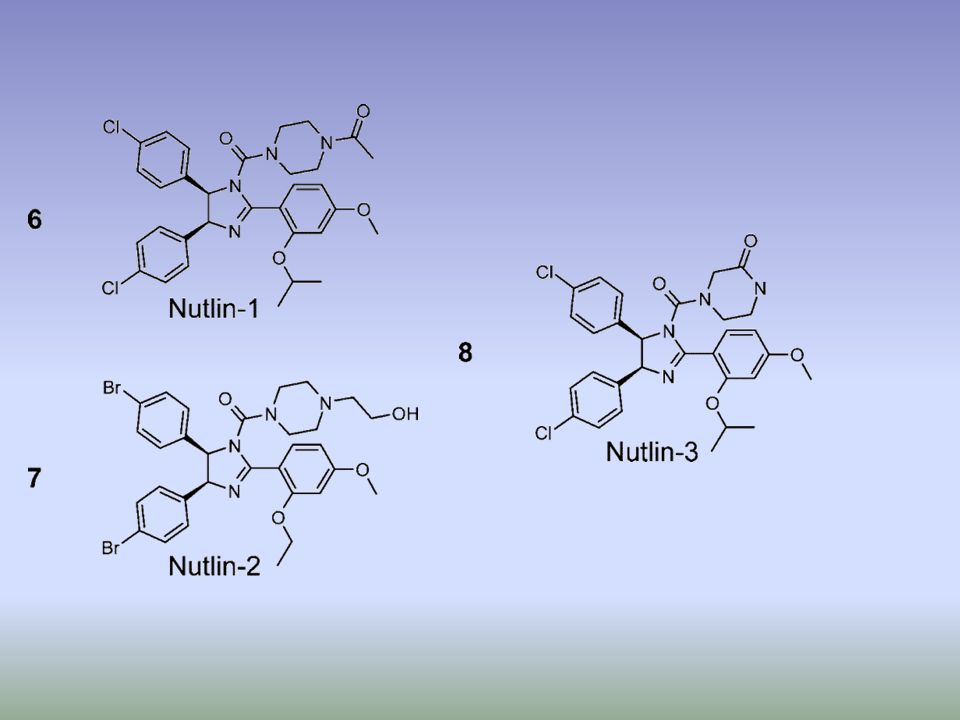
. b | Structure of Nutlin (left) and RITA (right), the first small-molecule inhibitors of the p53–MDM2 interaction73, 75. c | Structure of the Nutlin–p53 complex19. Left: the sides of p53-binding groove in MDM2 (red) is limited by two -helices and a short -sheet while the bottom is formed by two shorter -helices perpendicular to the pocket sides. The amino-terminal domain of p53 (green) is stabilized in -helical conformation upon binding to MDM2 due to a network of hydrogen bonds trapping three crucial residues of p53, Phe19, Trp23 and Leu26. These crucial p53 residues point towards the bottom of the groove. Right: Nutlin prevents interaction between p53 and MDM2 by mimicking the conformation of Phe19, Trp23 and Leu26 of p53 to block the MDM2 p53-interacting domain73. MDM2, double minute 2.
 is limited by two -helices and a short -sheet while the bottom is formed by two shorter -helices perpendicular to the pocket sides. The amino-terminal domain of p53 (green) is stabilized in -helical conformation upon binding to MDM2 due to a network of hydrogen bonds trapping three crucial residues of p53, Phe19, Trp23 and Leu26. These crucial p53 residues point towards the bottom of the groove. Right: Nutlin prevents interaction between p53 and MDM2 by mimicking the conformation of Phe19, Trp23 and Leu26 of p53 to block the MDM2 p53-interacting domain73. MDM2, double minute 2..")
40
Promotion of wt p53 activities through protection from Mdm2-mediated inhibition. Interruption of the p53–Mdm2 interaction can be achieved by treatment with Nutlins, syc-7 and chalcones leading to activation of p53-mediated growth inhibition (Section 2.1). Down regulation of Mdm2 can be achieved through the introduction of anti-sense mRNA directed toward Mdm2. The drug Cp appears to activate wt p53 by preventing p53 ubiquitination and possibly through the induction of DNA damage (Section ). The proteasome inhibitor bortezomib stabilizes p53 by preventing its degradation mediated by Mdm2, possibly by inhibiting p53 ubiquitination ( Section 2.4). Green arrows denote activating pathways while red bars denote inhibition.
. Down regulation of Mdm2 can be achieved through the introduction of anti-sense mRNA directed toward Mdm2. The drug Cp appears to activate wt p53 by preventing p53 ubiquitination and possibly through the induction of DNA damage (Section ). The proteasome inhibitor bortezomib stabilizes p53 by preventing its degradation mediated by Mdm2, possibly by inhibiting p53 ubiquitination ( Section 2.4). Green arrows denote activating pathways while red bars denote inhibition..")
41
STRATEGIE TERAPEUTICHE BASATE SU p53
TUMORI CHE PRESENTANO p53 wildtype attivazione della risposta di p53 blocco dell’espressione di E6 inibizione dell’espressione di Mdm2 inibizione della funzione di Mdm2 blocco del trafficking nucleare di p53 (leptomicina B)
")
43
Leptomycin B (LMB) is an unsaturated branched-chain fatty acid and can undergo a Michael-type addition by Cysteine-529 on the exportin CRM1. This covalent modification inhibits binding between CRM1 and the nuclear export signal (NES)-containing cargo. Nuclear transport as a target for cell growth, Pages Tweeny R. Kau and Pamela A. Silver

44
Some of the points at which p53 can affect metabolic pathways
Some of the points at which p53 can affect metabolic pathways. This is a new and rapidly moving area of research, and the influence of p53 on metabolism is likely to be much broader than illustrated here. In response to nutrient stress, p53 can become activated by AMP kinase (AMPK), promoting cell survival through an activation of the cyclin-dependent kinase inhibitor p21. Other functions of p53 include regulating respiration, through the action of SCO2, or in decreasing the levels of reactive oxygen species (ROS), through the actions of TIGAR (Tp53-inducible glycolysis and apoptosis regulator) or sestrins.
, promoting cell survival through an activation of the cyclin-dependent kinase inhibitor p21. Other functions of p53 include regulating respiration, through the action of SCO2, or in decreasing the levels of reactive oxygen species (ROS), through the actions of TIGAR (Tp53-inducible glycolysis and apoptosis regulator) or sestrins.")
45
p53 functions in the response to both the constitutive stress that is encountered during normal growth and development, and to the acute stress signals that would be associated with oncogenic progression and other types of trauma. In this model, p53 responds to conditions of low or constitutive stress to play an important part in decreasing oxidative damage, and provides repair functions to mend low levels of DNA damage. These activities of p53 contribute to the survival and health of the cell as well as to the prevention of the acquisition of tumorigenic mutations, and might contribute to overall longevity and normal development. By contrast, acute stress that results in a more robust induction of p53 leads to the activation of apoptotic cell death and thereby the elimination of the damaged cell. This function removes cells that have acquired oncogenic alterations, and can contribute to neural tube closure during development, but carries accompanying detrimental effects of stress-induced toxicity, such as radiation sickness, neurodegenerative disease and premature ageing.

46
A recent study from Evan and colleagues33 provides evidence that the immediate p53 response to DNA damage is detrimental and not necessary for tumour suppression. The absence of p53 immediately after DNA damage protects animals from radiation sickness, but does not prevent repair of the DNA damage. Persistent absence of p53 results in tumour development, as expected. However, even transient restoration of p53 activity after the resolution of the initial DNA damage can inhibit tumour development without the deleterious responses, such as widespread apoptosis in lymphoid organs and intestinal epithelium, that occur following the irradiation of mice with fully active p53. These results suggest that the induction of p53 in response to signals that persist beyond DNA damage, such as activated oncogenes, is the key to tumour suppression.

47
pifithrin-

Presentazioni simili




>")





/(+)>")




 DNA lineare>")




