Scaricare la presentazione
La presentazione è in caricamento. Aspetta per favore

1
PREPARAZIONI DI ORGANI ISOLATI IN FARMACOLOGIA

2
in vitro ex vivo ESPERIMENTI

3
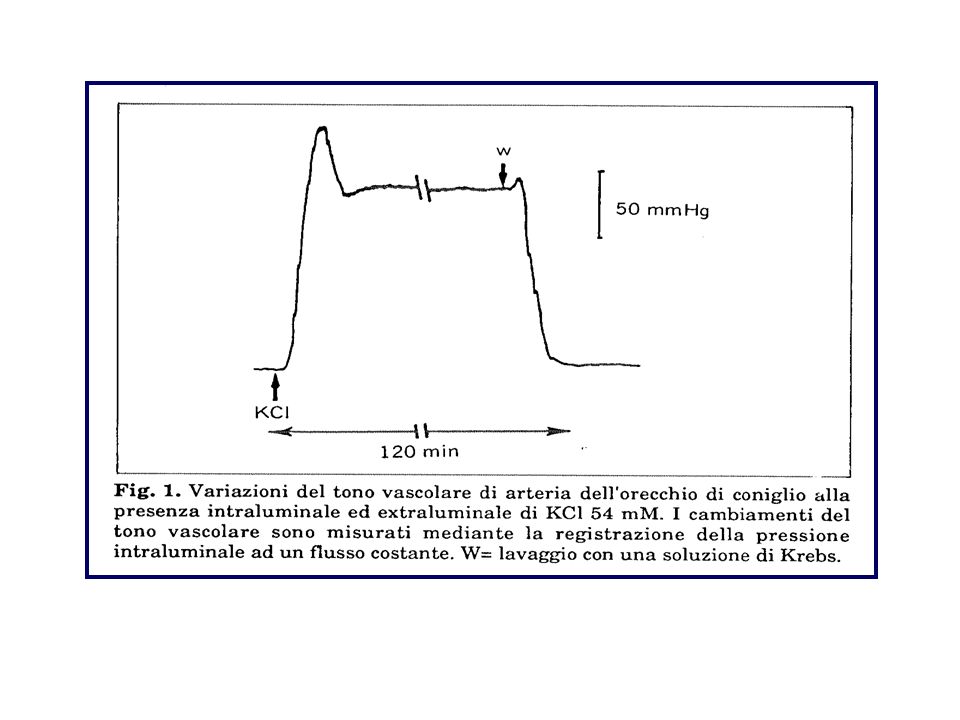
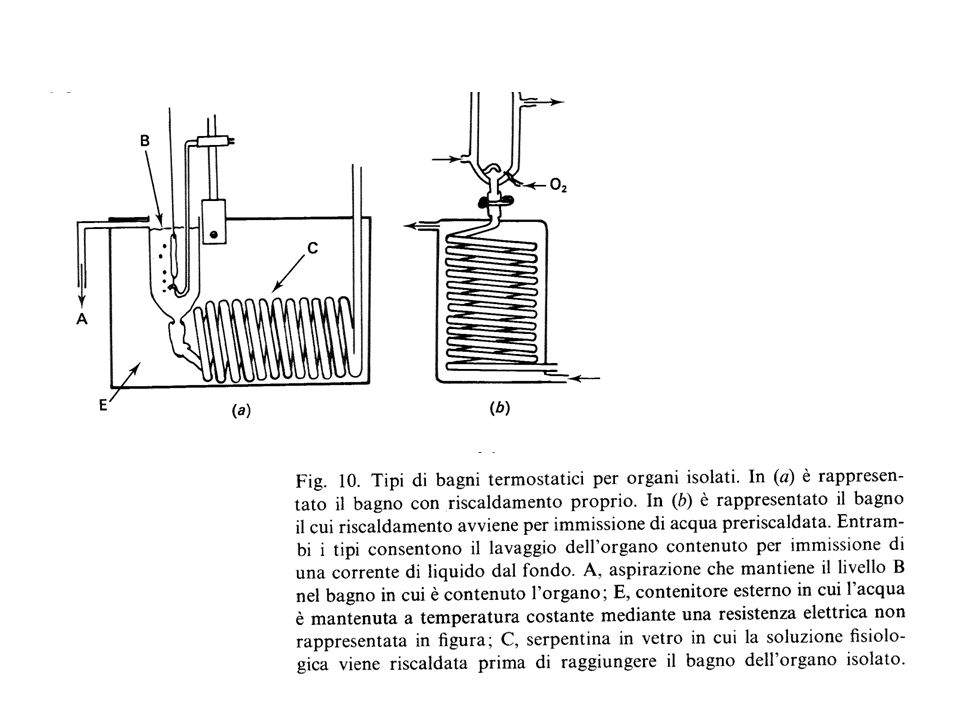
Soluzioni saline fisiologiche
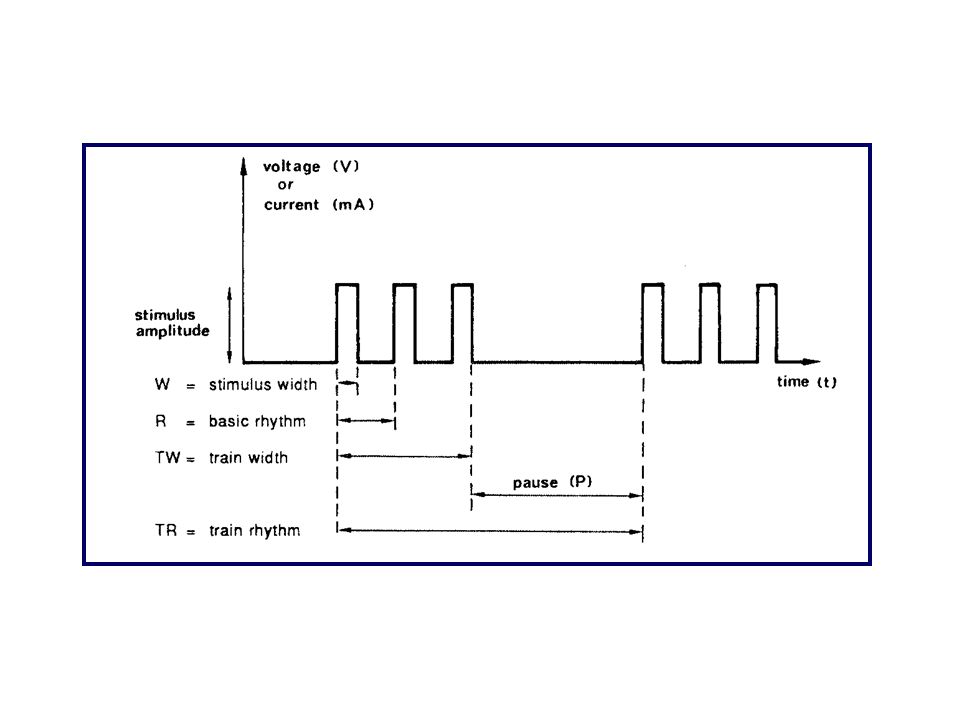
Temperatura Aereazione Apparati di registrazione Stimolazione

4
Composizione ionica Substrati energetici Tamponi Osmolarità

6
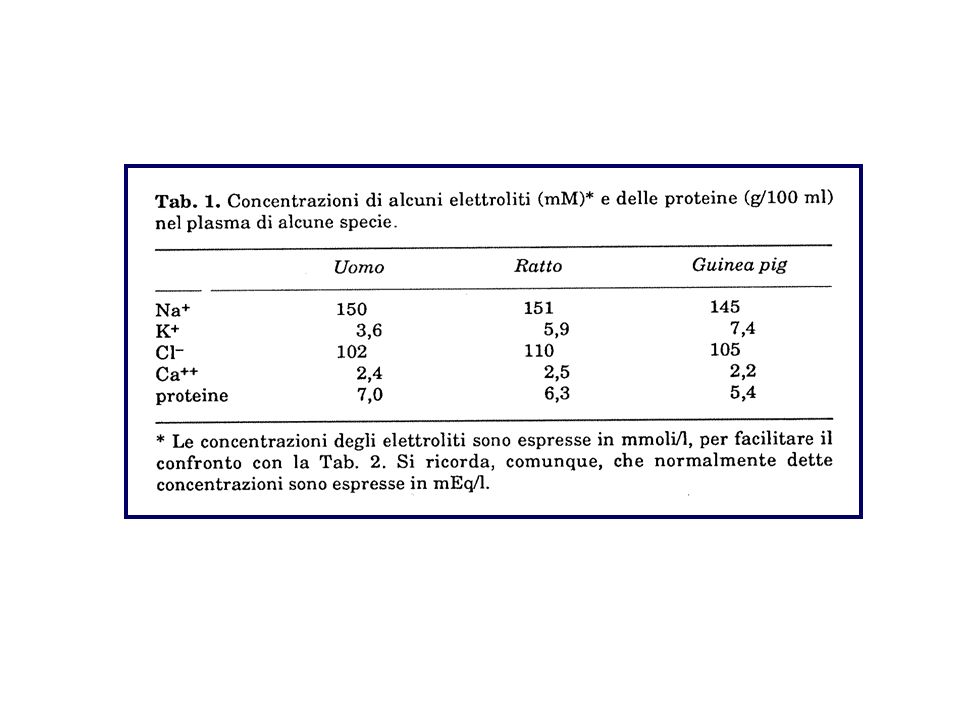
COMPOSIZIONE DI ALCUNE SOLUZIONI FISIOLOGICHE

8
Composizione ionica Substrati energetici Tamponi Osmolarità glucosio acetato, lattato, piruvato miscele di aminoacidi

9
Substrati energetici Tamponi Osmolarità Bicarbonato Fosfato

10
Substrati energetici Tamponi Osmolarità utilizzo di sangue completo; aggiunta di BSA aggiunta di sostanze varie (es. destrano) aggiunta di saccarosio

11
Soluzioni saline fisiologiche
Temperatura Aereazione Apparati di registrazione Stimolazione k(t+10) Q10 = kt
 Q10 = kt.")
13
Soluzioni saline fisiologiche
Temperatura Aereazione Apparati di registrazione Stimolazione

14
Soluzioni saline fisiologiche
Temperatura Aereazione Apparati di registrazione Stimolazione

15
CONTRAZIONE ISOTONICA

16
REGISTRAZIONE DI UNA CONTRAZIONE ISOTONICA

17
CONTRAZIONE ISOMETRICA

18
REGISTRAZIONE DI UNA CONTRAZIONE ISOMETRICA

21
Soluzioni saline fisiologiche
Temperatura Aereazione Apparati di registrazione Stimolazione elettrodi puntiformi elettrodi a campo

24
VALUTAZIONE QUANTITATIVA DEGLI EFFETTI FARMACOLOGICI

25
DOSI SINGOLE E DOSI CUMULATIVE

26
MUSCOLATURA SCHELETRICA
PREPARATI DI MUSCOLATURA SCHELETRICA

27
NERVO FRENICO-DIAFRAMMA DI RATTO (I)
")
28
NERVO FRENICO-DIAFRAMMA DI RATTO (II)
")
30
MUSCOLO RETTO ADDOMINALE DELLA RANA

31
Muscolatura liscia intestinale Ileo di cavia Digiuno di coniglio
Muscolatura liscia vascolare Striscia di aorta di coniglio Altri preparati di muscolatura liscia Striscia del fondo di stomaco di ratto Utero di ratto Catena tracheale di cavia Dotto deferente di cavia

32
ILEO DI CAVIA: PRELIEVO DEL PREPARATO

33
ILEO DI CAVIA: LAVAGGIO DEL PREPARATO

35
ILEO ISOLATO SECONDO TRENDELENBURG

36
Muscolatura liscia intestinale Ileo di cavia Digiuno di coniglio
Muscolatura liscia vascolare Striscia di aorta di coniglio Altri preparati di muscolatura liscia Striscia del fondo di stomaco di ratto Utero di ratto Catena tracheale di cavia Dotto deferente di cavia

37
DIGIUNO DI CONIGLIO: PRELIEVO DEL PREPARATO

38
DIGIUNO DI CONIGLIO SECONDO
LA TECNICA DI MAGNUS

39
DIGIUNO DI CONIGLIO: PRELIEVO SECONDO LA TECNICA DI FINKELMAN

40
DIGIUNO DI CONIGLIO SECONDO LA TECNICA DI FINKELMAN

41
Muscolatura liscia intestinale Ileo di cavia Digiuno di coniglio
Muscolatura liscia vascolare Striscia di aorta di coniglio Altri preparati di muscolatura liscia Striscia del fondo di stomaco di ratto Utero di ratto Catena tracheale di cavia Dotto deferente di cavia

42
STRISCIA DI AORTA DI CONIGLIO: PRELIEVO E ALLESTIMENTO DEL PREPARATO

43
STRISCIA DI AORTA DI CONIGLIO

44
PREPARAZIONI DI MUSCOLATURA CARDIACA

45
1. Preparati che utilizzano il cuore in toto
Cuore isolato e perfuso alla Langendorff 2. Preparati che utilizzano porzioni di muscolo cardiaco a) atri b) strips di ventricolo c) muscolo papillare
 atri. b) strips di ventricolo. c) muscolo papillare.")
46
ATRI DI CAVIA ISOLATI

47
ATRI DI CAVIA ISOLATI

48
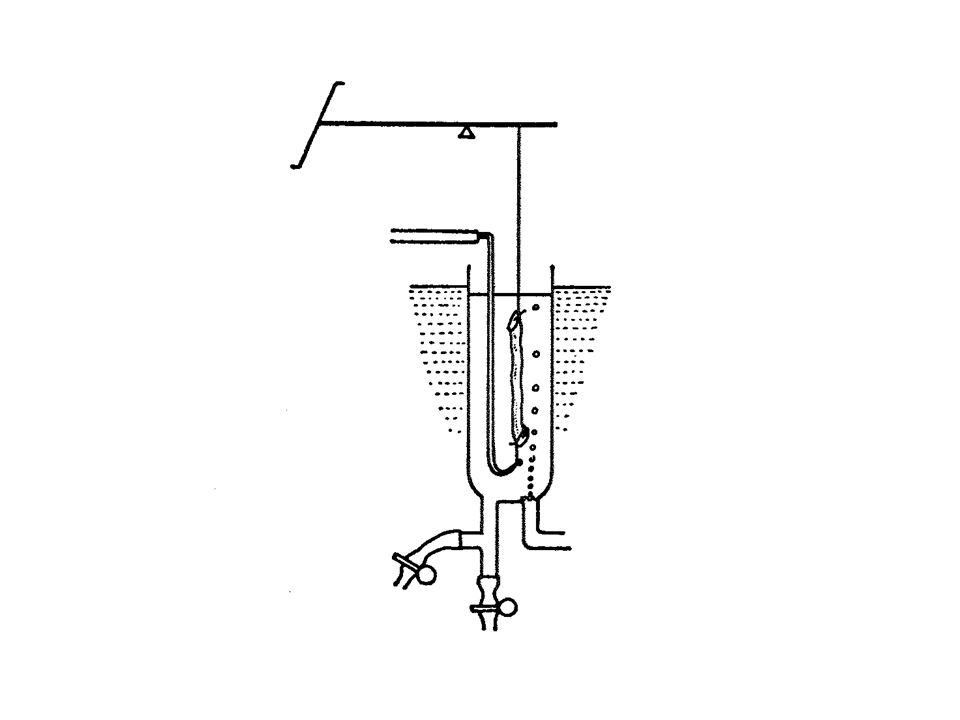
MUSCOLO PAPILLARE

49
Atri isolati di cavia Stimolazione elettrica ad una frequenza superiore a quella spontanea (240/min) tramite impulsi elettrici della durata di msec e di ampiezza pari al doppio della soglia. L’intensità dello stimolo si misura in Volt o in Ampere a seconda che si disponga di uno stimolatore a voltaggio costante o a corrente costante. I valori usati sono generalmente intorno a 5V oppure a 5 mA. Muscolo papillare di cavia o di ratto Striscia di ventricolo di cavia o di ratto La frequenza ottimale di stimolazione per ambedue i preparati è di 60/min. Si può usare una frequenza più elevata, per es. 150/min, quando si devono studiare gli effetti di farmaci che possono attivare processi di automatismo anomalo e che tendono quindi a provocare aritmie. Si usano stimoli della durata di msec e di ampiezza pari al doppio della soglia (circa 5V o 5mA).
.")
50
Test sull’attività di farmaci antiaritmici
Misura della massima frequenza seguita Misura del periodo refrattario funzionale

51
MUSCOLO PAPILLARE isometric
The force-velocity relationship describes how muscle fibers contract isotonically (i.e., shorten). To illustrate this, a papillary muscle is placed in an in vitro bath and a weight is tied to one end (Figure 1). When the muscle contracts, the fiber first generates tension isometrically (Figure 2, a® b) until the developed tension exceeds the load imposed upon the muscle. When this point is reached, the muscle fiber will then begin to shorten and the tension will remain constant and equal to the load (b® c). The maximal velocity of shortening (dL/dt) occurs shortly after the muscle begins to shorten. The muscle will continue to shorten until the muscle begins to relax. When active tension falls below the load (point c), the muscle will resume its resting length (i.e., preload) (point c). Active tension will continue to fall (c® d) isometrically until only the passive tension remains (point d).
. To illustrate this, a papillary muscle is placed in an in vitro bath and a weight is tied to one end (Figure 1). When the muscle contracts, the fiber first generates tension isometrically (Figure 2, a® b) until the developed tension exceeds the load imposed upon the muscle. When this point is reached, the muscle fiber will then begin to shorten and the tension will remain constant and equal to the load (b® c). The maximal velocity of shortening (dL/dt) occurs shortly after the muscle begins to shorten. The muscle will continue to shorten until the muscle begins to relax. When active tension falls below the load (point c), the muscle will resume its resting length (i.e., preload) (point c). Active tension will continue to fall (c® d) isometrically until only the passive tension remains (point d).")
52
If this same experiment with the papillary muscle is repeated with increasing loads (shift from curves a to c), there will be a decrease in both the velocity of fiber shortening and in the degree of shortening as shown in Figure 3.
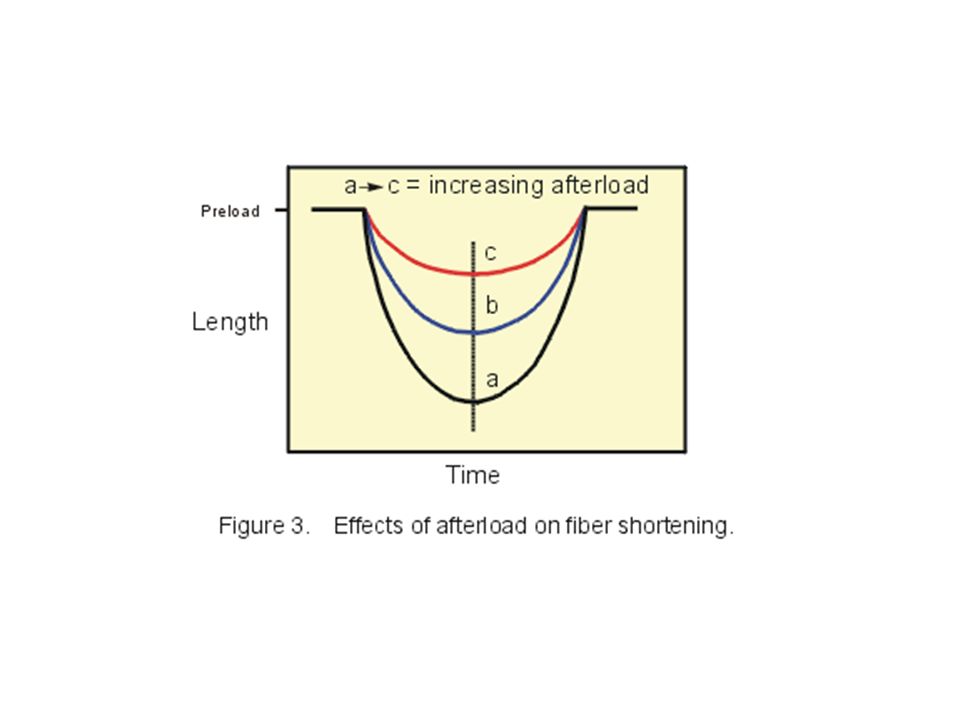
53
If the maximal velocity of shortening (dL/dt) is plotted against the load that the muscle fiber must shorten against (i.e., the afterload), an inverse relationship is found between velocity of shortening and afterload (Figure 4). In other words, the greater the afterload (the force in the force-velocity diagram), the slower the velocity of shortening. The x-intercept represents the point where the afterload is so great that the muscle fiber cannot shorten, and therefore represents the maximal isometric force. The y-intercept represents an extrapolated value for the maximal velocity (Vmax) that would be achieved if there were no afterload. The value is extrapolated because it cannot be measured experimentally. A muscle will not contract in the absence of any load. It is very important to note that a cardiac muscle fiber does not operate on a single force-velocity curve. This relationship is altered by changes in both preload and inotropy. The former shares some similarities with skeletal muscle; the latter, however, is unique to cardiac muscle. If the preload is increased, a cardiac muscle fiber will have a greater velocity of shortening at a given afterload. This occurs because the length-tension relationship dictates that as the preload is increased, there is an increase in active tension development. Once the fiber begins to shorten, the increased tension generating capability at the increased preload results in a greater velocity of shortening. In other words, increasing the preload enables to muscle to contract faster against a given afterload. Figure 5 shows the effects of increasing preload on the force-velocity relationship. Note that increasing preload increases the maximal isometric force as well as the shortening velocity at a given afterload. Furthermore, note that changes in preload do not alter Vmax. Changes in inotropy also alter the force-velocity relationship. If the inotropic state of the cardiac fiber is increased, there is a parallel shift up and to the right in the force-velocity curve such that there is an increase in both Vmax and in maximal isometric force (Figure 6). The increase in velocity at any given preload results from the increased inotropy enhancing force generation by the actin and myosin filaments, and increasing the rate of crossbridge turnover. The increase in Vmax is particularly noteworthy because Vmax represents the intrinsic capability of a muscle fiber to generate force independent of load, and this is one way in which inotropy can be defined. Therefore, Vmax can be viewed as an index or measure of inotropy for the muscle fiber.
. The increase in velocity at any given preload results from the increased inotropy enhancing force generation by the actin and myosin filaments, and increasing the rate of crossbridge turnover. The increase in Vmax is particularly noteworthy because Vmax represents the intrinsic capability of a muscle fiber to generate force independent of load, and this is one way in which inotropy can be defined. Therefore, Vmax can be viewed as an index or measure of inotropy for the muscle fiber.")
Presentazioni simili














, specializzato per la distribuzione di:>")
, specializzato per la distribuzione di: gas.>")




>")